

Abridged English version
The Fluminense coastline, between Niteroi and Cabo Frio (Brazil; 23°S–43°W) is located in the humid tropical zone, characterized by a wet season dominated by northeastern winds (NE) and by a dry season, with intermittent southern sector winds generated by polar advections. But near Cabo Frio, the occurrence of a local upwelling, mainly controlled by wind patterns, generates arid conditions. Under northeast winds effect, cold waters from the deep resurface, whereas winds from south prevent it. In South America, strong El Niño events lead to important rainfall anomalies and changes in wind patterns [5]. In the Cabo Frio area, they induce stronger aridity and northeast winds due to an increase of the low level jet [5].
The Fluminense coastline presents a lagoonal system (Fig. 1) composed of an internal Pleistocene part with large lagoons (e.g., Araruama) in connection with the Atlantic Ocean, and an external Holocene part (e.g., Brejo do Espinho) with smaller ones, isolated by sand barriers. Physical and chemical characteristics of the lagoons are primarily controlled by dry season duration, which is linked to the upwelling occurrence. The upwelling induces a reduction in local rainfall, an increase in evaporation and thus, in salinity. These conditions induce hypersaline lagoons with organic-carbonate sedimentation which are present in the area influenced by the upwelling (eastern part of the coast) and hyposaline lagoons with organic-detrital sedimentation in the west. Previous mineralogical, isotopic, and diatom studies evidence a high hydrological variability during the last 5000 yr [20–22]. A first sedimentary record collected in this lagoon highlighted three dry phases at 3600–3500, 3100–3000 and 2200–2000 yr [21]; another core showed a phase of water level variability from 4500 to 2200 yr, an increasing dryness after this date and dry phases at 4500 yr, between 3000 and 2400 yr (ca. ∼2600 yr) and at 2200 yr [20]. This article aims at characterizing high resolution hydrological variability in a core from Brejo do Espinho's lagoon (eastern part of the littoral) from the last 6500 yr by coupling organic matter study (TOC, HI, C/N), isotopic (∂13C, ∂15N) and diatoms proxies.
Localisation du sondage LBE02 dans la lagune de Brejo do Espinho (Brésil, 23°S, 43°W).
Location of the core LBE02 in Brejo do Espinho lagoon (Brazil, 23°S, 43°W).
A 244 cm core (LBE02), collected in the central part of Brejo do Espinho's lagoon shows continuous organic-carbonate sedimentation. Five AMS radiocarbon ages performed on organic sediments (TOC varying between 2 and 15%) date the core between 14C yr B.P. (42 cm), and 14C yr B.P. (232 cm) (Fig. 2). Sedimentation rates varied between 25 and 112 cm kyr−1. Total carbon organic (TOC, %) and Hydrogen index (HI, mg HC/g TOC) were obtained on 37 samples from Rock–Eval 6 pyrolysis (RE 6, turbo; ISTO, France). Isotopic (∂13C (‰), ∂15N (‰)) and C/N analyses were performed by mass spectrometry on 243 samples. Diatom analysis was applied on 72 samples; only 63 samples were considered for multivariate analyses (AFC, CAH, VARCAR using BIOMECO program) due to chemical dissolution of diatom frustules in sediments from 24 cm to the top and from 146 to 136 cm.
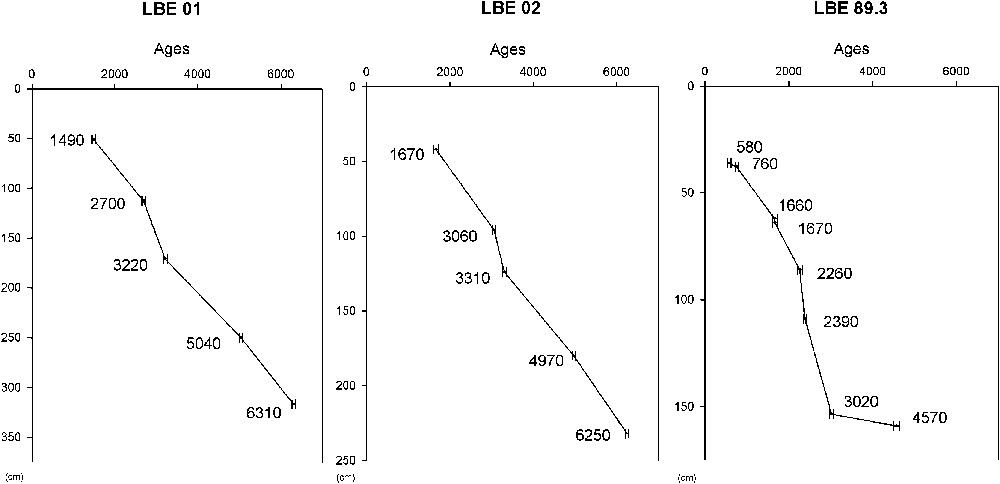
Modèles d'âges radiocarbone (14C ans B.P.) en fonction de la profondeur (cm) des sondages LBE01, LBE02 et LBE89.3.
Radiocarbon ages (14C yr B.P.) along the depth (cm) of LBE01, LBE02 and LBE89.3 cores.
The core begins by a first sequence [6500–2400 yr] consisting of an alternating organic and organic-carbonate sandy sedimentation (Fig. 3). From 6500 to 4000 yr the sequence is dominated by Achnanthes arenaria indicating shallow and mesosaline conditions. This phase is interrupted by (i) higher water level indicated by planktonic species and by (ii) low water level from 6200 to 5900 yr marked by sand and low amount of diatoms in the sediment accompanied by the decrease of other proxies (TOC, HI, ∂13C, ∂15N). It is also characterized by episodes (at 5500, 5200–5100, 4800–4600 and 4200–4000 yr) of more saline conditions (poly- to hypersaline) indicated by the occurrence of Amphora coffeaeformis and a decrease of planktonic species, diatoms amount and TOC values. From 4000 to 3700 yr, only few dissolved diatoms were observed simultaneously with organic sediments and ∂13C decrease. At 3700 yr, diatoms reappeared in the sediments indicating shallow poly- to hypersaline conditions associated with high HI values up to 600 mg HC/g TOC. From 3600 to 2600 yr, a phase dominated by Amphora coffeaeformis is interrupted by some levels marked by planktonic species increase (e.g., 2900 to 2600 yr). From 2600 to 2400 yr, Cocconeis neodiminuta dominates, indicating oligosaline to meso-polysaline conditions when associated to Amphora coffeaeformis. These conditions are accompanied by ∂13C values between −22 and and HI values up to 600 mg HC/g TOC suggesting the presence of well preserved algal organic matter.
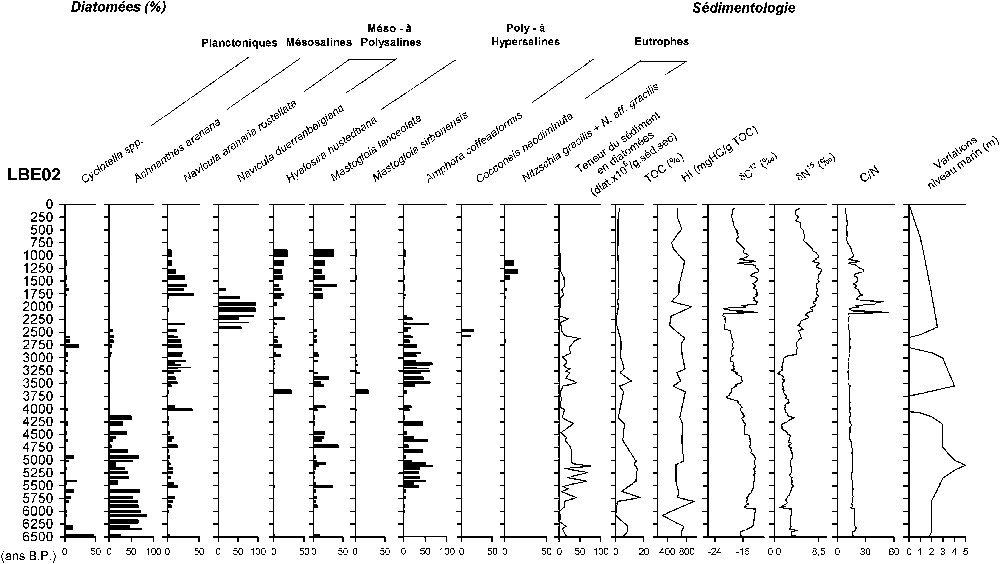
Abondance relative (%) des diatomées dominantes, teneur du sédiment en diatomées (nbre diat. 105/g. sédt. sec), taux en carbone organique (TOC, %), indice d'hydrogène (HI, mg HC/g. TOC), ∂13C (‰), ∂15N (‰), rapport C/N, variations du niveau marin (m) en fonction des âges.
Diatom summary: relative abundance (%) of dominant, diatom content for each sample (nbr diat. 105/g. dry sedt), organic carbon content (TOC, %), hydrogen index (HI, mg HC/g. TOC), C/N, ∂13C (‰), ∂15N (‰), sea-level variability (m) along the age.
The second sequence [2400–0 yr] is characterized by homogenous carbonate sedimentation associated with a decrease of diatoms amount and TOC content while ∂13C values and C/N ratio increase. From 2400 to 1800 yr, the lagoon records a high variability marked by successive meso- to polysaline and poly- to hypersaline conditions. Dilution episodes (at 2300–2250, 2200–1950 yr) are indicated by ∂13C values () and C/N ratios (<10). From 1800 to 950 yr, a salinity increase is evidenced by hypersaline species occurrence (Mastogloia lanceolata, Hyalosira hustedtiana). This phase is interrupted from 1300 to 1150 yr by eutrophic and mesosaline conditions as shown by an increase of ∂15N (), C/N (>20) and HI (>600 mg HC/g TOC) values. To the top, only few diatom ghosts were observed while HI is over 600 mg HC/g TOC and isotopic proxies decrease.
These results, from LBE02 core study, collected in the centre of Brejo do Espinho's lagoon, highlight a strong hydrological variability between 6500 and 2400 yr, whereas hypersaline conditions seem to take hold from 2400 yr.
In this lagoonal context, interpretation of hydrological variability has to take into account lagoon's morphology and record site of the core. Results are compared with two other cores, LBE89.2 and LBE89.3 collected in Brejo do Espinho's lagoon and covering respectively 6600 and 4500 yr [20,21]. This interpretation has also to consider sea level variations. A sea level reconstruction is available [22] (Fig. 3). It has been established from fossil beach ridges geometry observed to an extent of 3300 km along the Brazilian coast [22]. It is based on more than 700 radiocarbon ages.
From 6500 to 5100 yr, a phase of high water level is recorded in LBE02 by the presence of planktonic species while sea level rises to reach a maximum at 5100 yr. At that time, the lagoonal system is isolated from the Atlantic Ocean. This phase would be explained by the marine transgression. The two episodes of low water level observed at 5500 and 5200–5100 yr would correspond to dry climatic episodes.
From 5100 to 3900 yr, sea-level slopes down, provoking internal and external system division [22]. This suggests that higher water level recorded in LBE02 at 5500–5200, 5100–4800 and 4600–4200 yr would be related to meteoric inputs. At 4000 yr, diatom's lack would result from the lagoon confinement as a consequence of the sea level drop inducing salinity increase and frustule dissolution. It is in agreement with the dry phase recorded from 4500 to 3020 yr in LBE89.3 core [20]. Located on the margin of the lagoon, LBE89.3 recorded a long dry phase while LBE02 recorded a stronger variability suggesting a partial dry phase.
From 3900 to 3600 yr, the Fluminense coastline knows a marine transgression responsible in LBE02 at 3800–3700 yr for diatoms reappearance associated to ∂13C and HI values characteristics of an algal development [4,6,13].
From 3600 to 2900 yr, LBE02 is characterized by meso- to hypersaline conditions alternation while the sea level remains high till 2900 yr. These hypersaline conditions marking frequent dry episodes would be reinforced by the definitive isolation of the lagoon at 3450 yr [22]. This phase of strong variability is recorded in LBE89.2 from 3600 to 3000 yr (e.g., laminated sedimentation) flanked by two dry episodes at 3600–3500 yr and 3100–3000 yr [21], which would be only partial ones.
From 2900 to 2600 yr, the lagoon is characterized by a high water level while the marine transgression has stopped. The last transgression (2700–2500 yr) keeps this water level high. These conditions promote the increase of algal development marked in LBE02, from 2600 to 2400 yr, by epiphytic diatoms and ∂13C and HI values. Thus, the dry phase recorded between 3000 and 2400 yr (ca. ∼2600 yr) in LBE89.3 [20] would correspond to a partial dry episode resulting from eutrophication of the lagoon.
From 2500 yr, sea level slopes down progressively to reach its mean present level inducing a climatic origin to hydrological variability recorded in the lagoon.
From 2400 to 1800 yr, a succession of dilution/salinisation phases corresponds with sedimentation change from organic-carbonated to carbonated in LBE02. This change is recorded at 2200 yr in LBE89.2 and LBE89.3 consisting in a partial dry phase respectively at 2200–2000 yr and at 2200 yr [20,21].
From 1800 yr, increasing hypersaline conditions establishment traduces the growing confining of the lagoon due to an increasing aridity also observed in LBE89.3 [20]. In LBE02, ∂15N increase results from the progressive eutrophication of the environment [2] corroborated by eutrophic species presence and by a C/N ratio high traducing a strong productivity. Diatom's lack at the top of the core suggests salinity conditions provoking a chemical dissolution of diatoms frustules.
The study of LBE02 core allows completing data established by previous study by highlighting a phase dominated by marine inputs from 6500 to 5100 yr, frequent wet climatic episodes from 5100 to 4000 yr, a phase of lagoon eutrophication from 2600 to 2400 yr and a succession of wet and dry climatic episodes from 2400 to 1800 yr. It corroborates dry phases recorded in other cores [4500, 3600–3500, 3100–3000, ca. ∼2600 and 2200 yr] by establishing that it consists in partial drying of the lagoon and shows two new arid episodes at 5500 and 5200–5100 yr. Finally, it confirms the increase of aridity observed in previous cores from 2200 yr and observed in LBE02 from 1800 yr.
Dryness intensity in this lagoonal context of the Fluminense coastline is controlled by the Cabo Frio's upwelling occurrence generated by northeast winds. Thus, dry phases recorded in the Brejo do Espinho's lagoon would be combined with an increase of the upwelling activity related to stronger northeast winds. These results confirm, before 2400 yr, frequent humid climatic events would be connected to south–southwest winds predominance; after this date, northeast winds would be stronger [20]. Furthermore, phases of prolonged Cabo Frio's upwelling would be generated by strong El Niño events [12], implying that dry phases identified in the lagoon would correspond to strong El Niño phases. Thus, these results indicate El Niño events were infrequent before 4000 yr, frequent between 3600 and 2900 yr and 2400 and 1800 yr and very frequent from this date. Increase of El Niño phases activity during Late Holocene is in agreement with ENSO records from numerous sites of South America [3,17], with recent El Niño activity for 5000 yr [15], El Niño events infrequent between 6100 and 4000 yr, and intensity and frequency enhancement from 3000 yr [14].
This work continues with the study of a marine core collected off Cabo Frio to obtain a chronology of the upwelling. Likewise, to determine El Niño events intensity, a core has been collected in a lagoon from the west of the Fluminense coastline, where only upwelling phases of strong intensity are recorded.
1 Introduction
Le littoral Fluminense, de direction ouest–est entre Niteroi et Cabo Frio (Brésil ; 23°S–43°W), est situé en zone tropicale humide. Le climat est caractérisé par un été humide dominé par les vents de nord-est (NE) et un hiver sec soumis aux vents de sud/sud-ouest (S/SO) généré par les advections polaires [1]. Pourtant, la région de Cabo Frio présente un microclimat semi-aride, généré par un upwelling local, principalement contrôlé par le régime des vents. Lorsque les vents de NE repoussent les eaux chaudes côtières vers le large, les eaux froides du plateau continental remontent à la surface, entraînant localement une accélération du mouvement de subsidence de l'air refroidi, responsable de l'aridité. Au contraire, les vents du S–SO empêchent cette remontée en maintenant les eaux chaudes vers la côte. En Amérique du Sud tropicale, les forts événements El Niño entraînent d'importantes anomalies du régime des pluies et des vents [5]. Dans la région de Cabo Frio, cela se traduit par une augmentation de l'aridité et des vents de NE, due à un renforcement du jet tropical [5].
Le littoral Fluminense est constitué d'un système lagunaire, composé d'une partie pléistocène interne formée de grandes lagunes en communication avec l'océan Atlantique (par exemple, Araruama) et d'une partie holocène externe occupée par de petites lagunes (ex. : Brejo do Espinho) isolées par des cordons dunaires (Fig. 1). Les caractéristiques physico-chimiques des lagunes sont contrôlées par la durée de la saison sèche liée à l'intermittence de l'upwelling. Le renforcement de l'upwelling provoque une diminution des précipitations, une augmentation de l'évaporation et de la salinité. Ainsi, les lagunes sont hypersalines, avec une sédimentation carbonatée dans la zone d'influence de l'upwelling à l'est du littoral, et hyposalines, organodétritiques à l'ouest.
Des travaux antérieurs portant sur la caractérisation minéralogique et isotopique des sédiments ainsi que sur l'étude des diatomées, ont mis en évidence des fluctuations hydrologiques, en relation avec des modifications climatiques enregistrées à l'échelle des latitudes tropicales du continent sud-américain au cours de l'Holocène [20–22]. L'étude d'un premier enregistrement sédimentaire prélevé dans la lagune a mis en évidence trois phases d'assèchement à 3600–3500, 3100–3000, 2200–2000 ans [21] ; un autre sondage a établi une phase de variabilité du niveau d'eau de 4500 à 2200 ans, des conditions arides croissantes après cette date et des phases d'assèchement de la lagune à 4500 ans, entre 3000 et 2400 ans (ca. ∼2600 ans) et à 2200 ans [20]. Le présent article contribue à la caractérisation à haute résolution de la variabilité hydrologique de ces milieux lagunaires depuis ∼6500 ans, en couplant la composition spécifique des assemblages de diatomées à des données sur la composition organique et isotopique des sédiments (TOC, HI, C/N, ∂13C, ∂15N).
2 Matériel et méthodes
La lagune hypersaline de Brejo do Espinho est située à l'est du littoral Fluminense (22°56′S, 42°14′W ; Fig. 1). Un sondage de 244 cm (LBE02), prélevé en 2000, a fait l'objet de cinq datations, réalisées sur des sédiments organiques en spectrométrie de masse par accélérateur (AMS) à Beta Analytic Inc. (États-Unis). Les âges cités sont exprimés en « 14C ans BP » et notés dans le texte « ans ».
Le taux de carbone organique total (TOC,%) et l'indice d'hydrogène (HI, mg HC/g. TOC) ont été obtenus sur 37 échantillons par pyrolyse Rock–Eval (Rock Eval 6 version turbo, institut des sciences de la Terre d'Orléans, France). La composition isotopique en ∂13C (‰), ∂15N (‰) et les rapports C/N ont été mesurés en continu (243 échantillons) par spectrométrie de masse à l'Environmental Isotope Laboratory de l'université de Waterloo (Canada).
La teneur du sédiment en diatomées (nbre diat./g. sédt. sec) a été mesurée sur 72 échantillons au microscope inversé Zeiss-Axiovert 25 [11]. La composition spécifique des diatomées a été déterminée en comptant un minimum de 400 valves au microscope Nachet NS 400, sur ces échantillons préalablement traités à l'H2O2 30% et à l'HCl 30% et montés entre lame et lamelle à l'aide de Naphrax. La détermination taxonomique est basée sur la classification de Krammer et Lange-Bertalot (1986–1991) [7–10], avec la prise en compte de la nomenclature de Round et al. (1990) [16]. L'interprétation écologique s'appuie sur de nombreux ouvrages [7–10,18,23] et sur une étude des diatomées actuelles de la lagune d'Araruama (au nord de Brejo do Espinho ; Fig. 1) [19]. Des analyses statistiques multivariées (analyse factorielle des correspondances [AFC], classification ascendante hiérarchique [CAH] avec le logiciel Biomeco) ont été appliquées aux données de comptage, afin d'obtenir des classes d'échantillons dont les espèces caractéristiques ont été mises en évidence par le programme Varcar (comm. pers. M. Roux).
3 Résultats
Le sondage LBE02 présente deux séquences sédimentaires : la première, de la base à 71 cm, caractérisée par une sédimentation hétérogène organo-carbonatée sableuse et la seconde, de 71 cm au sommet, par une sédimentation homogène carbonatée.
Il est daté à la base (232–234 cm) à ans et au sommet (42–45 cm) à ans. Il présente des taux de sédimentation variant de 25 à 112 cm ka−1. Les datations sont en accord avec la chronologie réalisée sur une carotte jumelle, LBE01, et sur une carotte de bordure de la lagune (LBE89.3 ; Fig. 2). On observe une accélération du taux de sédimentation dans LBE01 et LBE02, entre 2700 et 3000 ans, respectivement de 111 et 112 cm ka−1, dans LBE89.3 ; cette accélération apparaît entre 2260 et 2390 ans, avec un taux de 176 cm ka−1. L'effet réservoir évalué sur les âges mesurés sur LBE89.3 est considéré comme négligeable [18].
La flore des diatomées, composée de 103 espèces réparties en 35 genres, est dominée par des formes benthiques mésosalines à hypersalines et comprend trois espèces planctoniques, mais aucune d'eau douce (<1 g. l−1). Sur les 72 échantillons observés, 63 seulement ont fait l'objet d'analyses statistiques, du fait d'une quantité insuffisante de valves, fortement altérées chimiquement, dans les sédiments de 0 à 24 cm et de 146 à 136 cm. La CAH met en évidence neuf classes (Tableau 1).
Résultats des analyses statistiques multivariées : (a) numéro de la classe ; (b) profondeur des échantillons compris dans chaque classe ; (c) espèces de diatomées dominantes, sous-dominantes et accompagnatrices, définies par le programme VARCAR
Results of multivariate analyses: (a) number of sample group; (b) depth of samples included in each group; (c) dominant, subdominant and associated diatom species defined by the VARCAR program
| Classe | Echantillons (cm) | Espèces dominantes | Espèces sous-dominantes | Espèces accompagnatrices |
| 1 | 24, 36, 40, 42, 47, 136 | Navicula arenaria rostellata | Hyalosira hustedtiana | Fallacia litoricola Frustulia aff. isabelae Mastogloia lanceolata |
| 2 | 29, 33 | Amphora lineolata | Mastogloia lanceolata Hyalosira hustedtiana Nitzschia gracilis | Navicula arenaria rostellata Fallacia litoricola |
| 3 | 153, 157, 163, 175, 179, 182, 188, 191, 195, 198, 200, 206, 211, 214, 218, 222, 225, 230, 234, 236, 241 | Achnanthes arenaria | Amphora coffeaeformis Cyclotella striata | Cocconeis placentula euglypta Fragilaria sp. Navicula arenaria rostellata |
| 4 | 69, 78, 81, 85, 90, 92, 97, 102, 106, 111, 113, 118, 122, 127, 130, 132, 146, 148, 166, 168, 172, 184, 186, 202 | Amphora coffeaeformis | Navicula arenaria rostellata Mastogloia lanceolata | Frustulia aff. isabelae Achnanthes arenaria Amphora acutiuscula Paralia sulcata |
| 5 | 244 | Amphora interrupta Achnanthes arenaria | Cyclotella striata Mastogloia lanceolata | Opephora olsenii |
| 6 | 49, 65 | Navicula duerrenbergiana | Hyalosira hustedtiana Amphora coffeaeformis Mastogloia lanceolata | |
| 7 | 54, 58, 63, 68, 72 | Navicula duerrenbergiana | Amphora coffeaeformis | |
| 8 | 74 | Cocconeis neodiminuta | Navicula arenaria rostellata Gyrosigma balticum Rhopalodia accuminata | Amphora coffeaeformis Mastogloia lanceolata |
| 9 | 77 | Amphora coffeaeformis Cocconeis neodiminuta | Navicula arenaria rostellata | Gyrosigma balticum Cocconeis placentula euglypta Frustulia aff. isabelae Mastogloia lanceolata |
L'enregistrement débute par une phase [6500–5500 ans (240–202 cm)] (Fig. 3) dominée par Achnanthes arenaria (classe 3, mésosaline peu profonde), et ponctuée par des épisodes
- (i) d'augmentation du plan d'eau en relation avec la présence d'espèces planctoniques (Cyclotella striata) ;
- (ii) d'abaissement du plan d'eau, avec de 6200 à 5900 ans (230–220 cm), un niveau sableux associé à une diminution de la teneur en diatomées (16 à ), du TOC (9 à 0%), HI (700 à 225 mg HC/g. TOC), ∂13C (−12 à ) et ∂15N (3,3 à 1‰). De 5500 à 4000 ans (202–146 cm), une forte variabilité est enregistrée : des épisodes dominés par Amphora coffeaeformis (classe 4 ; poly- à hypersaline) apparaissent à 5500 ans (202 cm), 5200–5100 ans (186–184 cm), 4800–4600 ans (172–166 cm), 4200–4000 ans (148–146 cm), s'accompagnant d'une diminution des espèces planctoniques (<1%), de la teneur du sédiment en diatomées () et du TOC (<5%). À partir de 4800 ans (173 cm), cette variabilité est associée à une sédimentation laminée.
De 4000 à 3700 ans (146–136 cm), seuls quelques fantômes de diatomées fortement altérés sont observés, associés à des sédiments organiques et une diminution du ∂13C de −16 à .
À 3700 ans (136 cm), les diatomées réapparaissent dominées par l'espèce méso- à polysaline (Navicula arenaria rostellata, classe 1), accompagnée d'espèces poly- à hypersalines (Mastogloia lanceolata, M. sirbonensis, Hyalosira hustedtiana), marquant des conditions peu profondes associées à des valeurs en HI > 600 mg HC/g. TOC.
De 3600 à 2600 ans (132–78 cm), Amphora coffeaeformis (classe 4) domine, caractérisant des conditions poly- à hypersalines peu profondes, associées à des valeurs stables du TOC (4%), HI (500 mg HC/g. TOC), ∂13C (19‰), ∂15N () et C/N (12). Cette phase est jalonnée d'épisodes d'élévation du plan d'eau, indiqués par des augmentations de la teneur du sédiment en diatomées (10 à ) et des espèces planctoniques (Cyclotella spp ; 1 à 22%), en particulier de 2900 à 2600 ans (91–78 cm).
De 2600 à 2400 ans (78–72 cm), l'espèce épiphyte oligosaline, Cocconeis neodiminuta (classe 8) domine. Associée à Amphora coffeaeformis (classe 9), elle marque un milieu peu profond. Cet épisode est caractérisé par des valeurs du ∂13C comprises entre −22 et et en HI > 600 mg HC/g. TOC, indiquant la présence de matière organique algale bien préservée.
De 2400 ans à l'Actuel (72–0 cm), la seconde séquence est caractérisée par une sédimentation carbonatée, une diminution de la teneur du sédiment en diatomées (13 à ) et du TOC (<1%), alors que les indicateurs isotopiques augmentent (∂13C de −21 à , ∂15N de 5 à 9‰, C/N de 11 à 19). De 2400 à 1800 ans (72–49 cm), les sédiments laminés carbonatés enregistrent une forte variabilité caractérisée par une alternance de phases de dilution/salinisation, indiquée par l'espèce marine, mésosaline, Navicula duerrenbergiana, associée ou non à Amphora coffeaeformis (classe 6 ou 7). Les épisodes de dilution à 2300–2250 ans (68–65 cm) et 2200–1950 ans (63–54 cm) s'accompagnent de valeurs du ∂13C comprises entre −22 et et de valeurs de C/N < 10.
De 1800 à 950 ans (47–36 cm), les conditions deviennent hypersalines, avec une augmentation de Mastogloia lanceolata et de Hyalosira hustedtiana (classe 1). Cette phase est interrompue, de 1300 à 1150 ans (33–29 cm), par un épisode dominé par Amphora lineolata (classe 2, holo-eurysaline à polysaline), associée à Nitzschia gracilis (mésosaline eutrophe) et une augmentation du ∂15N (), du C/N (>20) et du HI (>600 mg HC/g. TOC). Au sommet de la carotte, on n'observe plus que quelques fantômes de diatomées, alors que les valeurs d'HI sont > 600 mg HC/g. TOC et que les paramètres isotopiques diminuent, indiquant une moins bonne préservation de la matière organique.
4 Discussion
Ces résultats, obtenus à partir de l'étude du sondage LBE02, prélevé dans le centre de la lagune de Brejo do Espinho, mettent en évidence une très forte variabilité hydrologique entre 6500 et 2400 ans, alors que des conditions hypersalines semblent s'installer à partir de 2400 ans.
Dans ce contexte lagunaire, l'interprétation de la variabilité hydrologique doit tenir compte de la morphologie de la lagune et du lieu d'enregistrement de la carotte. Pour cela, les résultats sont comparés à deux autres sondages, LBE89.2 et LBE89.3, prélevés dans la lagune de Brejo do Espinho et couvrant respectivement 6600 et 4500 ans [20,21]. En outre, cette interprétation doit également considérer les variations du niveau de la mer. Une reconstitution des variations du niveau marin est disponible [22] (Fig. 3). Elle a été établie à partir de la géométrie des cordons littoraux fossiles, observés sur une distance de 3300 km le long de la côte brésilienne. Elle est basée sur plus de 700 datations radiocarbones [22].
De 6500 à 5100 ans, une phase de haut niveau d'eau est enregistrée dans LBE02 par la présence d'espèces planctoniques, alors que le niveau marin s'élève pour atteindre un maximum à 5100 ans, et que le système lagunaire, interne (Brejo do Espinho) et externe (Araruama), est isolé de l'océan Atlantique. Cette phase serait expliquée par la transgression marine. En revanche, les deux épisodes d'abaissement du plan d'eau observés à 5500 et 5200–5100 ans correspondraient à des épisodes climatiques arides.
De 5100 à 3900 ans, le niveau marin s'abaisse provoquant la séparation des systèmes interne et externe [22]. Cela suggère que les élévations du plan d'eau enregistrées dans LBE02 à 5500–5200, 5100–4800 et 4600–4200 ans seraient liées à des apports météoriques. À 4000 ans, l'absence de diatomées résulterait d'un confinement de la lagune par suite de l'abaissement du niveau marin, entraînant une augmentation de la salinité et donc une dissolution des frustules. Ceci est en accord avec la phase d'assèchement enregistrée de 4500 à 3020 ans dans le sondage LBE89.3 [20]. Situé en bordure de lagune, LBE89.3 a enregistré une longue phase d'assèchement, alors que LBE02 a enregistré une plus forte variabilité, suggérant que la lagune n'a connu qu'un assèchement partiel.
De 3900 à 3600 ans, le littoral Fluminense connaît une phase de transgression marine qui serait responsable, dans LBE 02, à 3800–3700 ans, de la réapparition des diatomées, associée à des valeurs du ∂13C et de l'HI caractéristiques d'un développement algaire [4,6,13].
De 3600 à 2900 ans, LBE02 est caractérisé par une alternance de conditions méso- à hypersalines, alors que le niveau marin reste élevé jusqu'à 2900 ans. Ces conditions hypersalines marquant des épisodes arides fréquents seraient renforcées par l'isolement définitif de la lagune à 3450 ans [22]. Cette phase de forte variabilité est enregistrée dans LBE89.2 de 3600 à 3000 ans (i.e. : sédimentation laminée) encadrée par deux épisodes d'assèchement à 3600–3500 et 3100–3000 ans [21] qui ne seraient donc que partiels.
De 2900 à 2600 ans, la lagune est caractérisée par une élévation de son plan d'eau, alors que la transgression marine est interrompue. La dernière transgression marine (2700–2500 ans) maintient ce plan d'eau. Ces conditions favorisent l'accroissement du développement algaire marqué dans LBE02, de 2600 à 2400 ans, par des diatomées épiphytes et les valeurs du ∂13C et du HI. Ainsi, la phase d'assèchement enregistrée entre 3000 et 2400 ans (ca. ∼2600 ans) dans LBE89.3 [20] correspondrait à un assèchement partiel de la lagune résultant de l'eutrophisation de la lagune.
Après 2500 ans, le niveau marin s'abaisse progressivement pour atteindre son niveau moyen actuel, induisant une origine climatique des variations hydrologiques enregistrées dans la lagune.
De 2400 à 1800 ans, une succession de phases de dilution/salinisation de la lagune traduit le changement de sédimentation organo-carbonaté à carbonaté dans LBE02. Ce changement est enregistré à 2200 ans dans LBE89.2 et LBE89.3 et se manifeste par une phase d'assèchement partiel de la lagune respectivement à 2200–2000 ans et à 2200 ans [20,21].
À partir de 1800 ans, l'établissement de conditions hypersalines croissantes traduit le confinement croissant de la lagune, dû à une aridité croissante également observée dans LBE89.3 [20]. Dans LBE02, l'augmentation du ∂15N résulte d'une eutrophisation progressive du milieu [2], corroborée par la présence d'espèces eutrophes et un rapport C/N élevé traduisant une forte productivité du milieu. L'absence de diatomées au sommet du sondage suggère que les conditions de salinité ont provoqué une dissolution chimique de leurs frustules.
L'étude du sondage LBE02 permet donc de compléter les données établies par les précédentes études, en mettant en évidence une phase dominée par les apports marins de 6500 à 5100, des épisodes climatiques humides fréquents de 5100 à 4000 ans, une phase d'eutrophisation de la lagune de 2600 à 2400 ans et une succession d'épisodes climatiques humides et secs de 2400 à 1800 ans. Elle corrobore les phases d'assèchement enregistrées dans les autres sondages (4500, 360–3500, 3100–3000, ca. ∼2600 et 2200 ans) en établissant qu'il s'agit d'assèchements partiels de la lagune et met en évidence deux nouveaux épisodes arides à 5500 et 5200–5100 ans. Enfin, elle confirme l'augmentation de l'aridité observée dans les précédents sondages à partir de 2200 ans et observée dans LBE02 à partir de 1800 ans.
5 Conclusion
L'intensité de l'aridité dans ce contexte lagunaire du littoral Fluminense est contrôlée par la présence de l'upwelling de Cabo Frio généré par les vents de NE. Les phases sèches enregistrées par la lagune de Brejo do Espinho seraient donc associées à une augmentation de l'activité de l'upwelling en relation avec une augmentation des vents de NE. Ainsi, ces résultats confirment que, avant 2400 ans, des événements climatiques humides fréquents seraient liés à une prédominance des vents de S–SO ; après cette date, on assisterait à un renforcement des vents de NE [20]. De plus, des phases d'upwelling de Cabo Frio prolongées seraient générées par de forts événements El Niño [12] impliquant que les phases arides enregistrées dans la lagune correspondraient à des phases de forts El Niño. Cette étude indiquerait donc des événements El Niño peu fréquents avant 4000 ans, fréquents entre 3600 et 2900 ans et entre 2400 et 1800 ans et très fréquents depuis 1800 ans. L'augmentation d'une activité des phases El Niño durant l'Holocène tardif, est en accord avec des enregistrements des phénomènes ENSO observés dans de nombreux sites d'Amérique du Sud [3,17], avec une mise en place de l'activité El Niño actuelle après 5000 ans [15], des événements El Niño présents mais peu fréquents entre 6100 et 4000 ans, et une intensité et une fréquence accrues depuis 3000 ans [14].
Ce travail est poursuivi par l'étude d'un sondage marin prélevé au large de Cabo Frio afin d'obtenir une chronologie de l'upwelling. De même, dans la perspective de déterminer l'intensité des événements El Niño, un sondage a été prélevé dans une lagune à l'ouest du littoral Fluminense, où seules les phases d'upwelling de fortes intensités sont enregistrées.
Remerciements
Ce travail a été financé par le programme national Eclipse du CNRS. Nous remercions l'université Federal Fluminense de Niteroi (Brésil) pour son aide logistique sur le terrain et Didier Keravis de l'institut des sciences de la Terre d'Orléans (France) pour son soutien concernant les analyses Rock–Eval.