

1 Introduction
Since the Oligocene to the Late Miocene, the Mediterranean experienced several zooxanthellate coral building episodes (for an overview, see Perrin and Bosellini, 2012). Many authors mentioned a strong decrease in the biodiversity of corals during this time interval, with the lowest generic diversity during the Messinian (e.g. Chevalier, 1961; Esteban, 1996; Perrin and Bosellini, 2013). Such a decrease was generally related to the progressive closure of seawater exchanges between the Indian Ocean and the Mediterranean through the Mesopotamian Basin during the Miocene (see Popov et al., 2004), and also to climate and/or seawater circulation and temperature changes (e.g. Budd et al., 1996; Chaix et Saint-Martin, 2008; Chevalier, 1961; Esteban, 1996). Among the Cainozoic carbonate systems of the Mediterranean, the Upper Burdigalian coral reefs remain poorly known (Esteban, 1996; Perrin and Bosellini, 2012), but well exposed in southern Corsica. The aim of this work is to provide information on their architecture and generic diversity in order to better constrain the z-coral building story through the Miocene in the Mediterranean.
2 Geological setting
The coral build-ups of southern Corsica occur in the Bonifacio area (Fig. 1; Arnaud et al., 1992; Monleau et al., 1996; Orszag-Sperber, 1978; Orszag-Sperber and Pilot, 1976), in the Cala di Labra Formation (Ferrandini et al., 2002, 2003), which dates from the Upper Burdigalian (ca. 19 Ma at oldest and 15.97 Ma at the top, calibration issued from the biostratigraphic works by Ferrandini et al., 2002, 2003, 2010). Only localized information is available from these build-ups (Chevalier, 1961; Galloni et al., 2001; Tomassetti and Brandano, 2013).
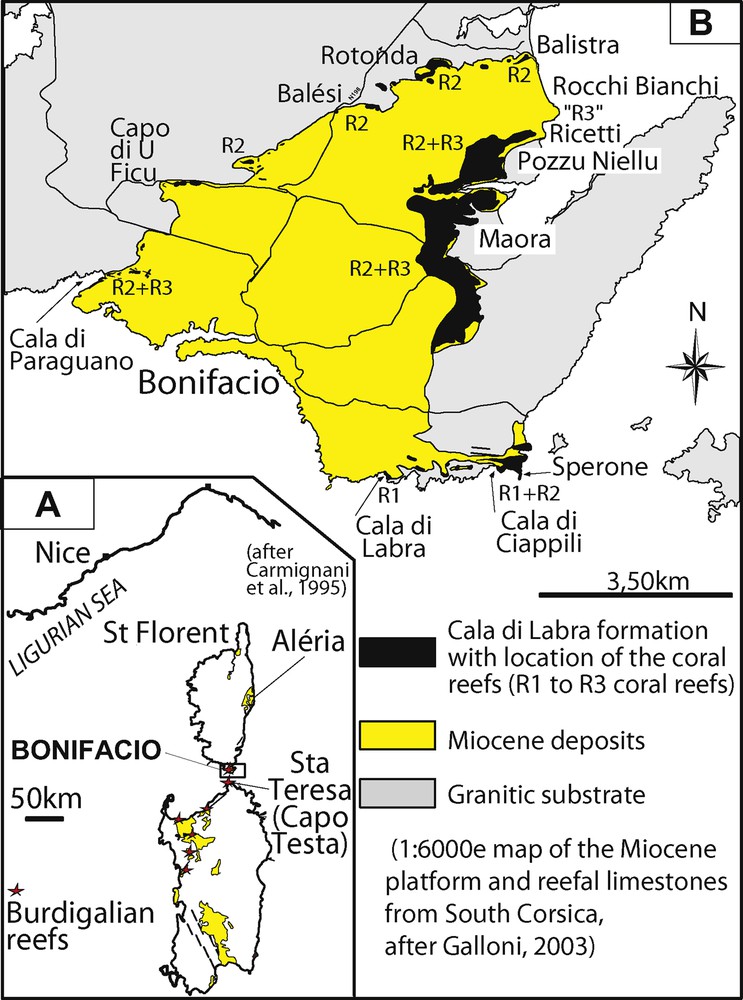
(Color online) (A) Geological setting of the Corsica-Sardinia microblock; (B) Map of the Bonifacio Miocene outcrops and location of the studied areas.
The Cala di Labra Formation was deposited at the end of the fast, anticlockwise rotation of the Corsica–Sardinia microblock, responsible for the opening of the Ligurian-Provençal micro-ocean in the western Mediterranean (e.g., Gattacecca, 2001; Gattacecca et al., 2007; Oudet et al., 2010). This formation is 50 metres thick. It comprises, from bottom to top, five superimposed lithofacies (Fig. 2; Ferrandini et al., 2002, 2003; Galloni, 2003; Tomassetti and Brandano, 2013; Tomassetti et al., 2013), which have been mapped and correlated over the whole area (Ferrandini et al., 2003; Galloni, 2003; Orsini et al., 2010): a coral reef unit (F1 = U1 of Galloni, 2003) resting on boulders of a Variscan granite, a conglomeratic then shelly litharenitic unit (F2 = U2 + U3A), a sandy to silty unit (F3 = U3B + U4), and two bioclastic units (F4 = U5; F5 = U6A). The siliciclastic facies F2 was deposited in a shoreface environment and the siliciclastic facies F3 in a distal shoreface to upper offshore environment (Ferrandini et al., 2002, 2003; Galloni, 2003; Reynaud et al., 2012). This formation is overlain by the mixed carbonate-siliciclastic, wave-to-tide-dominated Early Langhian Bonifacio Formation (Ferrandini et al., 2003, 2010; Reynaud et al., 2012).

(Colour online) Stratigraphical setting of the Cala di Labra Formation and studied sedimentary units (U1 to U5).
From mapping and biochronological investigations, Ferrandini et al. (2002, 2003), Galloni et al. (2001), Galloni (2003) and Orsini et al. (2010) demonstrated that three superimposed, reefal building episodes were recorded in the Cala di Labra formation, in the vicinity of granitic palaeohighs: R1, at the bottom (lithofacies F1), R2, coeval with lithofacies F3 (divided into sub-episodes R2A and R2B) and R3, coeval with lithofacies F4 (Fig. 2). All these reefal episodes were submitted to siliciclastic inputs supplied from the erosion of the neighboring Variscan granites or from underlying Aquitanian arkosic deposits (Balistra Formation; Fig. 2).
3 Methods
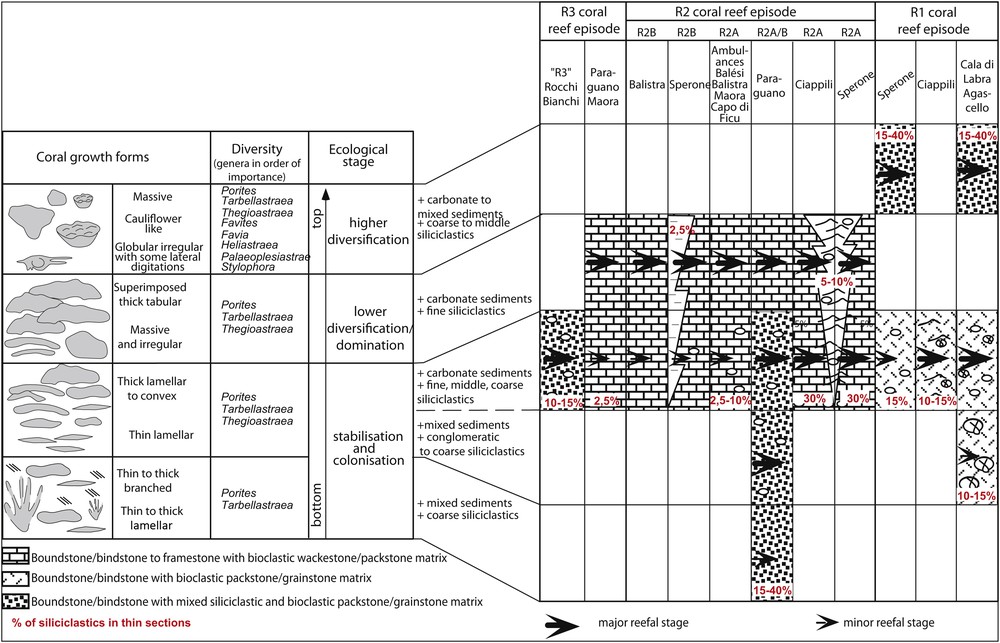
The coral reefs and adjacent rocks were mapped at the 1/6000 scale (Galloni, 2003). Coral reefs were also mapped on photographs from cliffs. The better-exposed outcrops were also investigated at the scale of the coral colonies to illustrate the coral growth patterns and estimate the relative contents of the different genera. Detailed cross-sections were conducted on each build-up. A hundred thin sections were studied for palaeoenvironmental reconstructions. The petrographic examination included semi quantitative visual estimation of the organic and inorganic components in thin sections. The reef anatomies, geometries and internal architectures were identified for each coral reef episode by using coral species, growth forms, size of colonies, matrix, associated organisms and siliciclastic content. A total of 16 coral species and 8 coral growth forms were listed for the first time.
4 Results
4.1 R1 coral reef episode
The R1 coral reefs are only present in the Cala di Labra–Ciappili–Sperone area, where they onlap the granitic basement (Fig. 1). They consist in three- to five-metres-thick bodies, lying upon a granitic conglomerate with decimetric to metric elements surrounded by carbonate packstone. The packstone/grainstone matrix comprises corals (Porites, Thegioastraea and Tarbellastraea), red algae, bryozoans, bivalves (oysters, pectens), larger Foraminifers (miogypsinids, amphisteginids) and 10 to 15% of coarse-grained siliciclastics. The reefs contain the highest diversity of coral assemblage of the Bonifacio area (Fig. 3), with 16 species belonging to the genera Porites, Thegioastraea, Tarbellastraea, Favia, Favites, Palaeoplesiastraea, and Stylophora. At Sperone (Fig. 1), the assemblage is completed with Heliastraea s.l. sp.1, Heliastraea sp. 2, Favia valceppii, and Favites detecta. From base to top, the reefs reveal a vertical zonation: in their lower part, they display lamellar and binding colonies with few genera; in their uppermost part, they display highly diversified lamellar, massive and cauliflower-like colonies (Fig. 4) bored by lithophagid organisms. At Cala di Ciappili, several coral thickets alternate with wavy bedding bioclastic packstone/grainstone limestones. The R1 reefs are overlain by the coarse-grained sandstones of the F2 unit, which were deposited in shoreface environments, without erosion at the top of R1.

Main coral species in the R1 and R2 coral episodes (systematics, C. Chaix).
(Color online) Relationships between coral growth forms, vertical zonation, hydrodynamism and siliciclastics in the reefs of Bonifacio.
4.2 R2 Coral reef episode
The R2 coral reefs are often comprised of two subunits, R2A and R2B, reaching ten metres in thickness. R2A and R2B are generally separated by a sharp surface or by bioclastic beds.
R2A reefs are massive and stratified. They are well developed in the Pointe Sperone and Maora areas, and localized outcrops are found along the western edge of the Bonifacio basin (Fig. 1B). R2A reefs are present only in the vicinity of granitic highs (Sperone, Paraguano, Ricetti, Balistra, Capo di U Ficu). Between Ciappili and Sperone, littoral channels infilled with bioclastic grainstones with current ripples and thickets of lamellar coral colonies are interbedded in the coral complex. Except at Cala di Paraguano, R2A reefs display thin to thick lamellar colonies dominated by poritids and Tarbellastraea (Fig. 4). The coral assemblage comprises Tarbellastraea reussiana, Thegioastraea haimei, Th. burdigalensis, Porites arenosa, and Porites sp. (Fig. 3). The matrix of the reefs is comprised of packstones/grainstones/floatstones, with rounded bioclasts of red algae and benthic foraminifers (amphisteginids, miogypsinids, operculinids, rare miliolids, Sphaerogysina globula, Rupertina and rare Martinotiella communis). Coarse siliciclastic grains are present and can represent up to 5–10% of the matrix. At Cala di Paraguano, lamellar colonies of Porites and Tarbellastraea are locally associated with columnar and branching colonies (length 20–50 cm, 2 cm in diameter) upward changing into thick tabular colonies. The packstone/grainstone matrix is dominated by red algae, rotaliids, bivalves, echinoids and gastropods, and siliciclastic grains reach 15 to 20% in abundance (Fig. 4).
R2B reefs are stacked above R2A reefs. Porites, Tarbellastraea and Thegioastraea are the main framebuilders (Fig. 3). At Sperone, the coral growth forms are dominantly thin to thick lamellar, associated with red algae, bryozoans and bivalves (Fig. 4). The matrix is a foraminifer-rich grainstone (20%: amphisteginids, operculinids, rotaliids, arenaceous) with red algae debris (15%). The reefs grade towards the northeast (in the direction of granitic substrate) into muddy and red algae/benthic foraminifer-enriched packstones to floatstones. At Balistra, the R2B reefs are made of peri-reefal bioclastic limestones with sparse coral colonies, together with some Miogypsinidae, Operculina gr. complanata, Amphisteginidae, bivalves and echinoids. From north to south, this facies laterally changes into operculinids and red algae-enriched argillaceous limestones, then into bryozoan enriched siltstones and finally into sandy/silty marls with planktonic foraminifers of Unit 4 (Fig. 2).
4.3 R3 coral reef episode
R3 reefs are present in the Rocchi Bianchi-Maora area and at Cala di Paraguano (Fig. 1B). They are massive and bedded build-ups resting on both the granitic basement and the R2B reefs. They are widely dominated by Thegioastraea and Porites genera organized into lamellar to massive colonies (Fig. 4). The matrix displays frequent bryozoans, larger Foraminifers (Miogypsinids and Amphisteginids), encrusting foraminifers, miliolids, some planktonic foraminifers, bivalves, echinoids, and ostracods. R3 reefs laterally change into bedded red algae boundstone (Maora) or into red algae-rich wackstone to floatstone with a micritic, pelloidal matrix. At Rocchi Bianchi, only R3 reef-derived debris of tabular Poritids are found in a conglomerate dominated by granite-derived elements.
5 Discussion
5.1 Depositional models
All of the Upper Burdigalian coral reefs of the Bonifacio area onlap the granitic basement, indicating that they formed during a general relative sea-level rise culminating during lithofacies F5 deposition (U6A, Fig. 2; Ferrandini et al., 2002, 2003; Galloni, 2003; Reynaud et al., 2012). They are parts of an Upper Burdigalian transgressive system tract (Reynaud et al., 2012). The reefs of the Bonifacio area are dominantly organized into biostromal build-ups emplaced under high-to-medium energy conditions, under fair-weather wave action (domination of binding, lamellar and thicket coral forms, abundance of broken bioclasts, frequent siliciclastic inputs). Current activity is clearly recorded, for example, during deposition of reefs R2A at Sperone, where grainstone deposits are preserved in ENE–WSW-trending littoral channels. The associated larger Foraminifers, amphisteginids, miogypsinids, operculinids, and the absence of heterosteginids are also indicative of shallow-water reefs (BouDagher-Fadel, 2008). As a whole, the corals reefs of the Bonifacio area are reefs fringing a rocky, irregular coastline submitted to wave action (Fig. 5). In some rare cases, protected areas can be found, as for example at Cala di Paraguano. There, branching colonies developed during R2A deposition and branching red algae in muddy sediments during R3 episode. Our results contradict the deep building setting (limit of the upper photic zone) of the R1 reefs at Cala di Labra proposed by Tomassetti and Brandano (2013). Moreover, R1 reefs display a clear vertical zonation and were drowned by shoreface deposits of unit 2, indicating that the R1 reefs are only part of a transgressive–regressive cycle, not a single cycle (Tomassetti and Brandano, 2013).

(Color online) Depositional models for the R1, R2 and R3 upper Burdigalian reefs at Bonifacio.
5.2 Biodiversity and vertical zonation of the reefs
The R1 coral reefs display the highest coral diversity in the Bonifacio area, with eight genera (Porites, Thegioastraea, Tarbellastraea, Heliastraea, Favia, Favites, Palaeoplesiastraea, Stylophora) related to 16 species (Fig. 3). R2 and R3 coral reefs are more extended than R1 ones, but they display only three genera (Porites, Tarbellastraea and Thegioastraea) related to a total of six species.
The vertical organization also displays major differences from bottom to top. The R1 reefs show a clear vertical zonation (Fig. 5) with at the base few species and at the top a maximum diversification in the coral assemblage. This indicates that the R1 reefs reached a climax stage. In contrast, R2 and R3 reefs do not display significant vertical changes concerning both internal architecture and biodiversity, excepted for the R2 at Cala di Paraguano. R2 reefs did not reach a pronounced diversification stage. The genera Heliastraea, Favia, Favites are no more present. Such an impoverishment in the coral assemblage from R1 to R2–R3, the dominant ubiquitous coral genus as Porites, Tarbellastraea and Thegioastraea in R2–R3 coeval with the development of red algae are significative of a change in seawater parameters (§ 5.3).
5.3 Upper Burdigalian reefs in the Corsica–Sardinia microblock
The general impoverishment in coral diversity during Cainozoic times in the Mediterranean, culminating with Porites monogeneric reefs during the Late Miocene, was explained by the interplaying effects of a global cooling, the re-organization of the Tethyan realm and the emergence of the Alpine chains, driving the gradual northward movement of the whole region outside the tropical/subtropical belt (Perrin and Bosellini, 2012). During the Burdigalian, Corsica and Sardinia were fully located in the reefal tropical belt (Fig. 6; Perrin and Bosellini, 2012; Popov et al., 2004). The coral diversity often reached 20–25 genera (Harzhauser et al., 2007; Perrin and Bosellini, 2012) and a total of respectively 75 and 40 genera were recorded in the subreefal facies of the Atlantic or the Alps (Fig. 6; Budd et al., 1996; Cahuzac and Chaix, 1996; Chevalier, 1961).
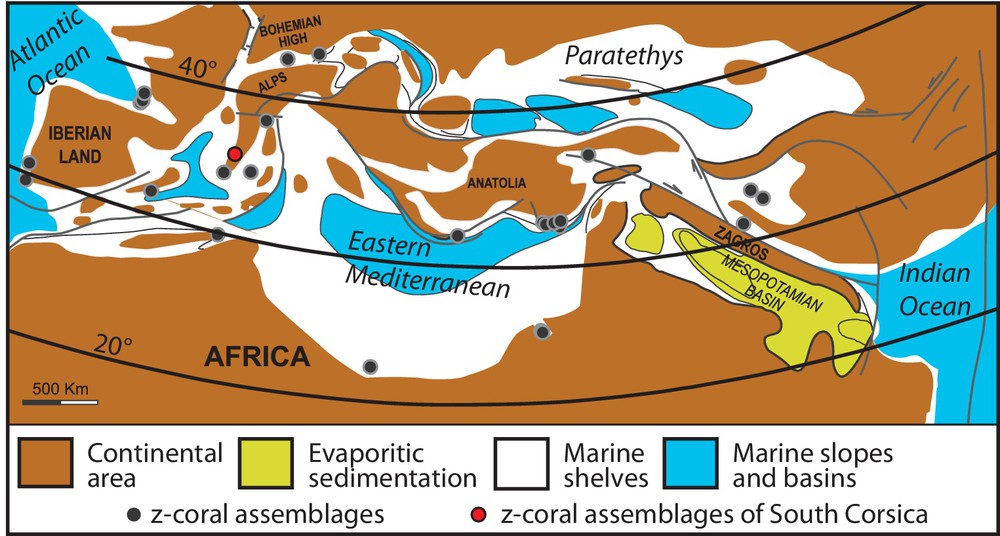
(Color online) Distribution of coral reefs during Burdigalian in the Mediterranean area (after Popov et al., 2004 and Perrin and Bosellini, 2012, modified).
Upper Burdigalian coral reefs are present in northern Sardinia (Arnaud et al., 1992; Assorgia et al., 1988; Brandano et al., 2010; Martini et al., 1992; Mazzei and Oggiano, 1990). At Capo Testa (Fig. 1A), they display rather similar biodiversity and organization than in southern Corsica. They are considered as patch reefs that developed in the 35–45m depth interval dominated by the coral genus Porites (Brandano et al., 2010). Ferrandini et al. (2010) also noticed that the coral diversity decreased upward in the reefal succession, suggesting similarities with R2 and R3 reefs of southern Corsica. By contrast, coral build-ups are poorly known in the Upper Burdigalian deposits of northern, central and eastern Corsica (Fig. 1A; Orszag-Sperber, 1978). Only some coral bioclasts were found in the sediments, suggesting that coral activity extended to Alpine Corsica in the North (Ferrandini et al., 1998, 2003; Löye-Pilot et al., 2004).
The Bonifacio area displayed a favourable area for the installation of coral build-ups, with a shallow-water, granitic shoal between Corsica and Sardinia (Bonifacio Straight; Reynaud et al., 2012). Nevertheless, the presence of 8 then 3 coral genera in South Corsica indicates a low coral diversity regarding other Mediterranean areas, as in North Sardinia (Brandano et al., 2010), reflecting local vanishing conditions through upper Burdigalian times in these areas. Less favourable conditions are also recorded by the changes in internal organization and biodiversity observed between R1 and R2–R3 coral reefs.
The main factors controlling coastal reef growth and diversity are changes in nutrient levels, temperature, salinity, seawater circulations, atmospheric CO2 concentration and Ca2+oceanic concentration (for an overview, see Montaggioni and Braithwaite, 2009). In the southern Corsica–northern Sardinia area, such vanishing conditions can be related to:
- • constant siliciclastic inputs (Fig. 5) issued from the emergent, neighbouring Variscan basement during the reefs building episodes; such inputs probably increased the nutrients level in coastal, protected areas;
- • during the Upper Burdigalian–Early Langhian transition (Units 5 to 6, Fig. 2) the middle part of the Upper Burdigalian Bonifacio straight subsided and water depth increased (Reynaud et al., 2012); such a depth increase was responsible for major changes in current activity: the Upper Burdigalian, a wave-dominated shoal, changed into a high-energy, tide-dominated, bioclastic shoal between the Tyrrhenian shelf and the western Mediterranean (Reynaud et al., 2012; Fig. 6).
6 Conclusion
During the Upper Burdigalian, coral reef build-ups were emplaced in southern Corsica. Three major superimposed, shallow-water fringing reef episodes are recorded in the Cala di Labra Formation (Bonifacio area). The first coral reef, R1, developed against a rocky granitic coast. It reached a diversification stage with 16 coral species. The following R2 and R3 episodes are wide-extent build-ups. They display a decrease in coral diversity, with six coral species, among which Porites, Thegioastraea and Tarbellastraea dominate. These reefs are often related to coeval red algae-rich deposits. By comparison to other contemporaneous coral build-ups of the Mediterranean, the relative lesser biodiversity than that in the southern Corsica–northern Sardinia area is interpreted as resulting from localized less favourable seawater conditions, because of both siliciclastic inputs and deepening of the Bonifacio straight at the Burdigalian–Langhian transition.
Acknowledgments
The PHD of FG study was sponsored by Cécile Pabian Goyheneche and Michel Rebelle (ELF EP) then by TOTAL throughout the ARTEP Research Project (“Quantification des réservoirs carbonatés”) in collaboration with IFP and GdF(ELF EP 11833 and 13083). This work is dedicated to the memory of Jean-François Babinot. Michelle and Jean Ferrandini are strongly thanked for help and scientific discussions. J.-Y. Reynaud, F. Fournier and S. Bourquin provided constructive reviews.