

1 Introduction
The aim of this article is to review the history of desertification in Africa, based on the palaeontological, sedimentological and stable isotope evidence obtained over the past 20 years in southern, central, northern and eastern Africa. The morphofunctional data yielded by the faunas (mode of locomotion, dietary adaptations such as hypsodonty, brachyodonty) provide evidence for the changes in the environment, which can be interpreted in terms of climatic changes.
At present, desert covers approximately one third of the surface of Africa, the Sahara in the north extending over 9 million km2, the Namib ca 135,000 km2, the Kalahari some 260,000 km2, with other smaller desert zones in Kenya, Somalia, Eritrea and Ethiopia and a large desert in neighbouring Arabia. The Neogene and Quaternary were marked by a succession of changes which affected the climate and played an important role in modifying African floras and faunas. Geological and palaeontological studies in areas that, at present, are total desert, such as Gebel Bellorat in the Western Desert, Egypt, Sahabi and Gebel Zelten in Libya, the rift valley deserts of northern Kenya, Ethiopia and Eritrea, and the Namib Desert in Namibia, reveal that during the Neogene all these areas were for a time more humid than they are today, and some areas such as the Congo Basin which, today, are very humid were drier during the Miocene (Fig. 1).
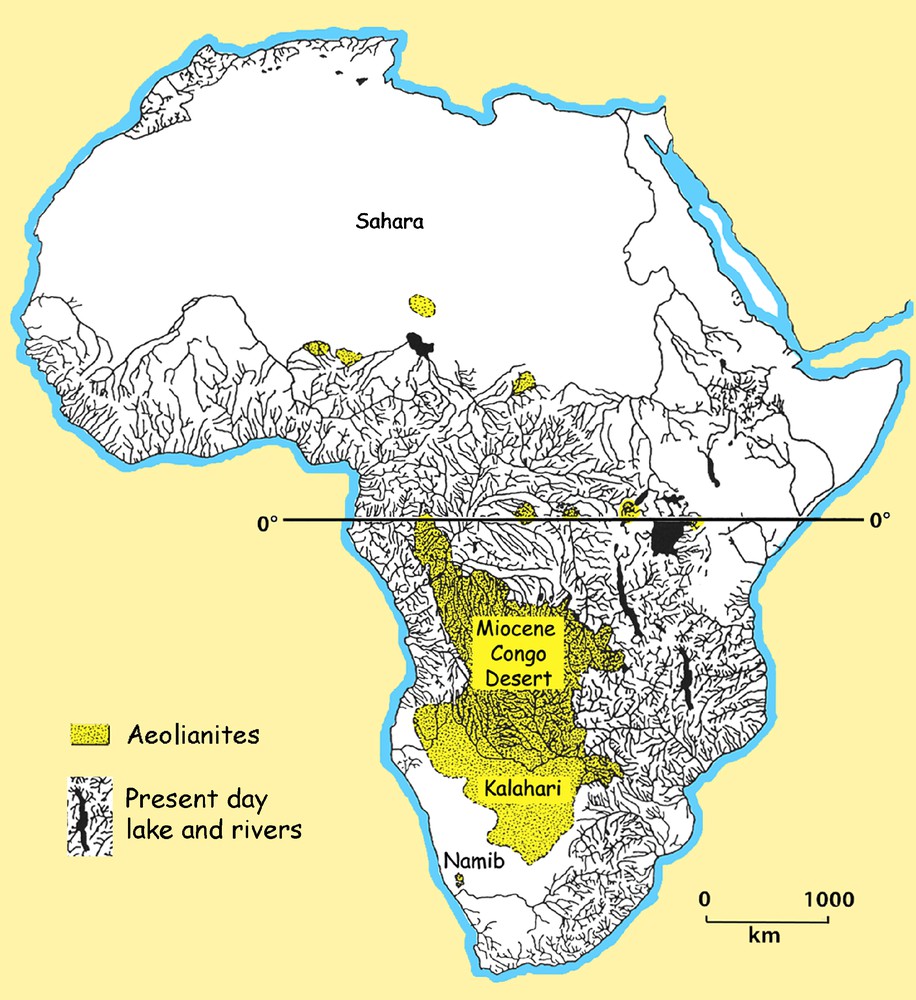
Distribution of stabilised dune sands in Africa. These indicate that areas that are now among the wettest in the continent (shown by the present-day permanent rivers) used to be arid to hyperarid during the past.
Fig. 1. Répartition des dunes sableuses fixées au Miocène en Afrique. Elles indiquent que des régions qui sont aujourd’hui parmi les plus humides du continent (représentées par les rivières permanentes actuelles), étaient arides à hyperarides dans le passé.
2 Southern Africa
Early Miocene sediments at Ryskop (South Africa) have yielded a faunal assemblage which, despite its fragmentary nature, suggests that the climate of Namaqualand 18 Ma ago was tropical to subtropical and differed strongly from the present temperate situation with winter rainfall [42]. In the northern Sperrgebiet (Namibia) [38], faunal rich deposits indicate the presence of woodland to open forest conditions from 21 to 19 Ma, after which the fauna changed to steppic and arid types [23,25,32,33]. In the lower Middle Miocene (17.5 Ma ago), along the Oranje river, there was a large diversity of mammals and numerous taxa such as Hyracoidea, Rhinocerotidae, Macroscelidea and Pedetidae which suggest a bushy savannah and such as Afromastodon coppensi, Prodeinotherium hobleyi and Nguruwe kijivium which indicate more or less wooded savannah. All Arrisdrift micromammals are terrestrial or subterranean, and none of them are arboreal or climbers. The faunal association indicates the presence of a gallery-forest, and the occurrence of crocodiles (Crocodylus gariepensis) and giant tortoises suggest a warm to subhumid climate. In Namaqualand, the younger site of Bosluis Pan (15–16 Ma) [41] has yielded a fauna which, despite its limited diversity provides information on the Miocene environments. The conditions were more tropical until the base of the Middle Miocene (Fig. 3) as shown by the crocodiles. A riparian woodland or forest in a regionally semi-arid environment agrees with the presence of the hypsodont macroscelidid, Myohyrax, of a giraffid, a bunodont gomphothere, the hypsodont Choerolophodon pygmaeus [21] and a brachyodont bovid. These data support the results from palaeobotanical studies at Brandvlei (Northern Cape Province) which show that the area was well wooded with a tropical to subtropical type of vegetation [9].
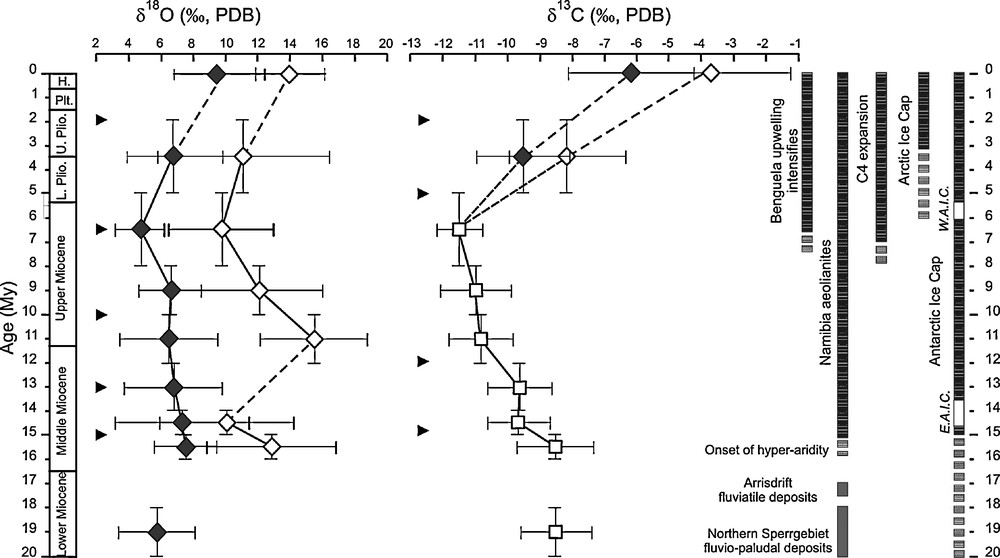
Trends in δ 18O, δ13C of Namib ratite eggshells during the Neogene. Black diamonds: southern Namib; open diamonds: Central Namib (from [33]).
Fig. 3. Évolution des variations dans les δ 18O, δ13C de coquilles de ratites du Namib au cours du Néogène. Diamants noirs : Namib du Sud ; diamants ouverts : Namib du Centre (d’après [33]).
In contrast, slightly younger deposits in the Namib Desert (ranging from Middle Miocene to Holocene on the basis of fossil mammalian chronology) consist of fossil dunes (aeolianites) which are endowed with a diverse fossil fauna including ratite eggshells which form the basis of the first biochronological scale for the Neogene of Southern Africa [26,35,40] (Fig. 2). No two types of eggshells have ever been found together in the same level, they are always found in superposition. In several levels, the eggshells are associated with a mammalian assemblage (micro- and macrofauna) which permits the calibration of the sequence by comparisons with other sequences in Africa, which have been dated by radiometric techniques. These aeolianites and their fossil components demonstrate that a major climatic change occurred about 17–16 Ma (Fig. 3) resulting in the onset of hyperarid conditions which have persisted ever since [34]. The Aepyornithoid eggshells have been found in Lower Miocene sediments in the Northern Sperrgebiet where the deposits are fluviatile and fluviopaludal. The aeolianites formed in the Middle Miocene as shown by the mammalian sequence. This climatic change is demonstrated by analytical methods such as stable isotopes of C and O applied to the ratite eggshells, a study that led to a better understanding of the climatic changes in the area during the Neogene. The original data used to compile the figures are published in Pickford and Senut [23,25] and in Senut [35].
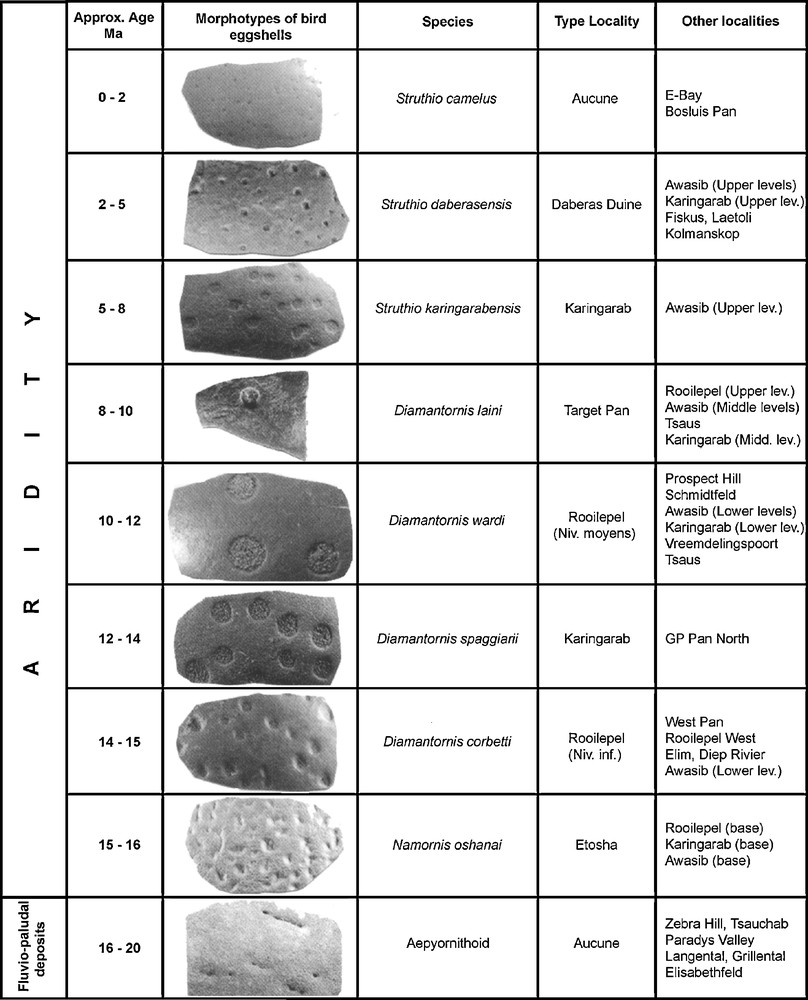
Neogene avian biostratigraphy of the Namib Desert (Namibia) (after [35]).
Fig. 2. Biostratigraphie avienne du désert de Namib (Namibie) pour le Néogène (d’après [35]).
2.1 The stable isotopes of C and O
δ13C and δ18O in ratite eggshells can be used as a surrogate for dietary and climatic conditions [16]. The extant African ostrich is a mixed feeder adapted to subhumid to arid conditions. As an opportunist feeder, its diet consists of C3, C4 and CAM plants. Because they are non-obligate drinkers, modern African ostriches obtain their water requirements from leaf-water. Several studies revealed that eggshell δ18O is linked to bird body water which is in turn dependant on the influence of relative humidity on plants and average δ18Ometeoric water values [13,16]. In arid areas evaporation due to low relative humidity and high soil temperature promotes enrichment in 18O in environmental water. Later, soil water may be enriched in 18O in leaves during evapotranspiration. Plants present different photosynthetic pathways due to discrimination against 13C. Therefore they have distinct isotopic signatures: the mean δ13C of C3 plants is −27‰ (−35‰ to −21‰) and −12‰ (−15‰ to −10‰) for C4 plants whilst CAM plants have intermediate values from −10‰ to −22‰ [4,19]. C3 plants have larger isotopic ranges than C4 plants because the C3 photosynthetic pathways is more sensitive to the [CO2]/[O2] ratio in the atmosphere, light intensity, temperature, and humidity and moisture conditions [10,11]. Low radiation and low temperature during the growing season control the distribution of C3 plants; in contrast high radiation and warm conditions during this period control the distribution of C4 and CAM plants. In the Namib, because of the pattern of wind circulation, two different ecosystems prevail related to two rainfall seasons:
- • South of Lüderitz rain falls during the winter because of the westerlies wind system; the area is dominated by C3 and CAM plants;
- • North of Lüderitz rain falls during the summer because of a tropical rainfall system; C4 plants occur (Fig. 3).
In general, eggshell carbonate is enriched by 15–16‰ in 13C compared to the ingested food [13,43]. Mean δ13C values for modern ostrich eggshell from southern and central Namib regions are distinct: −6.10 ± 1.90‰ and −3.65 ± 2.40‰, respectively and these values are in agreement with the plant distribution in the two areas. For the oxygen, the mean δ18O is 9.55 ± 2.70‰ in the south and 14.20 ± 2.15‰ in the central Namib. These differences are related to the influence of enriched plant water by evapotranspiration on the bird body water and eggshell δ18O values [13,16]. This difference is also linked to different moisture conditions between the two regions. In the aeolianite outcrops, isotopic studies of the fossil ratite eggshells led to the establishment of an isotopic sequence through the last 20 Ma [33]. The main results were that the δ18O values, although variable, are always lower in the southern compared to the central Namib up to 4–6‰ [33] probably related to moisture conditions due to the proximity of the Atlantic Ocean in the south. δ18O trends during the Miocene differ for the two regions: in the southern Namib the mean values are around 6‰; in the central the mean values are between 10 to 16‰. But the trends track each other during the Late Miocene and Pliocene revealing that regional distinction and global climatic change have been recorded by the fossil eggshells. Throughout the Miocene δ13C values for ratite eggshells from both the central and southern Namib regions are indistinguishable showing that the flora comprised C3 (δ13C mean < −9‰) plants. The relative positive mean during the Lower Miocene may suggest a small amount of CAM in the dietary habits of these birds or a relative isotopic high value for the atmospheric CO2 [33]. The mean δ13C values are significantly distinct for these regions as known today. During the Pliocene the mean δ13C values are –9.50‰ in the south and −8.20‰ in the central Namib. Proliferation of C4 plants is observed in the central Namib after ∼5 Ma. This expansion was probably related to energy budgets and rainfall seasonality shifts resulting from large-scale atmospheric and oceanic circulation reorganisation (with the development of cold upwelling cells along the coast) [33].
Eggshells also occur in the Middle Miocene of South Africa and comparisons have been made at a regional scale. The coastal aeolianites at the Prospect Hill site contain eggshells of Diamantornis wardi (10–12 Ma), which have been compared with the Namibian ones. The mean values for δ13C and δ18O of eggshells from Prospect Hill are more negative (about –13‰ for the carbon and of 1‰ for the oxygen) than those from southern and central Namib (Fig. 4). The δ13C isotopic range (between −13‰ to −9‰) is related to the isotopic variability of C3 plants to climatic conditions. The oxygen variation is related to evaporation conditions in the ecosystems. Isotopic data show that during the Upper Miocene the environment at Prospect Hill was clearly more humid (wet) and/or cooler than the Namib with vegetation dominated by C3 and CAM plants. Similar conditions occur today in the Prospect Hill area and similar isotopic values can be found in modern eggshells around this site [13].
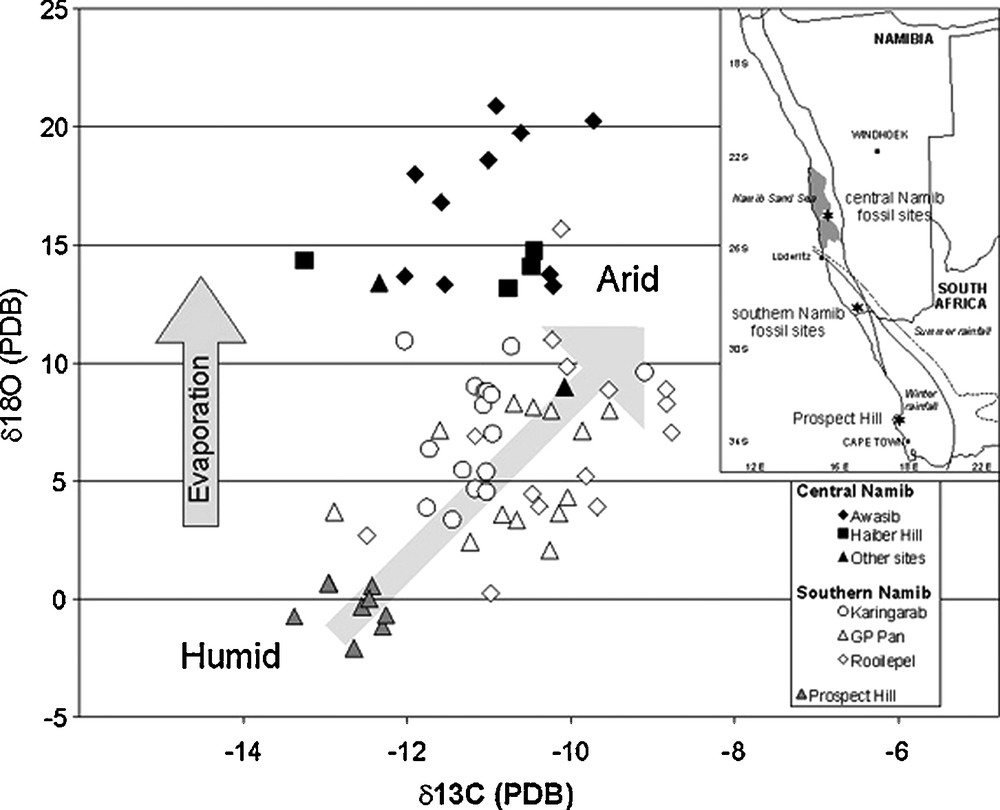
δ13C and δ 18O values of Diamantormis wardi eggshells (10–12 Ma) from Prospect Hill (South Africa) and Namib Desert sites.
Fig. 4. Valeurs des δ13C et δ 18O dans les coquilles de Diamantornis wardi (10–12 Ma) provenant de Prospect Hill (Afrique du Sud) et des sites du Désert de Namib (Namibie).
2.2 Northern Africa
Coeval with Arrisdift in Namibia, the Libyan site of Gebel Zelten (17.5 Ma) has yielded fossil bracket fungus [14] along with tropical forest trees, and a woodland fauna. In the Upper Miocene, the fossils from Sahabi [7] indicate the presence of wooded savannah in northern Libya some 7 Ma.
A suite of micromammals from a vast system of karstic deposits at Gebel Bellorat (11 Ma), includes several forest-adapted lineages such as tree porcupines, galagids and fruit bats [28]. Fossil ratite eggshells have also been recorded in North Africa, which allow us to realise some isotopic comparison at the intracontinental scale. In Egypt, isotopic studies of eggshell remains from the Late Miocene–Early Pliocene sediments at Wadi El-Natrun shows that the mean δ13C and δ18O values are of −9.4 ± 06‰ and 6.3 ± 1‰ respectively (n = 10). These values are quite similar to those obtained for fossil eggshells from the southern Namib [33] and indicate that northern Egypt was dominated by a C3 ecosystem indicating moist conditions with winter rainfall. The data seem to reveal a Mediterranean climate in Egypt at that time, but this needs to be confirmed by further research.
Fossil piscivorous crocodiles from 12–10 Ma deposits in central Tunisia indicate the former presence of large and deep rivers flowing northwards across the country, an indication borne out by the presence of extremely thick (up to 30 m) point bar deposits in the sedimentary successions of Central Tunisia [20].
2.3 Congo Basin
In contrast to Northern African regions, the Congo Basin is today covered in tropical forest which grows on Miocene and Pleistocene aeolian sand deposits up to 250 m thick [12]. These deposits cover much of Congo, northern Angola and Gabon and even reach as far east as Western Uganda where Middle Miocene (13–12 Ma) sand deposits containing large gypsum crystals attest to semi-arid conditions in what is now a very humid part of the Albertine Rift Valley [27,37,39]. Between 10 and 8 million years ago, tectonic activity in the Western Rift led to the creation of a depression and the environment became more and more humid and tropical as indicated by the fossil plants and animals. The fossil fruits and woods were very diverse and similar to those which grow today in North Kasai in the Democratic Republic of Congo [8,39] (Fig. 5).
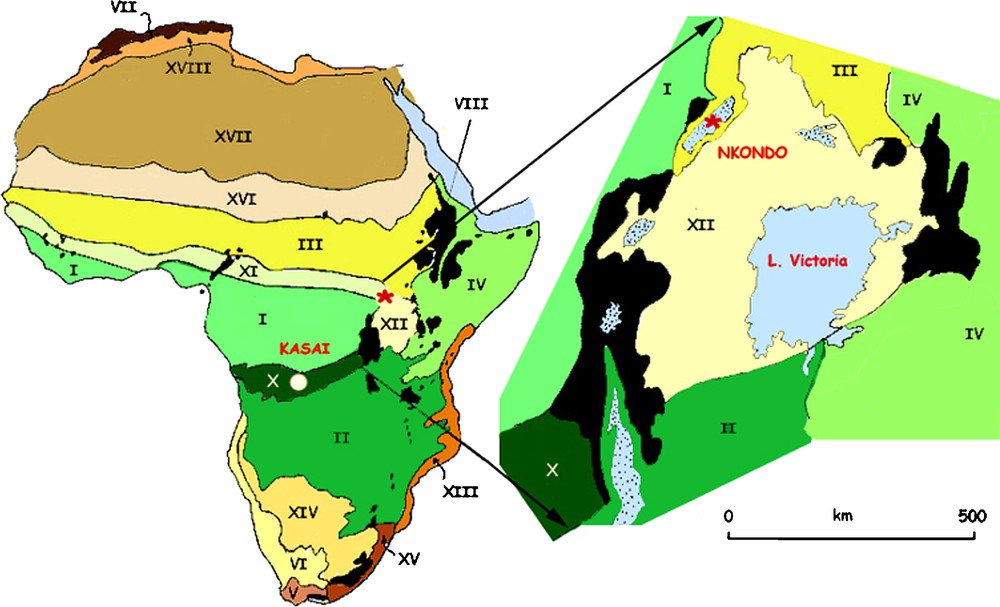
Simplified vegetation map of Africa (after [44]) showing the position of the Nkondo area, Uganda and the Kasai region in Democratic Republic of Congo. Today, Nkondo falls in the vegetation category III (Sudanian) whereas during the Upper Miocene/Lower Pliocene it was in the vegetation category X (regional transition Guineo-Congolian/Zambezian) with the closest analogy being Northern Kasai (DRC). In the Plio-Pleistocene, it fell into the vegetation category III as it is today, suggesting a major change during the Middle to Upper Pliocene.
Fig. 5. Carte de végétation simplifiée de l’Afrique (d’après [44]) montrant la localisation des régions de Nkondo en Ouganda et de Kasai en République Démocratique du Congo. Aujourd’hui, Nkondo tombe dans la catégorie de végétation III (centre régional d’endémisme soudanien), alors qu’au Miocène supérieur/Pliocène inférieur, la végétation y était de type X (zone de transition régionale guinéocongolaise/zambézienne), dont on trouve aujourd’hui l’analogue au Nord Kasai (RDC). Au Plio-Pléistocène, cette région revient dans la catégorie de végétation III, ce qui est toujours le cas aujourd’hui suggérant un changement majeur dans le Pliocène moyen ou supérieur.
Studies of the fossil records of these different areas of Africa reveal a complex history of desertification, with different areas going arid at different times, and some areas going arid at the same time that neighbouring zones were becoming more humid. The geographic position of arid and humid areas appears to have changed throughout the Neogene.
2.4 East Africa
In East Africa, uplift related to tectonic activity during rifting had an important impact on the environments during the Neogene [5,27]. In the Middle Miocene, due to global and local events, the forested environments began to dry out and the faunal composition changed in relation to altitude as shown by the differences in the environments of the Tugen Hills and Lothagam during the Upper Miocene [22,37]. The faunas and floras in the Lukeino Formation are more humid [17,18,24,36] than those of Lothagam [15]. In the Latest Miocene, the vegetation of the Congo Basin contrasted strongly with the more open vegetation in Ethiopia and Kenya which consisted of gallery forests, wooded savanna and savanna [2,6] (and Bamford, pers. comm.). In East Africa expansion of grassland began in the Upper Miocene around 7–8 Ma, but it remained a low component of the environments until the Late Pliocene [3,31]. Then, during the Plio-Pleistocene, desertification took place in low-lying areas, for example in the horn of Africa.
2.5 Toward a synthesis of the desertification of Africa
The earliest evidence for hyperarid conditions documented in the Cainozoic of Africa occurs in the Namib Desert, where thick aeolianites started accumulating ca 17–16 Ma in zones that had been well wooded and subhumid before (Fig. 8) [25]. The Namib has been arid to hyperarid ever since, with fluctuations in aridity indicated by changing faunas and the development of different kinds of palaeosols. The earliest evidence for aridity in the Sahara is much younger than that in the Namib, certainly younger than 10 Ma, and likely to be in the region of 7–6 Ma [23,28–30]. In East Africa, desertification progressed from the Late Pliocene (ca 3 Ma) into the Pleistocene and persists in parts of the region to this day.
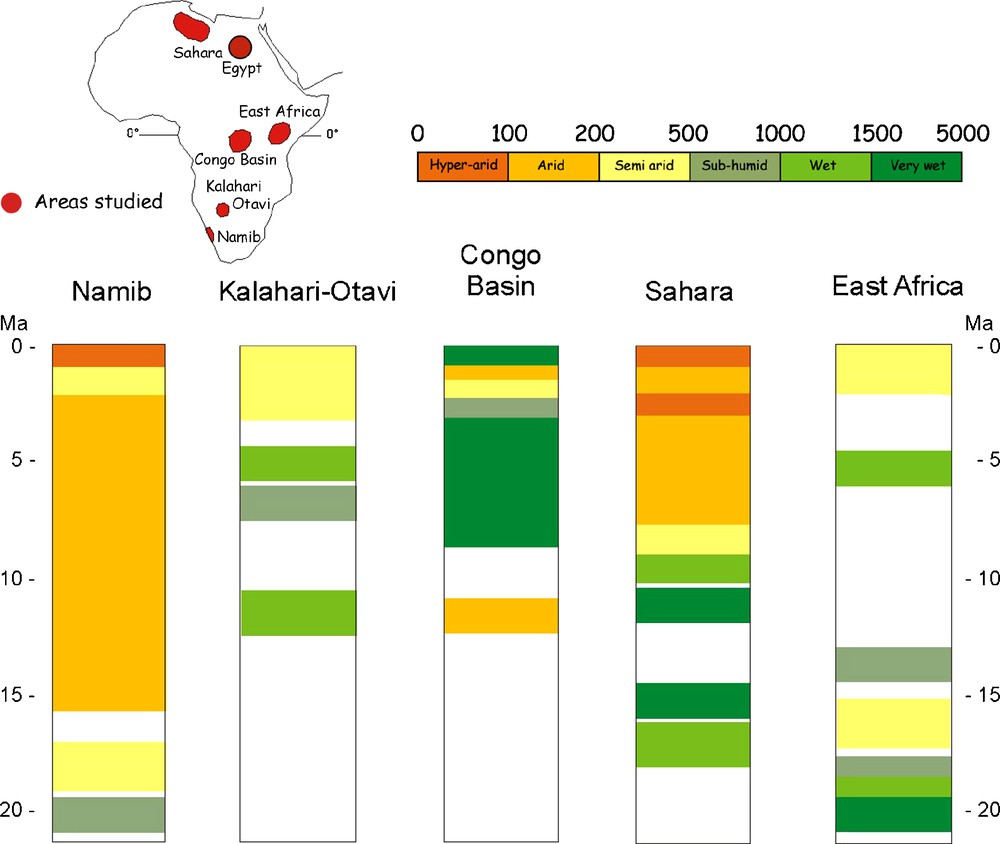
Fossil record showing the complex history of desertification in Africa, with different areas going arid at different times, and some areas going arid at the same time that neighbouring zones were becoming more humid. The geographic position of arid and humid areas appears to have changed throughout the Neogene. Figure modified from [25] in which the original data are available.
Fig. 8. Données fossiles montrant l’histoire complexe de la désertification en Afrique, les différentes régions devenant arides à différentes époques, certaines régions devenant arides, alors que les voisines étaient plus humides. La position des zones arides et humides a changé au cours du Néogène (figure modifiée d’après [25] où les données originales sont publiées).
Interpretation of Cainozoic African fossil localities in terms of the type of phytochore that existed at the time of deposition reconstructed on the basis of the faunas and floras that they have yielded (Fig. 6) (Table 1) shows an overall trend towards aridification in both southern and eastern Africa, with southwestern Africa becoming more arid earlier than East Africa did. This is revealed by the position of the younger localities in the more arid phytochores and the older localities in the more humid phytochores [22].
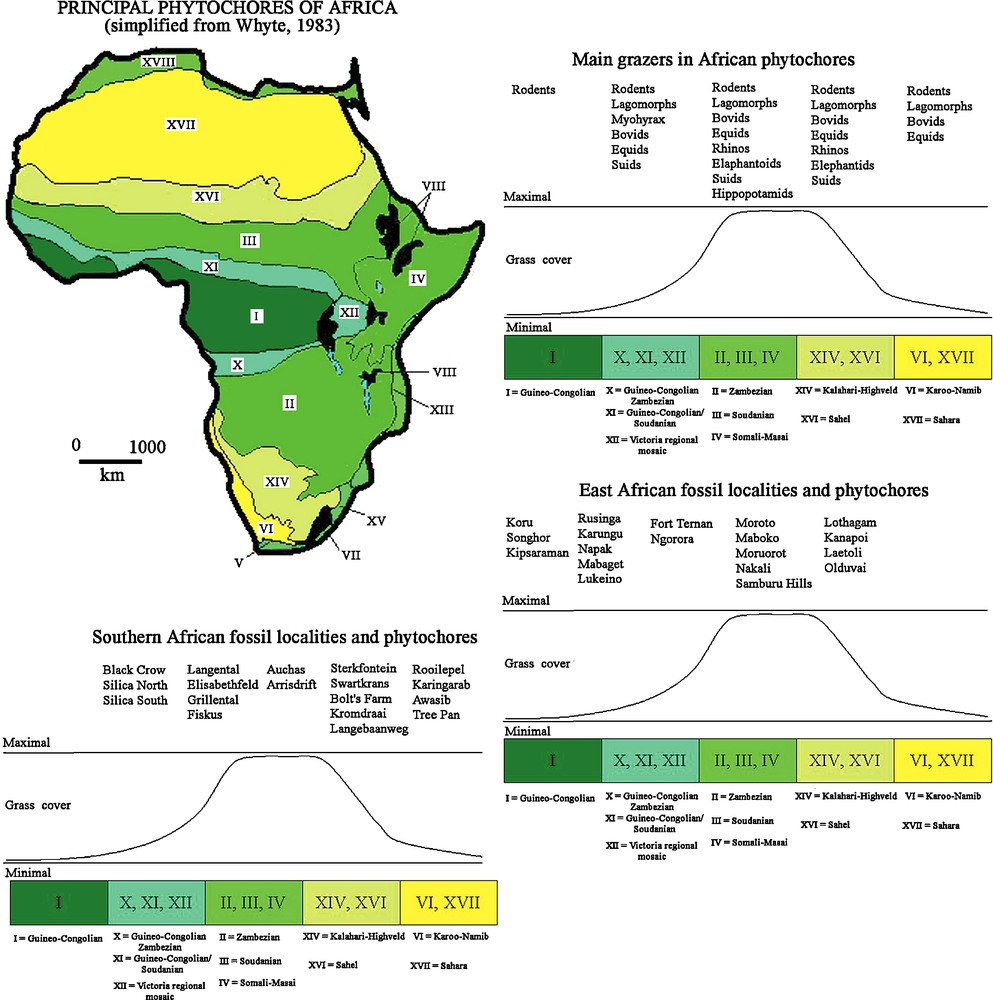
Extant phytochores of Africa and interpretation of Cainozoic fossiliferous localities of Africa. During the Neogene there was an overall trend towards aridification in southern and eastern Africa, with southern Africa developing arid biotopes earlier than East Africa.
Fig. 6. Distribution des phytochores actuels d’Afrique et interprétation des localités fossilifères cénozoïques d’Afrique. Au cours du Néogène, on observe une tendance générale à l’aridification en Afrique australe et orientale, mais les biotopes adaptés à l’aridité se sont développés plus tôt en Afrique australe qu’en Afrique orientale.
List and age of the fossil sites used for the palaeoenvironmental reconstructions in Figure 6.
Tableau 1 Liste et âge des sites fossilifères utilisés pour les reconstitutions environnementales de la Figure 6.
| Eastern Africa | Southern Africa | ||
| Fossil site | Age | Fossil site | Age |
| Koru | Lower Miocene | Black Crow | Palaeogene |
| Songhor | Lower Miocene | Silica North | Palaeogene |
| Kipsaraman | Middle Miocene | Silica South | Palaeogene |
| Rusinga | Lower Miocene | Langental | Lower Miocene |
| Karungu | Lower Miocene | Elisabethfeld | Lower Miocene |
| Napak | Lower Miocene | Grillental | Lower Miocene |
| Mabaget | Pliocene | Fiskus | Lower Miocene |
| Lukeino | Upper Miocene | Auchas | Lower Miocene |
| Fort Ternan | Middle Miocene | Arrisdrift | Middle Miocene |
| Ngorora | Middle and Upper Miocene | Sterkfontein | Plio-Pleistocene |
| Moroto | Middle Miocene | Swartkrans | Plio-Pleistocene |
| Maboko | Middle Miocene | Bolt's Farm | Plio-Pleistocene |
| Moruorot | Middle Miocene | Kromdraai | Plio-Pleistocene |
| Nakali | Upper Miocene | Langebaanweg | Pliocene |
| Samburu Hills | Middle and Upper Miocene | Rooilepel | Middle Miocene to Holocene |
| Lothagam | Mio-Pliocene | Karingarab | Middle Miocene to Holocene |
| Kanapoi | Pliocene | Awasib | Middle Miocene to Holocene |
| Laetoli | Pliocene | Tree Pan | Middle Miocene |
| Olduvai | Plio-Pleistocene |
A major consequence of the early development of hyperarid environments in southern Africa, at the end of the Early Miocene, as far away from Eurasia as it is possible to go, is that plants and animals in the region had a considerable period of time to adapt to these conditions with minimal interference from plants and animals from Eurasia that may have been adapting to similar environments. The result was the evolution of a whole suite of autochthonous plant and animal lineages adapted to arid, climatically unstable environments. Lineages involved include crocodiles, ostriches, various rodent families (Pedetidae, Bathyergidae), macroscelidids (elephant shrews or senges), prohyracid hyracoids, tubulidentates, some carnivores and bovids among others. Plants were also involved in this evolutionary activity, but the details are less well-known due to a paucity of palaeontological studies, but grasses came to dominate ecosystems in Namibia much earlier than they did in East Africa, and in several mammal lineages this led to the evolution of a wide range of dental strategies (hypsodonty, ptychodonty, cementodonty, pliocodonty, polylophy) to extend the life of cheek teeth that were experiencing excessive wear due to the presence of opal phytoliths in the grass that they were consuming.
Adaptation of animals and plants to arid environments continued unhindered in Namibia for up to 10 million years before the first signs of aridity became evident in the Sahara and some 14 million years before East Africa started going dry. An interesting point is that when the environment in northern and eastern Africa began to open up due to increasing aridity, local plants and animals had no time to adapt before they were displaced by arid-adapted lineages spreading from South Africa on the one hand, and from arid zones in Eurasia on the other. This is demonstrated in Fig. 7 which shows the changes in the ratio between brachyodont to hypsodont mammalian species in Africa during the Neogene. Each fossil site has been allocated a colour (dark in brachyodonts, light in hypsodonts) on the basis of the presence or absence of hypsodont and/or brachyodont species. For example, in the Lower Miocene, hypsodont and brachyodont species are equal in number in southern Africa, but in East Africa, brachyodont species clearly dominate the fauna. In the lower Middle Miocene, the hypsodont species are still better represented in South than in East Africa. The East African fossil record thus shows an interesting history of faunal changes implicating lineages from southern Africa (pedetids, bathyergids, some suids, some bovids, aardvarks, ostriches, and crocodiles, for example) and from Eurasia (porcupines, leporids, hyaenas, giraffes, some suids, some bovids, equids, and camels, among others). Many East African animal lineages either went extinct (the crocodiles Euthecodon and Rimasuchus for instance) or retreated west as the forest cover shrank (osteolamine and cataphractine crocodiles, and tragulids for example). In contrast, fossil faunas from North Africa, such as those at Sahabi, Libya, tend to show much more evidence of faunal interchanges with Eurasia than do East African and South African faunas, although more fossil evidence is required from the Late Miocene of the latter regions to throw light on the details of the exchanges.
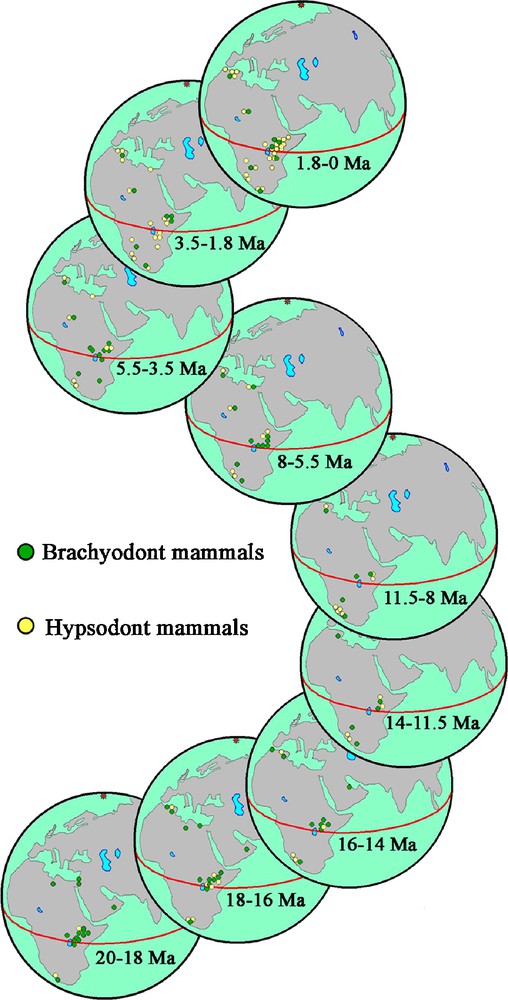
Changes in the ratio between brachyodont to hypsodont mammalian species in Africa during the Neogene. Functionally, hypsodonty is associated with grazing, and the history depicted here shows the increase in the preponderance of open biotopes with grassland through the Miocene and Plio-Pleistocene.
Fig. 7. Changements dans les rapports entre espèces mammaliennes brachyodontes et hypsodontes en Afrique au cours du Néogène, estimé sur chaque localité fossilifère. Fonctionnellement, l’hypsodontie est associée à l’ingestion d’herbes et l’histoire retracée ici montre l’expansion des biotopes ouverts avec des prairies au cours du Miocène et du Plio-Pléistocène.
As a consequence of the complex history of desertification in Africa, it is essential to study the fossil record from all parts of the continent in order to obtain a holistic view of how desertification progressed, when it occurred, and what effects it had on the development of faunas and floras in the different regions of the continent. Only then can it be appreciated that the Namib Desert, even though it is tiny compared to the Sahara, had a considerable impact on the development of African plant and animal communities, simply because it has been arid for so much longer (since the Middle Miocene) and was so far removed from other arid areas in Europe and Asia. The animals and plants that live in the Sahara, in contrast, show a great deal of influence from Eurasia [1]; indeed much of the northern Sahara (Maghreb, Libya, Egypt) is populated by plants and animals (deer, bears, various land snails) with Palaearctic affinities, although it is clear from the fossil record that there were periods when these same areas had a substantial presence of animals that had spread northwards from equatorial and southern Africa (gelada baboons, aardvarks, wildebeest, phacochoeres).
When the ratio between brachyodont and hypsodont mammals is considered for the Neogene of Africa (Fig. 7), it becomes obvious that brachyodont animals dominated the faunas in the Early Miocene and hypsodont animals during the Pleistocene. However, the changes did not occur simultaneously throughout the continent. Hypsodont taxa started to dominate the faunas in Namibia long before they became dominant in eastern and northern Africa.
3 Conclusion
Aridification of parts of Africa during the Cainozoic can be reconstructed by studying fossil sites (palaeontology, sedimentology, isotope geochemistry). Studies carried out over the past two decades reveal that different parts of the continent became arid at different times, and that some areas became more humid at the same time that other areas were going more arid (Figs. 3, 6, 8). Correlation of the local and regional palaeoclimatic changes at various localities reveal that the history of desertification in Africa (indeed, throughout the world) was intimately linked to global climatic changes, in particular to the history of polar ice caps (Fig. 3). The Antarctic Ice Sheet started to develop during the Oligocene, but it did not expand to cover the entire continent until the end of the Early Miocene. It was this expansion that produced the conditions in the southern mid-latitudes for desertification to proceed on land, with the consequence that the Namib in Africa and the Atacama in South America became arid to hyperarid some 17–16 Ma. At this time there was no counterbalancing Arctic Ice Cap, a consequence of which was the northward displacement of the world's ecoclimatic belts (temperate belt, subtropics, and tropics). However, between 8 and 7 Ma, the Arctic Ice Cap grew to immense proportions, first on Greenland and then onto the Arctic Ocean, and as a result the world's ecoclimatic belts were shifted back towards the south. Mid-latitude Europe, which had enjoyed a tropical to subtropical climate for most of the Middle and Late Miocene (16–8 Ma) [1] became temperate and boreal. The septentrional arid zones which used to be located in midlatitude Eurasia, also shifted southwards, and it is from this time that the Sahara, Arabian and Thar Deserts became arid.
An understanding of the processes and timing of desertification in Africa is thus greatly dependent on a proper understanding of the African fossil record, not just from one area, but from all latitudes of the continent.
Acknowledgements
We thank the organizers of the Colloquium of the Academy of Sciences “African and Arabic Deserts: Environment, climate and impact on populations” for allowing us to present our data on the Neogene desertification of Africa. We are grateful to Prof. Y. Coppens for his longstanding support and all the colleagues who have been involved in the different projects around Africa.
This work was funded by several institutions: the CNRS (GDR, GDRI, UMR 5147), the Collège de France (Chaire de Paléoanthropologie et de Préhistoire, Prof. Y. Coppens; Chaire de Biologie historique et Évolutionnisme, Prof. A. de Ricqlès), Université Pierre-et-Marie-Curie (BQR), the Ministère des Affaires Étrangères et Européennes (Commission des fouilles), the SCAC of the French Embassy in Windhoek, the French Embassy in Uganda, the Uganda Museum in Kampala, the Geological Survey of Namibia, Namdeb Diamond Corporation (Pty) Limited, de Beers, the National Research Foundation of South Africa, the Palaeoanthropology Scientific Trust, the University of Cape Town, and Menoufia University at Shebin-el-Kom. The comments of the anonymous reviewers have been useful for improving the paper.