

Abridged English version
Inoceramids are common in the geological site of Tercis (Landes, France) where the Campanian–Maastrichtian stage boundary is defined [4]. Fifty specimens were formerly identified by Annie Dhondt [9]; they documented the presence of the genus Trochoceramus below the boundary. More recently, contemporaneous faunas have been studied in several European and American areas [12,14] and the Tercis fauna has been revised in the light of this new knowledge using an additional material totalising 400 specimens.
Shells are irregularly distributed along the Tercis section with two rich beds around levels 46 and 67 (Fig. 1). Their size distribution shows that large specimens are restricted to the 33–98 interval. Following a first size decrease at level 98, a strong size drop of adult shells is documented between levels 111 et 117. The major drop in size around level 115 is a character of the species and has a stratigraphical meaning, whereas the smaller mean size observed around level 46 may be connected either to a problem of preservation or to the juvenile character of many specimens.
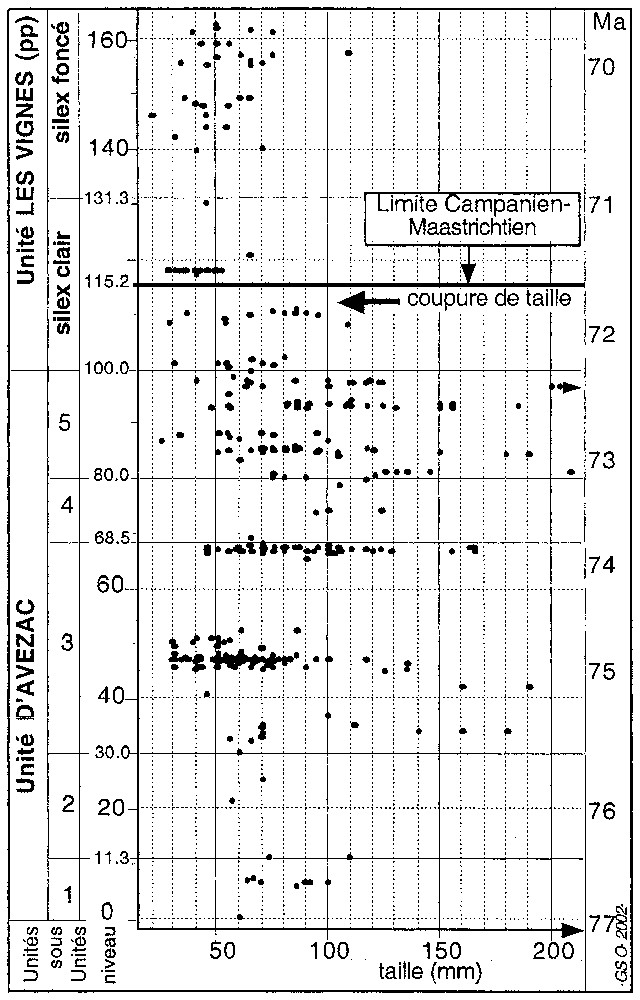
Distribution des tailles maximales de 400 inocérames du site de Tercis. Les sous-unités 1 à 5 sont alternativement riches et pauvres en glauconie; calibrage numérique [3,5] revu en fonction des corrélations par inocérames.
Size distribution of about 400 inoceramid specimens from Tercis. Subunits 1, 2, 3, 4, 5 are alternatively rich and poor in glaucony ; numerical calibration [3,5] revised according to inoceramid correlation.
Amongst 240 taxonomically identified specimens, 36 species-level taxa were recognised (Fig. 2), with two species described as new, dedicated to W.A. Cobban and N. Jolkicev. Two faunal turnovers are evidenced: the first one occurs between levels 94.0 and 96.7, the second one, between levels 111.3 and 117.2, is contemporaneous with the major size cut. As a result, inoceramids document the same two major evolutionary changes identified at Tercis for other fossil groups [6] and they allow the recognition of the stage boundary selected within the second change.
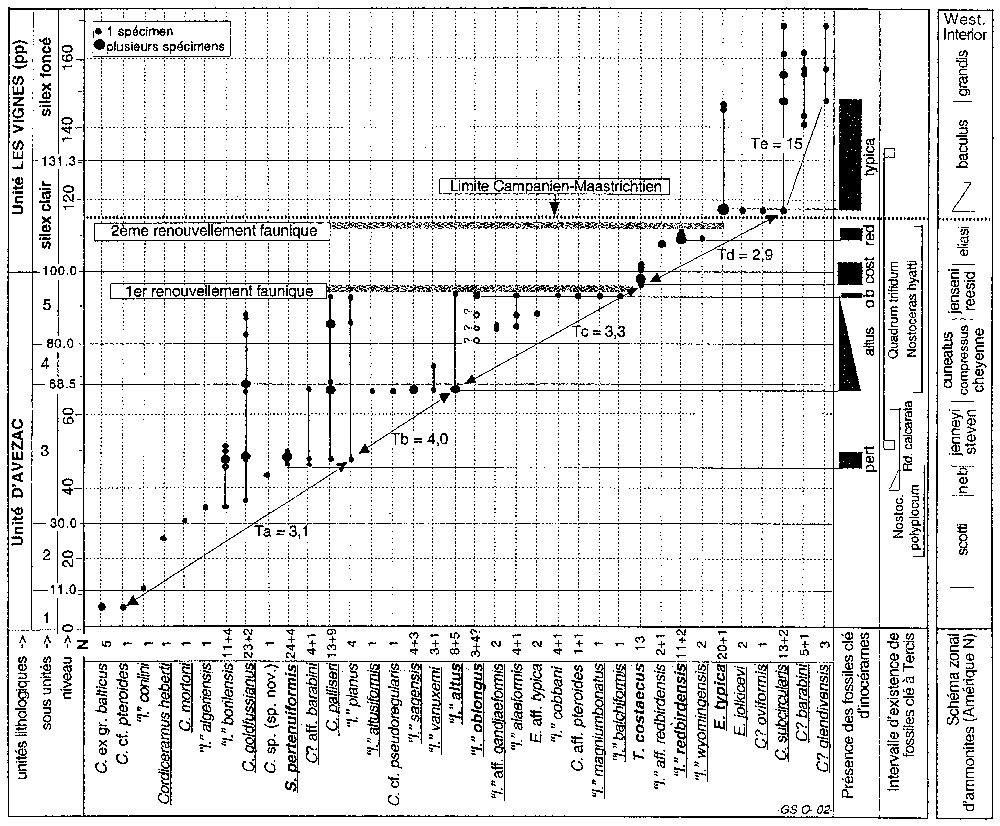
Distribution des taxons d'inocérames à Tercis. Noms soulignés : taxons communs Tercis–Amérique du Nord ; noms en gras: espèces clé ; N : nombre de spécimens probables (+incertains); Ta, Tb... : épaisseurs moyennes qui voient apparaı̂tre un nouveau taxon ; zones d'ammonites du Western Interior [1], de la base au sommet : Baculites scotti, Didymoceras nebrascense, D. stevensoni, Exiteloceras jenneyi, D. cheyennense, B. compressus, B. cuneatus, B. reesidei, B. jenseni, B. eliasi, B. baculus, B. grandis. Les « fossiles clés » de Tercis sont uniquement ceux dont l'intervalle d'existence est entièrement compris dans la section pour comparaison avec les intervalles d'existence très courts des inocérames. Abréviations : C., Cataceramus ; E., Endocostea ; ‘I.’, ‘Inoceramus’ ; S., Sphaeroceramus ; T., Trochoceramus.
Stratigraphical distribution of inoceramids along the Tercis succession. Underlined names : taxa common with North America ; bold : key taxa ; N : number of probable (+uncertain) specimens ; Ta, Tb... : mean thicknesses of appearance of a new taxon ; ammonites zones of Western Interior [1] ; from bottom to top : Baculites scotti, Didymoceras nebrascense, D. stevensoni, Exiteloceras jenneyi, D. cheyennense, B. compressus, B. cuneatus, B. reesidei, B. jenseni, B. eliasi, B. baculus, B. grandis. The column ‘fossiles clés à Tercis’ only shows the taxa for which the total range is present in the section for comparison with the generally very short total ranges of inoceramids. Abbreviations : C., Cataceramus ; E., Endocostea ; ‘I.’, ‘Inoceramus’ ; S., Sphaeroceramus ; T., Trochoceramus.
In an interval between levels 6 and 117 of the Tercis succession, one inoceramid species appears every 2–4 m in average, which corresponds to 0.1 to 0.2 Ma. It is a rate comparable to that observed in ammonites in the same outcrop [10]. Six inoceramid key fossils are recognised: Sphaeroceramus pertenuiformis, ‘Inoceramus’ altus, ‘I.’ oblongus, Trochoceramus costaecus, ‘I.’ redbirdensis, and Endocostea typica. They generally document short ranges (Fig. 2); much shorter than the ranges of the best key fossils of the Tercis series such as Radotruncana calcarata or Nostoceras polyplocum [4].
A distinct drop in an adult size located below the entry of E. typica at Tercis seems to be an over-regional phenomenon; it is also perceptible in the US Western Interior according to the published information [12]. The North American inoceramid fauna comprises 37 taxa [12, (fig. 5)] for the same time interval. Apart from T. costaecus, the other key fossils of Tercis are common to both areas as well as 14 accompanying taxa (Fig. 2). It must be pointed out that the recognised stratigraphical ranges of most of the key taxa are short and precise synchroneity of these known ranges between the two areas is difficult to document; there is a possibility that they correspond to diachronous partial records.
If one hypothesizes the contemporaneity of (i) the ‘I.’ redbirdensis occurrence around level 110 and the E. typica acme occurrence around level 117.5 at Tercis and (ii) the single occurrences of these two taxa in the Redbird section [12], if one admits a constant depositional rate in each of the two areas, then the Campanian–Maastrichtian boundary defined at Tercis may be drawn about 15 m below the B. baculus/B. eliasi ammonite zone boundary in North America. It might also be located slightly below in the upper part of the 80-m-thick B. eliasi zone (Fig. 2, rightmost column).
Among macrofossils, inoceramids unexpectedly constitute the most powerful stratigraphic tool to correlate the Campanian–Maastrichtian boundary between the two sides of the Atlantic Ocean; even microfossils are less powerful. The correlation potential is also excellent toward other European areas where the same forms have been quoted in the literature and these results encourage further study on this group, which quickly evolves for still 3–4 Ma before its extinction.
1 Introduction
L'investigation de terrain réalisée pour que le site géologique de Tercis (Landes, France) soit qualifié comme site de référence mondial pour la limite Campanien–Maastrichtien a mis en évidence la présence d'inocérames en nombre significatif [4] en accord avec les travaux antérieurs [2]. Cinquante fossiles de notre collection ont été identifiés de façon préliminaire par Annie Dhondt en 1993 [9]. Cette étude avait souligné l'intérêt du genre Trochoceramus, qui apparaissait, à Tercis, 0,7 Ma avant la limite nouvellement définie. Depuis lors, les études développées par l'un de nous (I.W.) sur les faunes contemporaines en Allemagne du Nord, dans la vallée moyenne de la Vistule, en Ukraine, dans le Nord-Est du Caucase [14] et en Amérique du Nord [12] ont mis en lumière l'intérêt stratigraphique de ce groupe fossile. On a revu la faune de Tercis sous cet éclairage après avoir complété notre collection, portée à près de 400 spécimens. Leur étude paléontologique, rendue cohérente avec celle des autres régions, permet de proposer une caractérisation biostratigraphique valable à l'est et à l'ouest de l'Atlantique pour des dépôts situés autour de la limite Campanien–Maastrichtien. Cette présentation rend compte des résultats obtenus sur notre collection, qui est la seule dont les éléments puissent être situés très précisément ainsi que de leurs conséquences sur les corrélations Europe–Amérique, pour lesquelles les inocérames se révèlent, de façon inattendue, comme l'outil le plus efficace parmi les macrofossiles.
2 Le site de Tercis et la limite Campanien–Maastrichtien
Le site de Tercis, limitrophe de Dax (Landes), était situé, voici 75 Ma, sur la marge septentrionale du domaine téthysien, dans un golfe de l'Atlantique allongé au nord des Pyrénées actuelles. Les dépôts de cette époque sont carbonatés [8] et suggèrent un environnement de plate-forme continentale externe.
Le passage du Campanien au Maastrichtien se fait sans interruption décelable dans des couches riches en témoignages diversifiés, qui ont permis d'identifier, notamment, une évolution biologique s'accélérant à deux reprises autour des cotes 98 et 115 [6]. La limite entre les deux étages a été définie par convention à la cote 115,2 selon le principe nouvellement élaboré d'un choix multicritère tenant compte de la position de 12 événements marqueurs [4] autour d'un événement guide, afin d'éviter la sacralisation du critère unique.
3 Les inocérames de Tercis
Les inocérames constituent la macrofaune la plus abondante des dépôts du site géologique de Tercis, mais leur récolte est difficile. La déformation tectonique a souvent altéré les fossiles, dictant une sélection sévère sur le terrain, où les spécimens de plus de 15 à 20 cm ne peuvent être extraits sans dégât. Pour les plus petits, seul le dégagement fin au laboratoire permet l'obtention de spécimens déterminables. Nos spécimens sont localisés dans la section avec une précision de l'ordre du décimètre, ce qui représente ±5 ka [7]. Leur répartition verticale est irrégulière, avec deux bancs riches autour des cotes 46 et 67 (Fig. 1). Ailleurs, les fossiles sont souvent rassemblés en niveaux comportant jusqu'à quelques individus au mètre carré.
La plus grande dimension des individus a été mesurée systématiquement pour les spécimens dégagés au laboratoire, ainsi que sur le terrain pour quelques grands exemplaires (Fig. 1). Des spécimens de plus de 130 mm ne sont présents qu'entre les cotes 33 et 98, avec des « géants » dans l'intervalle 80–98. Dans le banc riche inférieur, autour de la cote 46, les spécimens ont une taille moyenne inférieure à celle observée au-dessous comme au-dessus. La phase d'identification de notre étude a permis de relier ce fait à la fréquence des formes juvéniles. Les pycnodontes présentent un phénomène voisin (niveaux riches uniquement composés de formes juvéniles) ; ceci suggérerait une causalité environnementale, mais il pourrait s'agir aussi d'un effet lié à une fossilisation sélective.
Une première coupure peut être située vers la cote 98, où la taille moyenne s'abaisse. Plus remarquable est la coupure située entre les niveaux 111 et 117 de la section, qui sépare des populations de taille moyenne (70 à 80 mm) et des populations de vraiment petite taille (40 mm en moyenne). Dans les 20 m au-dessus, un très petit nombre d'individus de plus de 75 mm a été observé, ainsi que des traces de grandes coquilles à la cote 172. L'interprétation (environnementale ou évolutive) de cette brusque diminution de taille mérite d'être posée. La phase de détermination de notre étude a révélé qu'il s'agissait d'un caractère propre aux espèces, ce qui suggère une valeur évolutive d'intérêt stratigraphique pour cette observation morphologique simple.
4 Paléontologie et distribution
Parmi les 240 spécimens identifiés, 36 taxons sont notés sur la Fig. 2. Six autres formes encore non déterminées existent, dont cinq provenant des niveaux situés entre les cotes 80 et 98. Deux espèces nouvelles ont été nommées à Tercis et dédiées à W.A. Cobban et N. Jolkicev. L'abondance de certains taxons a permis de préciser la validité de quelques espèces ; c'est ainsi que Cataceramus goldfussianus (d'Orbigny), défini en Charente-Maritime, et C. gandjaensis (Aliev), défini dans le Caucase, ont pu être mis en synonymie au vu des formes intermédiaires de Tercis ; le même principe a été appliqué pour mettre en équivalence Trochoceramus costaecus (Khalafova) et T. morgani, tel que ce nom a été utilisé particulièrement par Walaszczyk [14]. Le détail des déterminations est publié par ailleurs [13].
Deux renouvellements complets des faunes apparaissent, le premier entre 94,0 et 96,7 et le second entre 111,3 et 117,2, pratiquement en accord avec les deux accélérations d'évolution de l'ensemble des fossiles du site [6]. La coupure lithologique principale (apparition de silex à la cote 100) concorde presque avec le premier changement ; la coupure de taille des espèces d'inocérames renforce le second événement. La limite d'étage, conventionnellement fixée en correspondance avec la seconde accélération de l'évolution, pourra donc être reconnue avec les inocérames.
La faune de Tercis n'est pas encore entièrement connue, mais celle identifiée ici est représentative de l'évolution du groupe dans cette expansion orientale de l'Atlantique autour de la limite Campanien–Maastrichtien. Pour quantifier l'évolution dans l'intervalle de récolte compris entre les cotes 6,5 et 148,2, un « taux d'apparition » a été calculé pour divers intervalles notés a, b, c, d, e, limités par les changements visibles sur la Fig. 2 aux cotes voisines de 47, 67, 97 et 117. Le taux, T, sera le quotient entre l'épaisseur de sédiment en mètres et le nombre d'espèces nouvelles. On obtient les valeurs de Ta : (47−6,5)/13=3,1 ; Tb : (67−47)/5=4,0 ; Tc : (97−67)/9=3,3 ; Td : (117−97)/7=2,9 ; Te : (148−117)/2=15. Si l'on tient compte des cinq formes non identifiées entre 67 et 97,l, Tc devient 30/14=2,1 : un taxon apparaı̂t tous les 2,1 m. Ainsi, il semble apparaı̂tre un taxon tous les 2 à 4 m, jusqu'à – et y compris – la base du Maastrichtien. D'après le taux moyen de dépôt admis pour ces divers intervalles [7], un taxon nouveau apparaı̂t tous les 0,1 à 0,2 Ma sur l'essentiel de la section. C'est un intervalle maximal. La partie maastrichtienne paraı̂t moins féconde à Tercis. On pourrait y voir le prélude à la disparition prochaine du groupe, mais nous avons peut-être une lacune d'information pour ces niveaux, car le groupe semble encore vigoureux plus haut dans le Maastrichtien inférieur, d'après nos observations (I.W.) dans d'autres bassins.
Six taxons peuvent être qualifiés de fossiles clés, parce qu'ils réunissent la plupart des qualités suivantes : présence en nombre suffisant, localisation stratigraphique limitée et connue, morphologie reconnaissable avec confiance, présence dans plusieurs régions. Sphaeroceramus pertenuiformis, défini en Amérique du Nord [12], est présent à Tercis (28 spécimens) dans l'intervalle 45,4–49,2 qui pourrait représenter 0,3 à 0,5 Ma de dépôt. ‘Inoceramus’ altus est représenté, dans notre collection, par une dizaine de spécimens dans l'intervalle 66,5–67,4 et par un autre à 93,6. L'acmé initiale représenterait moins de 0,5 Ma. Cette espèce nord-américaine a aussi été observée une fois en Pologne. ‘I.’ oblongus est encore une espèce nord-américaine. Elle est peu représentée à Tercis (trois spécimens sont sûrs) entre les cotes 93,0 et 94,0 ; son intervalle d'existence est de l'ordre de quelques dizaines de milliers d'années, mais l'espèce pourrait être présente un peu en dessous ; elle a été observée en Pologne centrale et en Bulgarie. Trochoceramus costaecus est bien représenté à Tercis (13 spécimens en collection et bien d'autres vus sur le terrain) exclusivement entre les cotes 96,7 et 101,9, c'est-à-dire durant un intervalle de dépôt estimé à 0,2–0,3 Ma. L'espèce fait partie d'un genre dont elle constitue l'apparition, aisé à reconnaı̂tre même en fragment sur le terrain [9,14], ce qui est très exceptionnel pour un inocérame. Cette espèce, définie dans le Caucase, a été citée en Pologne centrale et en Amérique du Nord. ‘I.’ redbirdensis est une espèce définie dans la province du Western Interior [12]. Nos 11 spécimens proviennent de l'intervalle 109,0–111,3, ce qui représenterait une durée de dépôt d'environ 0,1 Ma. Une forme affine semblant ancestrale à la base de cette distribution, suggère que nous aurions bien localisé une apparition évolutive. L'ornementation de côtes fortes est unique pour Tercis et permet encore une reconnaissance sur le terrain. Endocostea typica, toujours une espèce nord-américaine, est représentée par 20 spécimens dans notre collection, mais il en a été vu davantage encore sur le terrain entre les cotes 117,3 et 117,7, à raison de quelques spécimens par mètre carré localement, tandis que de rares individus sont présents autour de la cote 155, environ 1,5 Ma plus tard. Ainsi, quatre des six fossiles clé d'inocérames sont représentés à Tercis dans des intervalles courts, plus courts que ceux des autres fossiles clé récoltés dans cette succession (Fig. 2), même les plus momentanés, comme le foraminifère Radotruncana calcarata ou l'ammonite Nostoceras polyplocum [4,6].
La limite d'étage est située entre l'intervalle d'existence de ‘I.’ redbirdensis, accompagné de ‘I.’ wyomingensis et l'acmé de E. typica est accompagnée de trois autres taxons. Une recherche prolongée n'a pas permis de récolte entre ces niveaux, cette lacune d'information représentant 1/4 de Ma.
5 Comparaison avec l'Amérique du Nord
Il y a aussi, en Amérique du Nord, une coupure de taille des inocérames au niveau de l'apparition de E. typica, espèce observée là-bas dans des accumulations de coquilles de 30 à 50 mm. Environ 37 espèces sont citées [12, (Fig. 5)] pour l'intervalle de temps équivalent, d'après les études faites sur le terrain et en musée. Les taxons clé de Tercis (sauf T. costaecus) sont présents dans la province du Western Interior et 14 des espèces d'accompagnement sont les mêmes. Ces taxons clé ont une répartition souvent restreinte à un seul niveau, dont la succession verticale est semblable dans les deux bassins. On doit se poser la question du synchronisme de ces présences brèves entre les deux côtés de l'Atlantique, car cette brièveté pourrait traduire un enregistrement partiel des intervalles d'existence et il est alors peu probable qu'elles soient précisément synchrones.
À Tercis, on observe ‘I.’ redbirdensis entre 109 et 111 puis E. typica à 117,3–117,8, la limite d'étage étant fixée à 115,2. Dans la coupe de Redbird (Wyoming), ‘I.’ redbirdensis est signalé dans un niveau au milieu de la zone à Baculites eliasi, tandis que E. typica est présent dans un niveau à la base de la zone à Baculites baculus [12], ces deux niveaux étant distants de 50 m. Si l'on fait l'hypothèse que le niveau unique américain de ‘I.’ redbirdensis correspond à l'intervalle 110±1 à Tercis, que le niveau américain à E. typica correspond à l'intervalle basal de sa distribution à Tercis (117,5±0,3) et que les intervalles compris entre la limite (115,2) et les positions respectives de ces deux signaux sont proportionnels dans les deux bassins, alors on peut placer la limite Campanien–Maastrichtien 15 m sous la limite B. baculus/B. eliasi. E. elliptica d'Amérique pouvant correspondre à un moment plus jeune de la distribution à Tercis, la limite pourrait se situer en dessous, dans la moitié supérieure de la zone à B. eliasi, jusqu'à 40 m sous la limite interzone. Les inocérames permettent ainsi de préciser sensiblement notre estimation de corrélation antérieure [3,5].
6 Conclusions
Après 130 Ma de vie discrète du Lias à l'Aptien, puis un long temps (Albien à Campanien) pendant lequel une large répartition et une évolution rapide en ont fait de bons fossiles stratigraphiques, les inocérames demeurent, 3 à 4 Ma avant leur disparition vers le milieu du Maastrichtien, un groupe fertile en changements. À Tercis, les espèces se succèdent avec une rapidité équivalente à celle des ammonites [10]. Cependant, l'information est de nature différente avec, pour les ammonites, une présence dispersée illustrant des intervalles d'existence bien circonscrits, pour les inocérames, des arrivées brutales en quantité et qualité, séparées par des lacunes d'information. C'est ce qui peut être qualifié de bioévénements et qui semble caractéristique dans différents bassins. La causalité de ces bioévénements et, par suite, leur signification chronologique, caractère synchrone ou non en différents lieux d'observation, restent un mystère à éclaircir.
Six fossiles clés ont montré un remarquable potentiel de corrélation et le groupe se révèle plein de promesses pour une application à la fois vers le domaine boréal en Europe et vers l'autre côté de l'Atlantique. Il reste à entreprendre l'étude de ce groupe délaissé jusqu'ici dans les régions plus orientales de la Téthys, comme en Tunisie [11], car plusieurs formes clés sont reconnaissables sur le terrain et méritent d'être connues du géologue.
Remerciements
F. Amedro et F. Robaszynski ont bien voulu commenter ce travail et suggérer des améliorations. La correction de la version anglaise a été réalisée par M.-A. Lamaurelle. Les auteurs les remercient vivement pour leur coopération.