

Abridged English version
Extant stalked crinoids, especially pentacrinids, give a useful model to estimate palaeodepths of offshore environments [2–4,18]. Adaptive characters of pentacrinid stalk related to depth and current flow are the best tool that does not depend on taxonomy and can be applied to extinct taxa. Here, we analyse a crinoid-rich fauna collected in the Lutetian–Bartonian series exposed in the quarry of Miretrain at Angoumé (Aquitaine, southwestern France), and we compare palaeodepth estimates deduced from crinoids, benthic foraminifera and other palaeoenvironmental data. As the taxonomy of Eocene pentacrinids (Isselicrinus, Cainocrinus) is poorly documented except by a few studies [7,12], we remain here in open taxonomy. For bourgueticrinids (Conocrinus, Tormocrinus), we refer to a previous description of Eocene Pyrenean fauna [16,17].
The marl pit of Angoumé is located on the north flank of Tercis anticline and at the western edge of Dax diapir into the North-Pyrenean foreland basin (Fig. 1). The geological section (Fig. 2) exposes mainly the ‘Marnes de Miretrain’ Formation, which contains a fair number of layers (C0 to C6) with stalked crinoid ossicles (Table 1). The top of these marls passes rapidly into bioclastic limestones that were attributed to the ‘Calcaire de Siest’ Formation by Sztrákos et al. [21], and the outcrop ends at the Bartonian–Priabonian boundary. The marl series could be subdivided into the three following units. The lower part (lower pale-grey marl unit) is without large benthic foraminifera and dated NP15 from Mid-Upper Lutetian to Lower Bartonian. It contains (beds C0, C1 and C2) isolated stalk fragments and columnals belonging to a single new species of pentacrinid, Isselicrinus sp. 1 (Fig. 3a). Allochthonous large benthic foraminifera are frequent in the middle and upper units. About at the NP15–NP16 transition, the middle unit of pale-grey marls begins by a crinoid-rich mass flow (bed C3) mainly with ossicles and stalk fragments of Isselicrinus sp. 2 (Fig. 3d), the most frequent, and also I. sp. 3, Cainocrinus sp., and frequent large Conocrinus. Above, a bioclastic marly lens (bed C4) contains abundant ossicles and a few dorsal cups of Conocrinus elongatus and C. duperrieri, and less frequent columnals of Isselicrinus sp. 4 (Fig. 3b). Near the top of the middle unit (C5) a few isolated ossicles of Conocrinus and juvenile Isselicrinus were found. The upper unit displays dark-grey marls that belong to the Truncorotaloides rohri zone, and are characterized by frequent coarse detritic beds or lenses with reworked material from Tertiary–Mesozoic eroded basement [21]. It contains calcareous biodetritic layers (C6a) with large stalk fragments of Isselicrinus sp. 5 (Fig. 3c), columnals and dorsal cup of Conocrinus duperrieri and large Tormocrinus aff. archiaci. Isselicrinus sp. 2 displays affinities with I. diaboli, I. sp. 3 and I. sp. 5 with I. subbasaltiformis, and I. sp. 4 with I. didactylus. Cainocrinus sp. and Isselicrinus sp. 2 belong to an isocrinid morphotype adapted to turbulent current flow. The other species of the genus Isselicrinus show a balanocrinid morphotype adapted to laminar current flow.

Localisation et schéma géologique du bassin de l'Adour. Les croix indiquent les axes anticlinaux.
Location and geological sketch of the Adour Basin. Crosses indicate axes of anticlines.
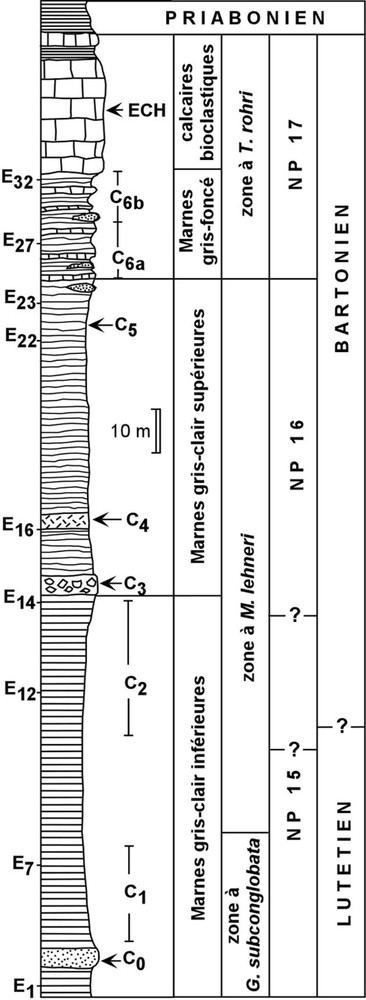
Série stratigraphique éocène affleurant dans la carrière de Miretrain. C : Niveaux à crinoïdes ; E : échantillons de Sztrákos et al. [21] ; ECH : gisement à échinides [15].
Stratigraphical series exposed in the Miretrain quarry. C: Beds with crinoids; E: sampling of Sztrákos et al. [21]; ECH: echinid-rich bed [15].
Répartition stratigraphique des crinoïdes pédonculés de la carrière de Miretrain. Localisation des niveaux, cf. Fig. 2. (b) : Phénotype balanocrine ; (i) : phénotype isocrine ; (g) : gros ; (m) : moyens ; (p) : petits ; xX : fréquence relative faible (x) ou forte (X)
Stratigraphical distribution of stalked crinoids from Miretrain quarry. See Fig. 2 for location of beds with crinoids. (b): Balanocrinid phenotype; (i): isocrinid phenotype; (g): large; (m): medium; (p): small; xX: low (x) or high (X) relative frequency
| Niveaux C | 0 | 1–2 | 3 | 4 | 5 | 6a | 6b |
| Isselicrinus (b)(p) | X | X | x | ||||
| Isselicrinus (b)(m) | X | X | |||||
| Isselicrinus (b)(g) | x | x | X | ||||
| Isselicrinus (i)(g) | X | ||||||
| Cainocrinus (i) | x | ||||||
| Conocrinus (p) | x | X | x | X | x | ||
| Conocrinus (g) | X | x | |||||
| Tormocrinus (p) | x | X | x | x | |||
| Tormocrinus (g) | X | ||||||
| Allochtonie | + | 0 | ++ | + | 0 | + ? | + ? |

Les quatre phénotypes du genre Isselicrinus des marnes éocènes de Miretrain. Articulations internodales du pédoncule. (a)–(c) Phénotypes balanocrines, (d) phénotype isocrine, (a) I. sp. 1 (niveau C2), (b) I. sp. 4 (niveau C4), (c) I. sp. 5 (niveau C6a), (d) I. sp. 2 (niveau C3).
The four phenotypes of the genus Isselicrinus from Eocene marls of Miretrain. Articular facets of internodal stalk joints. (a)–(c) Balanocrinid phenotypes, (d) isocrinid phenotype, (a) I. sp. 1 (from C2), (b) I. sp. 4 (from C4), (c) I. sp. 5 (from C6a), (d) I. sp. 2 (from C3).
Using adaptive characters related to depth known in extant pentacrinids [2,4], like the average columnal diameter, which decreases with depth, and the ratio of columnal height to diameter, which increases with depth (Fig. 4), we conclude that Isselicrinus sp. 1 lived in the deepest environment (600 m or more for the allochthonous sandy bed C0, about 800 m for C2, and more than 900 m for C1), and Isselicrinus sp. 5 in the shallower (200–300 m). The few Isselicrinus sp. 4 and the abundant bourgueticrinid fauna of C4 lived at intermediate depth (300–400 m), but slightly shallower than the biota of the ‘Marnes à pentacrines’ (I. didactylus) in the Eocene series of Biarritz, which were at a depth of 500–600 m [5,17]. The mass flow C3 gathered crinoids from different biota and depths: (1) Isselicrinus sp. 2 and large Conocrinus filtering turbulent water flow induced by very irregular substrata, probably with abundant food supply as observed in seamount environment [14], at a depth of either more than 400 m if the two taxa were living in the same community, or each at different depth such as less than 400 m for large Conocrinus and more than 400 m for Isselicrinus sp. 2; (2) Isselicrinus sp. 3, Cainocrinus sp. and small Conocrinus living at a depth of about 300 m in currents weakly turbulent near the bottom and laminar above; (3) Isselicrinus sp. 1 reworked from C2 and filtering laminar water flow above a flat muddy substrate. Data from Angoumé and other areas of western Europe [17] suggest an Eocene distribution of the genus Isselicrinus with three species having a balanocrinid phenotype and living at different depths, i.e. I. subbasaltiformis (200–300 m), I. didactylus (300–600 m), and I. sp. 1 (600–1000 m or more), and one species having an isocrinid phenotype (i.e. I. diaboli sensu lato) when turbulent water flow predominates.
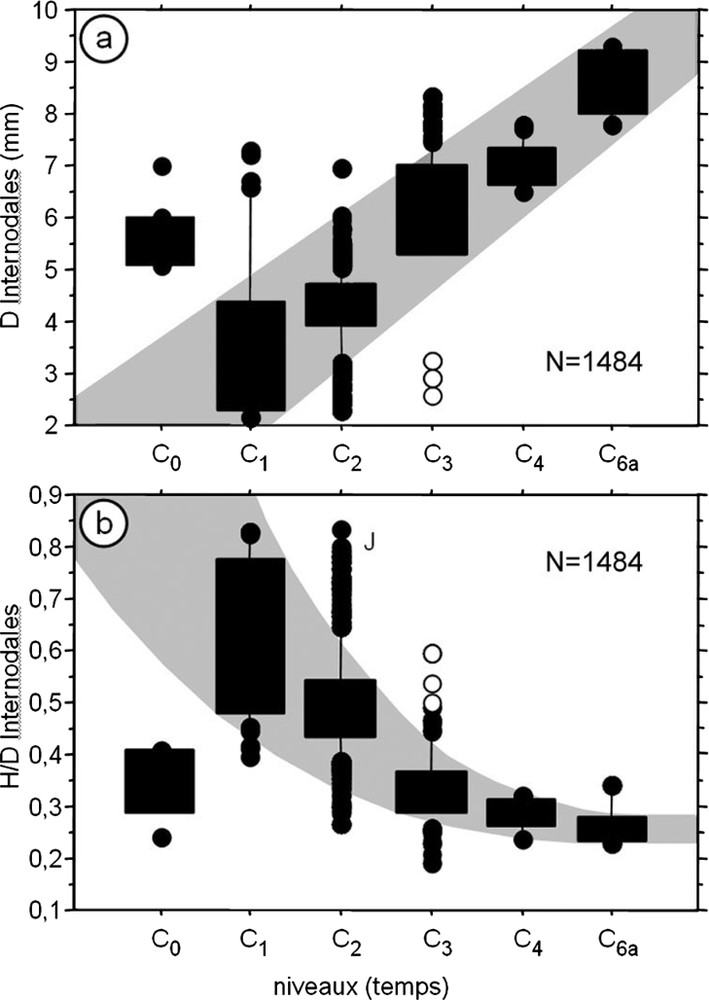
Analyse biométrique des columnales du genre Isselicrinus, recueillies dans la carrière de Miretrain. Paramètres dépendant de la bathymétrie. D : Diamètre, H : hauteur, J : spécimens juvéniles. Les larges bandes noires verticales indiquent le maximum de fréquence. Cercles blancs : columnales de I. sp. 1 remaniées dans C3. N = nombre de columnales.
Biometrical analysis on columnals of the genus Isselicrinus from Miretrain quarry. Parameters depending on depth. D: Diameter, H: height, J: juvenile specimens. Vertical wide black bands indicate highest frequencies. White circles: columnals of I. sp. 1 reworked into C3. N = number of columnals.
The benthic foraminifera associated with crinoids are used with the principles of depth estimation discussed in [10,20]. Nuttalides truempyi, classically used as an index of depths > 500 m [11] and frequently of 1000 m or more [9], is present in C1, and very scarce (reworked?) in C2. Cribroparella pteromphalia, Repmanina charoides and Bulimina trinitatensis are found in C2 suggesting a depth of 500–600 m. The mass flow C3 contains both shallow water taxa and bathyal species, like Cribroparella pteromphalia. Although large benthic foraminifera are frequent, taxa that could live in bathyal biota remain present up to the top of the middle unit (Bulimina trinitatensis in C5, and above Tritaxilina pupa, Uvigerina rippensis and U. acutocostata). In C6a, articles of large bathyal gorgonian (Isididae) were found. Numerous fragments of calcareous red algae appear at the base of the upper unit and were frequent above indicating the proximity of the euphotic zone. In the bioclastic limestone unit, echinid-rich fauna lived at a depth of about 50 m, and in marly layers relatively abundant plankton associated with ostracods and crinoids of bathyal affinities [15] suggests a location near offshore environment. A previous study of selachian fish fauna [1] shows that: (1) in C0, 50%of the taxa could live up to a depth of 1000 m, several genera (Apristurus, Deania, Centrosoymnus) having their optimum between 500 and 1000 m, or between 400 and 700 m (Etmopterus), (2) above C3 most taxa lived at a maximum depth decreasing from 500 m near C4 to 200 m near the top of the middle unit, a few taxa with their optimum between 400 m and 1000 m remaining present. Thus, taking in account that, in slope sedimentation, there are frequent and important transports of fossils before they are buried in composite assemblages [8], the palaeodepth estimation from stalked crinoid biota is consistent with arguments given by other fossils.
The series exposed in the Miretrain quarry and its succession of crinoid assemblages from deep to shallow water environments document a phase of progradational sedimentary system (Fig. 5), with a rapid decrease of depth, which results from a siliciclastic supply higher than the space creation by subsidence or sea-level change [19]. This phase took place between two major tectonic events: (1) during the Late Ypresian–Early Lutetian and (2) during the Late Bartonian–Priabonian [6,13]. The vicinity of the submerged diapiric structures of Tercis and Dax suggests local halokinetic deformation, which could explain the sudden mass flow C3, without any relation to regional unconformity. However, at the base of dark-grey marls, the rapid increase of reworked material from shallow water and from Mesozoic basement documents the uplift and erosion of the Tercis anticline and the beginning of the second tectonic event.

Déformation synsédimentaire du diapir de Tercis et progradation du talus sédimentaire alimentant en faunes allochtones la série marneuse à Miretrain. F : Grands foraminifères benthiques ; (a) et (b) : communautés à crinoïdes allochtones, remaniées dans le glissement en masse C3. Explications dans le texte.
Synsedimentary uplift of the Tercis diapir and progradation of the sedimentary slope providing allochthonous fauna in the marl series at Miretrain. F: Large benthic foraminifera; (a) and (b) allochthonous crinoid communities reworked in the C3 mass flow. See text for explanation.
1 Introduction
L'Éocène du domaine pyrénéen possède de nombreux gisements à crinoïdes pédonculés [17]. Le genre Conocrinus est le mieux représenté, en abondance et en diversité [16]. En dehors du célèbre gisement des « marnes à pentacrines » de la Côte des Basques (Biarritz), les pentacrines sont rarement signalées, bien que d'un grand intérêt pour les reconstitutions paléobathymétriques [3,18]. La marnière de Miretrain à Angoumé, près de Dax, localité-type de la formation des « marnes de Miretrain » [21], nous a livré une succession de faunes à crinoïdes, riches en pentacrines et accompagnées de foraminifères planctoniques et benthiques, permettant une bonne localisation stratigraphique et une comparaison des estimations bathymétriques issues des crinoïdes, des foraminifères et d'autres indices biologiques et taphonomiques.
2 Contexte géologique et stratigraphique
La localité d'Angoumé se situe dans l'avant-pays pyrénéen, sur le flanc nord de l'anticlinal de Tercis (Fig. 1). Ce dernier est un des éléments de la tectonique salifère qui affecte la couverture sédimentaire de l'Aquitaine méridionale. Discordances et remaniements de matériel mésozoïque indiquent que les marnes de Miretrain se sédimentent à la suite d'une importante phase de déformation datant de l'Yprésien terminal–Lutétien inférieur, et d'une érosion jusqu'au cœur triasique de certains diapirs, comme celui de Tercis [6,13]. Les biozonations utilisées ici sont tirées de Sztrákos et al. [21].
La marnière d'Angoumé comprend sept couches riches en crinoïdes (Fig. 2). D'une épaisseur d'environ 160 m, la série débute par des marnes gris clair, correspondant au Lutétien (zone à Globigerinatheka subconglobata et zone NP15) et se développant principalement au Bartonien (dans la zone à Morozovella lehneri et la zone NP16), puis se charge en bancs calcaréo-gréseux vers le sommet, où les marnes prennent une teinte plus foncée (zone à Truncorotaloides rohri et zone NP17). Ces marnes sont surmontées par des calcaires biodétritiques de même âge, qui sont assimilés au « calcaire de Siest » in [21], et au sommet desquels on passe au Priabonien.
L'ensemble des marnes gris clair comporte des lentilles ou passées détritiques, témoignant d'une sédimentation gravitaire qui s'accentue brusquement à son sommet. L'une d'elles, située au passage NP15–NP16, correspond à un glissement en masse, avec des éléments centimétriques issus de la dislocation des marnes sous-jacentes, emballés dans une matrice sableuse riche en crinoïdes, fragments de silex et graviers de calcaires crétacés. Ce niveau (Fig. 2, niveau 3 à crinoïdes) correspond à une limite au-dessus de laquelle les marnes se chargent fréquemment en grands foraminifères benthiques. Le passage entre les marnes gris clair et les marnes gris foncé sommitales, alternant avec des bancs calcaréo-gréseux, est marqué par une accentuation rapide du détritisme et par des ravinements locaux. Sztrákos et al. [21] envisagent une lacune de sédimentation, qui pourrait accentuer la coupure biostratigraphique (absence de la zone à Orbulinoides beckmanni). Néanmoins, il est difficile de suivre sur le terrain une discontinuité sédimentaire nette. Au-dessus, des lentilles détritiques ravinantes très hétérométriques à galets verdis, nodules ferrugineux et éléments centimétriques à décimétriques de matériel d'âge Triasique à Lutétien, témoignent de l'importance de l'allochtonie et de l'érosion qui affectait les reliefs environnants.
La marnière d'Angoumé peut donc être subdivisée (Fig. 2), de bas en haut, en :
- • (1) des marnes gris clair inférieures massives et dépourvues de matériel allochtone macroscopique, à l'exception d'une lentille de marnes sableuses ferrugineuses à la base ;
- • (2) des marnes gris clair supérieures, à passées bioclastiques allochtones, mais sans banc calcaréo-gréseux ;
- • (3) des alternances de marnes gris foncé et de bancs calcaréo-gréseux à matériel allochtone souvent grossier et abondant ;
- • (4) une barre de calcaire bioclastique lité à fins interlits marneux.
Au sein d'une période de forte alimentation terrigène [19], il s'agit de quatre phases du comblement sédimentaire d'une dépression et/ou de la progradation d'un talus s'appuyant sur les reliefs sous-marins environnants. D'après les micro-organismes planctoniques [9], on passerait ainsi d'une profondeur d'environ 1000 m en (1) à 700 m en (2), puis à 100 m en (3), et enfin à 50 m en (4).
3 La faune de crinoïdes pédonculés
La plupart des articles du pédoncule (columnales) de pentacrine récoltés dans la carrière de Miretrain appartiennent au genre Isselicrinus (Crétacé supérieur–Miocène), ayant un pédoncule dont les nodales ne portent généralement que deux ou trois cirres, et avec un nombre d'internodales généralement supérieur à 20. Quelques fragments de pédoncules se rapportent au genre Cainocrinus, uniquement éocène, aux nodales portant toujours cinq cirres et au nombre d'internodales inférieur à 15. Malgré quelques descriptions précises [7,12], nous ne disposons d'aucune synthèse détaillée sur les pentacrines éocènes, dont la taxonomie reste confuse au niveau de l'espèce. Nous laisserons donc les différentes espèces distinguées en nomenclature ouverte et indiquerons les espèces affines les plus proches. Pour les Bourgueticrinidae (Conocrinus, Tormocrinus), se caractérisant par des articulations du pédoncule à symétrie bilatérale, nous nous référerons aux études menées sur la faune éocène du domaine pyrénéen [16,17].
Les marnes gris clair inférieures ne livrent qu'une seule espèce, probablement nouvelle, Isselicrinus sp. 1, de taille petite à moyenne, au pédoncule ornementé ou lisse (Fig. 3a). Les fragments de pédoncule ou les columnales isolées sont les plus fréquents dans le niveau C2. Quelques exemplaires ont été trouvés remaniés à la base du niveau C3, tandis que ceux de C0 ont été pris dans un glissement en masse. Leur dispersion dans les marnes C1 et C2 et leur très bon état de conservation plaident en faveur d'un enfouissement dans leur biotope d'origine.
La base des marnes gris clair supérieures (niveau C3, à forte allochtonie) a livré trois espèces de pentacrines. La plus fréquente est une grande forme isocrine, Isselicrinus sp. 2 (Fig. 3d), très proche d'Isselicrinus diaboli. Elle est accompagnée de quelques columnales d'Isselicrinus sp. 3, grande forme balanocrine évoquant Isselicrinus subbasaltiformis, et d'une forme isocrine attribuée au genre Cainocrinus. Les columnales de Conocrinus sont abondantes. Elles appartiennent au moins à deux espèces différentes, l'une de grande taille, fixée par des rhizoïdes pénétrant le sédiment, l'autre de plus petite taille. Une coupe dorsale de Tormocrinus a été récoltée.
Le niveau C4 est une coulée bioclastique riche en grands foraminifères benthiques et en columnales de Conocrinus. Plusieurs coupes dorsales récoltées permettent d'identifier Conocrinus elongatus et C. duperrieri. Les columnales d'une grande pentacrine, Isselicrinus sp. 4, s'avèrent proches de celles d'I. didactylus (Fig. 3b).
Au sommet des marnes gris clair supérieures, le niveau C5 a livré quelques columnales dispersées, attribuables à Conocrinus et à un petit Tormocrinus, et un fragment de pédoncule juvénile d'Isselicrinus, l'ensemble étant probablement subautochtone.
Dans les marnes gris foncé, un premier niveau à crinoïdes (C6a) correspond à des marnes calcareuses biodétritiques où les columnales et coupes dorsales de petits Conocrinus sont fréquentes. Deux coupes dorsales sont attribuables à des spécimens juvéniles de C. duperrieri. Des columnales et coupes dorsales proches de celles de Tormocrinus archiaci se différencient de cette dernière espèce par une taille plus grande et une hypermorphie des apophyses interbrachiales caractéristiques de ce taxon. Plusieurs segments de pédoncules d'une grande pentacrine, Isselicrinus sp. 5 (Fig. 3c), sont très proches de ceux d'I. subbasaltiformis, espèce principalement connue de l'Éocène inférieur [12]. Le second niveau à crinoïdes des marnes gris foncé (C6b) a livré des columnales de petits Conocrinus et quelques coupes dorsales de Tormocrinus archiaci, de taille ordinaire. Le bon état de conservation, la variété des pièces squelettiques (columnales, brachiales, coupes dorsales) et leur dispersion dans les lits marneux constituant l'ensemble C6 suggèrent un enfouissement rapide à proximité du biotope.
Il existe donc un changement net dans les faunes de crinoïdes à partir du niveau C3, avec l'apparition d'assemblages à Conocrinus dominants et à grands Isselicrinus (Tableau 1). En dessous, il s'agit d'un assemblage monospécifique stable, à petits Isselicrinus. La diversification maximale du niveau C3 ne correspond pas à un optimum écologique pour les crinoïdes, mais, au contraire, à une déstabilisation sédimentaire qui accumule, dans le milieu de dépôt, des organismes provenant de divers biotopes.
Les niveaux à crinoïdes (C) ont été replacés au sein de l'échantillonnage (E) effectué par Sztrákos et al. [21]. La lentille de marne sableuse C0 et les marnes C1 correspondent à l'intervalle E1–E7 (Lutétien, zone à Globigerinatheka subconglobata). C2 couvre l'intervalle entre E12 et E14 et appartient donc à la zone à Morozovella lehneri. Dans un prélèvement de marne situé 2 m sous la base de C3, les foraminifères planctoniques sont très fréquents. Avec la présence de Turborotalia frontosa, T. possagnoensis, Hantkenina dumblei, H. liebusi, H. longispina, ils situent le sommet de C2 dans le Bartonien au sein de la partie médiane de la zone à Morozovella lehneri, juste après la disparition d'Acarinina broedermanni. C3, proche de E14, et C4, proche de E16, appartiennent à la même biozone, dans l'intervalle postérieur à la disparition d'A. broedermanni et antérieur à l'apparition de Turborotalia pomeroli. Les foraminifères planctoniques les plus fréquents dans C3 sont Subbotina cryptomphala, S. eocaena, Globigerinatheka mexicana, G. subconglobata, Guembelitrioides higginsi et Hantkenina dumblei ; on note la présence de Turborotalia frontosa et T. possagnoensis. C4 n'a fourni que de rares foraminifères planctoniques du Bartonien. C5, entre E22 et E23, se situe toujours dans la zone à M. lehneri avec Turborotalia frontosa, T. possagnoensis, Morozovella spinulosa, Truncorotaloides rohri, T. topilensis, et Guembelitrioides higginsi. C6a qui inclut E27, a livré Turborotalia cerroazulensis, espèce qui apparaît à la base de la zone à Truncorotaloides rohri.
4 Interprétations paléobathymétriques
L'ampleur des transferts gravitaires sur les pentes bathyales rend souvent l'allochtonie des bioclastes plus fréquente que leur autochtonie, ce qui amène, dans les études de faciès sédimentaires, à sous-estimer la bathymétrie des milieux de dépôt (discussion in [8]). En conséquence, le biotope (milieu de vie) des organismes et le lieu de leur enfouissement définitif (milieu de dépôt) doivent être impérativement distingués et, en cas de mélange, le biotope le plus profond prime sur les autres pour l'estimation du milieu de dépôt.
Ce sont les convergences écophénotypiques qui sont utilisées pour la comparaison entre les formes actuelles et fossiles de crinoïdes [3]. Elles sont indépendantes des taxons et s'appliquent aux genres fossiles, même s'ils n'ont plus de représentants actuels. En revanche, pour les foraminifères benthiques et les sélaciens, ce sont principalement les affinités taxonomiques avec les faunes actuelles qui fondent les interprétations, en considérant que les taxons repères vivaient aux mêmes profondeurs dans le passé.
4.1 Arguments tirés des crinoïdes
Les observations et expérimentations in situ dans l'océan actuel, qui se sont multipliées au cours des deux dernières décennies, ont confirmé et précisé les contraintes qui régissent la répartition bathymétrique des crinoïdes pédonculés, et notamment des pentacrines [2,4]. Étant des animaux sciaphiles, rhéophiles et dont la morphologie fonctionnelle ne tolère pas les fortes turbulences induites par les houles de tempêtes, ils sont cantonnés à des profondeurs supérieures à 100 m.
Les pentacrines, notamment les caractères de leur pédoncule, sont particulièrement sensibles au type d'écoulement du courant : ainsi, les formes isocrines sont les mieux adaptées aux écoulements turbulents, les formes balanocrines aux écoulements laminaires. Parmi les pentacrines de l'Éocène d'Angoumé, Isselicrinus sp. 2 et Cainocrinus sont les seules formes isocrines ; tous les autres Isselicrinus sont des formes balanocrines typiques.
Le diamètre moyen du pédoncule des pentacrines actuelles diminue avec la profondeur, tandis que le rapport H/D entre la hauteur des columnales et leur diamètre augmente [4]. Ces critères biométriques simples d'évaluation paléobathymétrique sont d'autant plus pertinents qu'ils s'appliquent à une entité taxonomique homogène, comme ici le genre Isselicrinus. L'augmentation du diamètre moyen des columnales d'Isselicrinus récoltées à Angoumé, couplée à la diminution du rapport H/D (Fig. 4), montre que la profondeur du biotope à pentacrines a été maximale en C1 et minimale en C6. Pour les niveaux à Isselicrinus sp. 1 (C0, C1, C2), le mode de fréquence des tailles change au sein du créneau bathymétrique occupé par l'espèce : C1 se situe à sa limite inférieure, C2 au milieu et C0 (allochtone) provient d'une moindre profondeur sans atteindre la limite supérieure du créneau. Les écarts significatifs constatés entre les Isselicrinus des différents niveaux incitent à envisager des biotopes successifs, séparés par plusieurs centaines de mètres de tranche d'eau, ce qui permet d'estimer au minimum à 700 m la différence de profondeur entre le biotope d'Isselicrinus sp. 1 du niveau C1 et celui d'Isselicrinus sp. 5 du niveau C6a. La profondeur minimale connue pour les formes balanocrines actuelles est d'environ 250 m (Neocrinus decorus) ; c'est aussi la profondeur où se trouvent les plus grosses pentacrines actuelles. Le biotope à Isselicrinus sp. 5 se situait vraisemblablement à une profondeur comprise entre 200 et 300 m. Pour le biotope du niveau C1, on aboutirait ainsi à une profondeur supérieure ou égale à 900–1000 m, qui entre dans le créneau inférieur de la distribution bathymétrique des pentacrines actuelles, qui s'étend principalement de 200 à 1200 m. Les espèces ou phénotypes de pentacrine qui peuplent ces profondeurs dans le Pacifique occidental n'apparaissent pas avant 600 m, ce qui pourrait être le cas d'Isselicrinus sp. 1. Le biotope des spécimens de C0 se situerait au moins vers 600 m et celui de C2 vers 800 m, ceci en gardant l'estimation minimale de 900–1000 m pour celui de C1.
Dans le niveau C3, trois associations sont mélangées, correspondant à trois biotopes différents, étagés du moins profond au plus profond de la manière suivante :
- • (a) les Isselicrinus sp. 2 et les grands Conocrinus, d'un milieu parcouru par des courants soutenus à écoulement turbulent dominant lié à l'irrégularité du substrat, probablement des fonds mixtes indurés, voire rocheux (affleurements ou blocs de Crétacé), et meubles ;
- • (b) les Isselicrinus sp. 3, Cainocrinus sp. et les petits Conocrinus, d'un milieu parcouru par des courants modérés à écoulement turbulent sur quelques décimètres à la base (filtré par Cainocrinus et Conocrinus) et laminaire au-dessus (filtré par Isselicrinus sp. 3) ;
- • (c) les Isselicrinus sp. 1 (remaniés du niveau C2) filtrant un courant faible à modéré à écoulement laminaire, sans turbulence, permis par un fond vaseux à surface plane.
Selon les données biométriques (Fig. 4), Isselicrinus sp. 2 vivrait à une profondeur (au moins 400 m) nettement supérieure à celle des pentacrines du niveau C6a (200–300 m). En revanche, les Conocrinus, qui atteignent une taille maximale pour ce genre, évoquent une profondeur inférieure ou égale (entre 100 et 200 m) à celle du biotope à Isselicrinus sp. 5 et à grands Tormocrinus. Ils auraient pu vivre dans un biotope similaire à celui d'Isselicrinus sp. 2, mais nettement moins profond. Néanmoins, Isselicrinus sp. 2 et les grands Conocrinus ont pu appartenir à une même communauté, plus profonde, mais vivant dans un milieu riche en particules nutritives, du type des milieux qui se développent actuellement au niveau des monts sous-marins [14]. En effet, certaines columnales de grands Conocrinus montrent des signes de creusement des facettes articulaires qui ne prennent de l'importance chez les phénotypes analogues actuels qu'au-dessous de 400 m de profondeur.
La faune crinoïdique du niveau C4 et de l'association (b) du niveau C3 est très proche de celle des marnes de la Villa Marbella (Bartonien, Côte des Basques, Biarritz). Dans ce dernier site, l'horizon où les Isselicrinus didactylus abondent, tandis que les Conocrinus se raréfient (base des Marnes supérieures de la Villa Marbella), correspondrait à une profondeur de 500 m ou plus [17], mais ne dépassant pas 600 m [5]. En C4, les proportions sont inversées, avec des Conocrinus et Tormocrinus abondants et des pentacrines peu fréquentes, ce qui évoque une profondeur moindre qui, pour être cohérente avec les résultats de la Fig. 4, se situerait entre 300 et 400 m. L'association (b) du niveau C3 avec Isselicrinus sp. 3, phénotype plus proche d'I. subbasaltiformis (comme Isselicrinus sp. 5) que d'I. didactylus, se place ainsi naturellement dans la partie la plus profonde (aux environs de 300 m) du créneau bathymétrique d'Isselicrinus sp. 5. La présence d'Isselicrinus dans C5 en atteste le caractère épibathyal.
On aboutit ainsi à une reconstitution de la zonation bathymétrique du genre Isselicrinus à l'Éocène. L'écophénotype balanocrine est représenté par trois taxons se succédant sur le talus bathyal : I. subbasaltiformis (200–300 m), I. didactylus (300–600 m) et Isselicrinus sp. 1 (600–1000 m ou plus). L'écophénotype isocrine (I. diaboli sensu lato) apparaît ici dans le créneau bathymétrique d'I. didactylus quand un courant à écoulement turbulent domine.
4.2 Autres indices paléobathymétriques et taphonomiques
Les foraminifères benthiques fournissent de bons indices paléobathymétriques, en tenant compte du fait qu'ils peuvent être facilement déplacés. Ici, les estimations proposées découlent des principes discutés par Mathelin et Sztrákos [10] et Sztrákos et du Fornel [20]. Les échantillons (E) prélevés dans la marnière de Miretrain par Sztrákos et al. [21] ont été réexaminés ainsi que les niveaux à crinoïdes (C). Les échantillons E1 à E7, intervalle incluant C0 et C1, ont livré Nuttalides truempyi, classiquement considéré comme une forme bathyale profonde, indiquant une bathymétrie toujours supérieure à 500 m [11] et atteignant souvent 1000 m ou plus [9]. La présence de Cribroparella pteromphalia, Repmanina charoides et Bulimina trinitatensis de E8 à E11, niveaux sous-jacents à C2, évoque un milieu bathyal d'environ 500 à 600 m de profondeur. Le niveau C2 se situe principalement entre les échantillons E12 et E14. Les marnes situées 2 m sous la base de C3 nous ont livré Cribroparella pteromphalia, Repmanina charoides et Bulimina trinitatensis, et un exemplaire de Nuttalides truempyi. Bien que le seul exemplaire de cette dernière espèce puisse faire partie de rares éléments remaniés comme Acarinina wilcoxensis, Morozovella aragonensis (un exemplaire) et Ganella neumannae qui proviennent de niveaux plus anciens, la profondeur de dépôt tirée de l'analyse des échantillons E8 à E11 se maintient probablement jusqu'au sommet de C2.
Le dépôt gravitaire C3 se situe à proximité de l'échantillon E14. Il contient à la fois des espèces néritiques (notamment Stichocibicides cubensis, Neorotalia lecalvezae, Korobkovella sublobatula et des Discocyclines) et des espèces épibathyales comme Cribroparella pteromphalia. La coulée bioclastique C4, proche de E16, fournit une grande quantité d'espèces provenant du domaine néritique (notamment Angotia aquitanica, Eponides polygonus, Stichocibicides cubensis, des Nummulites et des Discocyclines). Situé entre E22 et E23, C5 contient l'espèce bathyale Bulimina trinitatensis et des espèces de plate-forme externe pouvant dépasser 100 m de profondeur, comme Martinottiella bignoti et M. cocoaensis. Bien qu'ensuite les sédiments se chargent en espèces néritiques, dont de nombreuses Nummulites et Discocyclines, et en matériel remanié, qui ne permet pas une estimation fiable du milieu de dépôt, la présence de Tritaxilina pupa, Uvigerina rippensis et U. acutocostata jusqu'au sommet de la zone à M. lehneri pourrait indiquer un milieu encore épibathyal.
Dès la base de la zone à T. rohri (C6a), l'ampleur des remaniements s'accroît brusquement. En plus de quartz bipyramidés triasiques, on y recueille des foraminifères provenant du Crétacé (Globotruncana sp., Pseudotextularia sp.), du Thanétien (Globanomalina chapmani) et de l'Éocène inférieur et moyen basal (Tritaxilina cubensis, Morozovella subbotinae, M. gracilis, M. caucasica, Ganella neumannae).
Dès la base des marnes gris foncé, les Lithothamniées deviennent abondantes, témoignant de la proximité de la zone euphotique. En revanche, elles sont absentes des marnes gris clair et notamment du dépôt gravitaire C3. Par ailleurs, des articles d'Isididae (grandes gorgones épibathyales) ont été récoltés dans C6a.
Dans des bancs de calcaire marneux bioclastique à cachet néritique (zone à T. rohri), qui couronnent la série au-dessus de C6, une riche faune d'échinides évoque un milieu infralittoral pouvant se situer vers 50 m de profondeur [15]. Ces auteurs notent que le plancton y est toujours bien représenté et que les interbancs marneux contiennent encore quelques ostracodes et crinoïdes d'affinités bathyales.
La faune de sélaciens de la marnière de Miretrain a été étudiée par Adnet [1], en portant une attention particulière aux taxons benthiques ou bathypélagiques et à leurs équivalents actuels. Dans C0, la majorité des sélaciens identifiés indique une profondeur d'au moins 300–400 m, et plus de 50%des taxons peuvent atteindre 1000 m, certains (Apristurus, Deania, Centrosoymnus) ayant actuellement leur optimum de fréquence entre 500 et 1000 m, ou un autre (Etmopterus) entre 400 et 700 m. Les taxons ayant leur optimum entre 0 et 100 m sont peu représentés, ce qui indique un relatif éloignement du domaine littoral. Les autres échantillons prélevés au sein des marnes gris clair supérieures, au-dessus de E15, contiennent une majorité de taxons, qui suggèrent une profondeur maximale passant de 500 m (vers C4) à 200 m (au sommet de la zone à M. lehneri) ; les taxons ayant leur optimum entre 400 et 1000 m restent présents, bien que peu fréquents.
Les crinoïdes suggèrent une profondeur du biotope plus forte (800 m) pour C2 que les foraminifères benthiques (500–600 m). Ces derniers pourraient avoir été déplacés, puisque de rares éléments incontestablement remaniés ont été détectés. Mais une certaine surestimation des écarts de profondeur séparant les biotopes à Isselicrinus ne peut être exclue. Pour tous les autres niveaux, les divers indices de profondeur sont compatibles.
4.3 Évolution des milieux de dépôt
Les divers indicateurs bathymétriques convergent pour confirmer que la série éocène de la carrière de Miretrain s'est déposée dans des milieux passant d'une profondeur pouvant atteindre le millier de mètres à la base à une tranche d'eau égale ou légèrement supérieure à 50 m, au niveau des calcaires bioclastiques sommitaux. Ils permettent de préciser les modalités de cette rapide réduction de profondeur (Fig. 5).
Les marnes gris clair inférieures correspondent à une période assez stable de haut niveau marin relatif, qui permet à la faune benthique de se développer et de s'étager sur le talus séparant Angoumé du haut-fond formé par l'anticlinal de Tercis. Ce dernier devait être immergé sous une tranche d'eau généralement supérieure à 100 m, comme l'atteste la rareté des restes des sélaciens littoraux.
Ensuite, une baisse du niveau marin relatif s'effectue en deux étapes. La première, probablement enclenchée par la surrection de l'anticlinal de Tercis, accentue et déstabilise la pente sédimentaire, provoquant des glissements en masse (C3, C4) qui amènent au pied du talus un mélange de matériel infralittoral et épibathyal et des fragments de silex remaniés du soubassement crétacé structurant le haut-fond. Le maximum de profondeur envisageable ne dépassera plus 400–500 m après le dépôt de C3 et l'importance relative des bioclastes provenant d'une bordure de plate-forme restera élevée. La seconde phase débute par l'enfouissement d'une faune de grands suspensivores épibathyaux (200–300 m) au sein d'un niveau (C6a) comportant à la fois les premiers bancs de calcaire marneux bioclastique et d'abondants éléments remaniés témoignant de la reprise de l'érosion des anticlinaux et diapirs voisins. Vers le sommet du talus, l'installation du mégabenthos et la richesse relative en carbonates évoquent un épisode de stabilité lié à une réduction de l'apport terrigène avant la brusque reprise d'un détritisme, souvent grossier, au sein des marnes gris foncé supérieures. La tendance à la dominante carbonatée reprend avec la mise en place des calcaires bioclastiques néritiques qui couronnent la série. Ce retour à un contexte plus stable se traduit par l'installation d'une faune d'échinides particulièrement diversifiée. La présence, tout au long de la série, des crinoïdes pédonculés, et notamment des pentacrines, est favorisée par la permanence de courants bathyaux, canalisés par la morphologie sous-marine.
Les hauts-fonds qui bordent le secteur d'Angoumé sont les structures diapiriques de Tercis et de Dax (Fig. 1). La réactivation de l'halocinèse par une phase tectonique, même faible, peut amplifier localement leur déformation, stimuler les dépôts gravitaires et perturber l'enregistrement sédimentaire de phénomènes régionaux ou plus globaux. L'importance de C3 n'a probablement qu'une signification locale, car ce niveau ne correspond pas à un événement régional identifié. En revanche, l'accentuation du détritisme accompagnée d'importants remaniements, qui débute avec C6a, est une des premières manifestations de la phase tectonique majeure de raccourcissement dans la zone sous-pyrénéenne, qui se développe durant le Bartonien supérieur et le Priabonien [6,13]. La précédente étape de déformation régionale date de l'Yprésien supérieur–Lutétien inférieur. Elle est soulignée par la discordance des marnes de Miretrain sur les terrains antérieurs.
5 Conclusions
La série éocène de la carrière de Miretrain constitue un exemple remarquable de progradation sédimentaire en domaine bathyal entre deux phases de déformation tectonique. Les crinoïdes pédonculés, les foraminifères benthiques et les sélaciens fournissent des indices bathymétriques convergents, permettant de suivre les étapes de la réduction de profondeur d'au moins 800 m qui s'ensuit. Cette réduction s'opère sur une dizaine de millions d'années et en 160 m de sédiments compactés sans discontinuité majeure observable. La situation du site sur le flanc nord de l'anticlinal diapirique de Tercis incite à considérer que la déformation locale du substratum y contribue pour plusieurs centaines de mètres, notamment au Bartonien supérieur, dans un contexte géodynamique régional du bassin de l'Adour où l'accommodation reste toujours inférieure à la sédimentation [19].
La diversité en crinoïdes pédonculés, exceptionnelle sur une même coupe, a permis de reconstituer une zonation bathymétrique des pentacrines éocènes du genre Isselicrinus cohérente avec la microfaune associée et les étagements connus dans l'océan actuel. Dans l'Éocène aquitain, nous disposons désormais d'une succession de crinoïdes à Angoumé, dans un contexte de réduction de la bathymétrie, et à Biarritz, dans un contexte d'approfondissement. Les caractères du pédoncule des pentacrines actuelles, liés à l'adaptation à la profondeur, peuvent être utilisés avec succès sur un genre uniquement fossile, comme Isselicrinus, pour estimer les paléobathymétries des milieux de vie. Les nouvelles conditions hydrologiques liées à la production d'eaux froides antarctiques qui se mettent en place dans l'océan à partir de l'Oligocène et singularisent l'océan actuel [2] ne sont donc pas un obstacle à l'application de cette méthode à des faunes plus anciennes.
Remerciements
Nous remercions S. Favin, F. Guy, A. Fuchs, D. Delors et X. Drothière pour leur contribution à la réalisation de cette étude.