

1 Introduction
The sabkha of Gueran is an internally salty drained depression in the Sahara located 125 km southeast of the Atlantic coastal city of Boujdour, in southern Morocco (Fig. 1). The site was discovered in 2014, and dozens of skulls and partially articulated skeletons of late middle Eocene archaeocetes have been collected by local prospectors. During four successive field missions realized between 2014 and 2016, with the financial support of the National Geographic Society, hundreds of isolated remains of marine vertebrates have been collected by Zouhri, Gingerich, and Khalloufi in a unique fossiliferous bed that extends over 20 km.
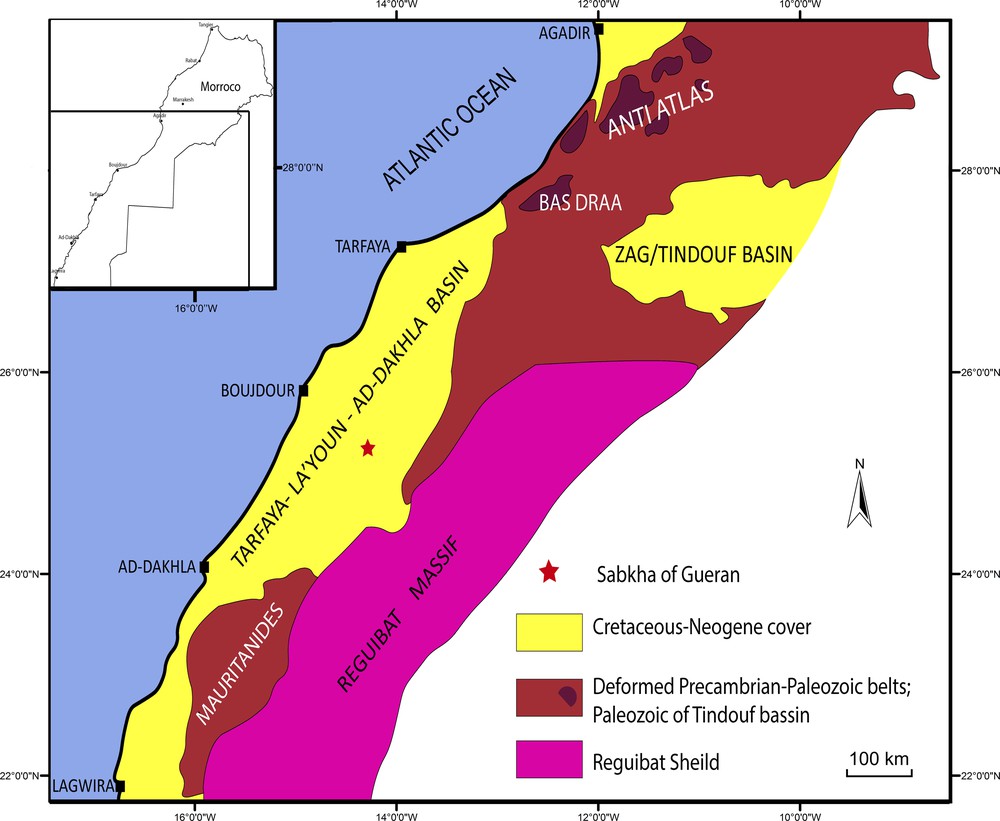
Simplified structural map of southern Morocco showing the location of the sabkha of Gueran.
Modified from Hollard et al., 1985.
The interest of the Gueran locality lays in the rarity of late middle Eocene faunas in northwestern Africa. The late Eocene fossil record documents the impact of global climate changes at the Eocene–Oligocene transition on the evolutionary and biogeographical history of biodiversity. Bartonian archaeocetes previously described by Gingerich and Zouhri (2015) are among the most diverse archaeocete faunas of this age in the world. The extraordinary richness and excellent state of preservation of Bartonian archaeocetes document a crucial period in the evolution of cetaceans. Gueran archaeocetes include both protocetids and basilosaurids and are dated to the Bartonian because basilosaurids are not known from the Lutetian, and a single protocetid is known to have extended into the earliest Priabonian. The discovery of new archaeocete specimens makes it possible to strengthen the previous study. Other vertebrate taxa are also well represented at Gueran and contribute to a better understanding of the evolutionary history of taxa still poorly documented in the Eocene, like proboscideans and pelagornithid seabirds.
This paper aims to provide a more complete picture of the fossil assemblage of Gueran and to refine the dating of this fauna by comparison with contemporaneous faunas of the Fayum Depression in Egypt, Dur At-Talah in Libya, and Eocene localities on the Atlantic coast of western Africa (Nigeria, Togo, Senegal, and Morocco). Additionally, we discuss the biostratigraphical, palaeobiogeographical, and paleoenvironmental significances of this diverse marine fauna from the upper middle Eocene strata of northwestern Africa.
2 Geological setting
The marine fauna described herein comes from the Aridal Formation in the Gueran Depression (Fig. 1). Geologically, this formation belongs to the sub-basin of Boujdour, which constitutes, along with the sub-basin of Dakhla, the Atlantic basin of Tarfaya–La’Youn–Ad-Dakhla (Davison and Dailly, 2010; Ranke et al., 1982). This basin is the onshore, proximal part of the Atlantic passive margin; the oldest, Triassic–Liassic deposits of the margin are lacking here beneath the continental Lower Cretaceous sandstones and overlying Upper Cretaceous–Cenozoic marine beds (Hafid et al., 2008).
Ratschiller (1967, 1970) was the first to map the geology and study the stratigraphy of the northern region of the current Moroccan Sahara. He placed the strata yielding the fossils described in this work in the Gueran Member of the Samlat Formation. The Gueran Member was named for strata exposed at Gueran (Ratschiller, 1967: 108; 1970: 25, 75; type section “eastern flank of Gueran”), and the Samlat Formation as a whole was named for strata exposed at Samlat Amgrach farther to the west. Ratschiller gave the thickness of the Gueran Member as 45 m, and divided it into (1) a lower 21 m of chalk with intercalated siltstones and sandstones, shark teeth, etc.; (2) an upper 22 m of white chalk with unclear bedding; overlain by (3) a 2-m limy crust.
Lindner and Querol (1971) published a more detailed geological map of the former Spanish Sahara, in which they mapped the Aridal Formation. This was named for exposures at Sabkha of Aridal, east of Boujdour. At Gueran, the Aridal Formation of Lindner and Querol (1971) is equivalent to the Gueran Member of the Samlat Formation of Ratschiller (1967, 1970). Aridal Formation is the name used on the current geological map of Morocco (Hollard et al., 1985). The Aridal Formation is considered to represent middle and late Eocene times or sometimes all the Paleocene and Eocene, possibly even Miocene (Lindner and Querol, 1971; Ratschiller, 1967, 1970; Rjimati et al., 2008). However, the Gueran fauna that includes both protocetid and basilosaurid archaeocetes is most likely Bartonian (Gingerich and Zouhri, 2015).
Gingerich and Zouhri (2015) summarized the geological context of the Gueran deposits. The stratigraphical section measured in 2016 on the northeastern flank of the depression (Fig. 2) is shorter than that of Ratschiller (1967, 1970), but described in more detail. The fossiliferous bed of interest here is ∼ 11 m above the base of the section. It is a 1-m-thick white to light-gray sandstone that is clayey, silty, poorly-sorted, and very fine to coarse in grain size (Fig. 2).
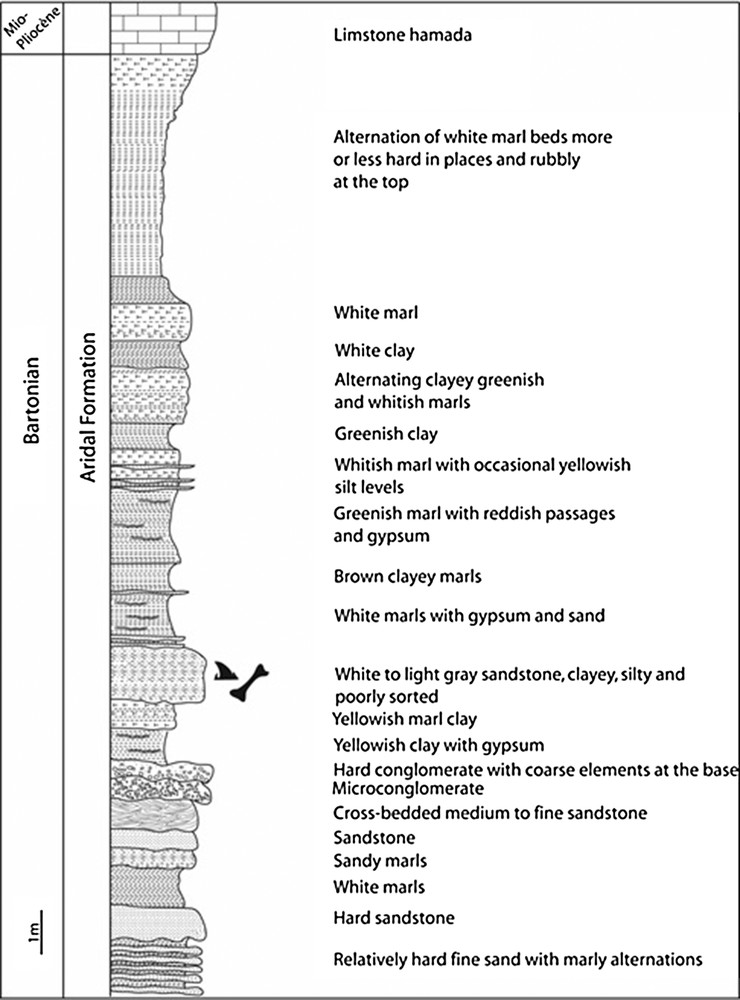
Stratigraphical section in the northeastern wall of the Gueran Depression, measured at 25.15042° N and 13.85990° W. The Aridal Formation as a whole is thicker than the 30.6 m exposed here, where the base of the formation is covered and the top truncated by an unconformity between the Aridal Formation and the overlying Miocene–Pliocene deposits.
3 Materials and methods
About 350 specimens were recorded in the fossil assemblage of the sabkha of Gueran. They consist mostly of incomplete articulated skeletons, skulls, mandibles, teeth, abundant vertebrae, and a few limb bones of archaeocete whales. Elasmobranchs are represented by abundant teeth and spines. Other taxa are represented by less abundant and fragmentary material.
Here we present the overall composition of the fauna and draw a detailed map of the spatial distribution of all vertebrate fossils found in the sabkha of Gueran, focusing on the actinopterygians, sauropsids, and mammals. All fossils come from a single 1-m-thick sandstone interval that extends for about 20 km near the base of the escarpments surrounding the Gueran Depression.
Cetacean remains dominate the fossil assemblage (78% of the specimens), with a predominance of Basilosauridae (69%), and a smaller proportion of Protocetidae (7%). A small proportion (2%) of fragmentary specimens is recorded as cetaceans indet. (Fig. 3). This low percentage illustrates the good state of preservation of fossils from the sabkha of Gueran. Other identified fossils (16% of all specimens, Fig. 3) belong to different taxa: actinopterygians, turtles, crocodiles, snakes, seabirds, and proboscideans. Teeth and spines of selacians are abundant, including large teeth of Otodus. However, chondrichthyan remains were not considered in the statistical analysis expressed in Fig. 3. Actinopterygian remains represent 4% of the fossil record. They mainly consist of vertebrae, rostra, tooth-bearing bones, and spines. Turtle remains represent 2% of the fossil record, crocodile remains 3% and bird remains about 2%. Seabird remains consist of beak fragments and a long bone of the family Pelagornithidae, which is also known in the Thanetian/Ypresian of the Oulad Abdoun Basin and in the Priabonian of Ad-Dakhla. Remains of proboscideans are very rare in Gueran. They consist of fragmentary teeth.
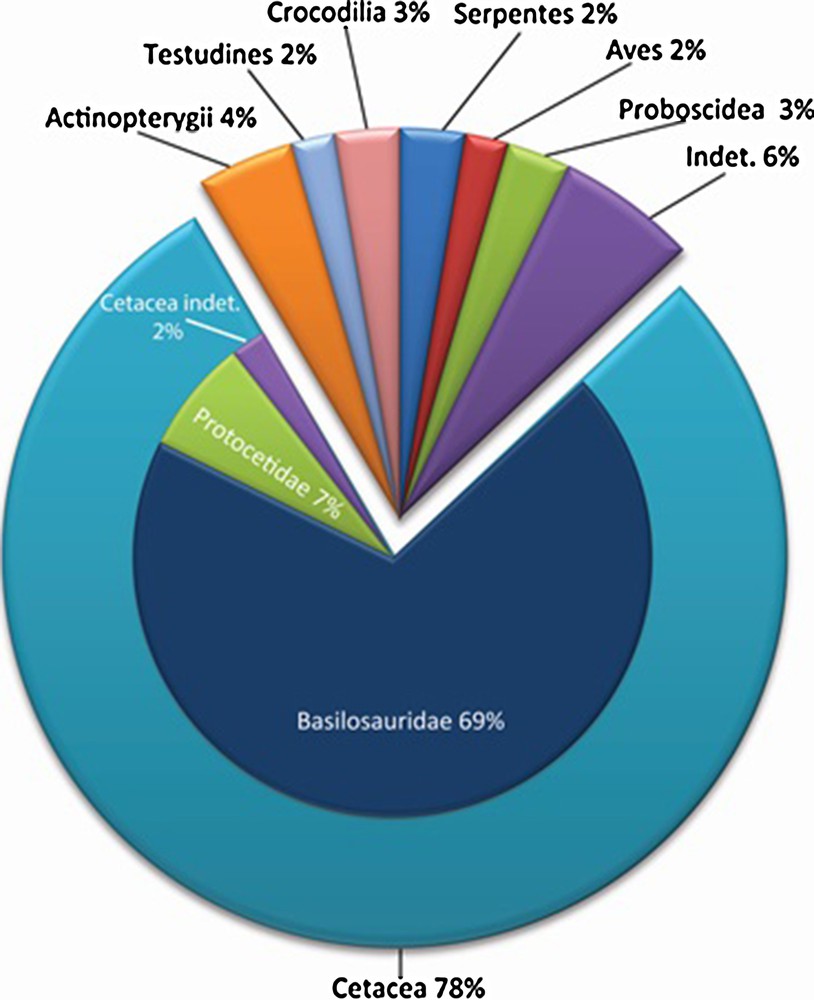
Overall composition of the vertebrate assemblage of the sabkha of Gueran. Isolated teeth and spines of elasmobranchs are not considered.
The exact position of each fossil was recorded using the Global Positioning System (GPS). Detailed taphonomic observations and preliminary identification of each fossil were made on the field. The good preservation of bones made it possible to taxonomically identify most specimens on the field. For other specimens, mechanical preparation and detailed study were essential for their taxonomic determination.
Specimens were plotted on a 1:100,000 topographic map built from SRTM data (SRTM3 USGS version 3). The positions of the fossils on the map are shown in Fig. 4.
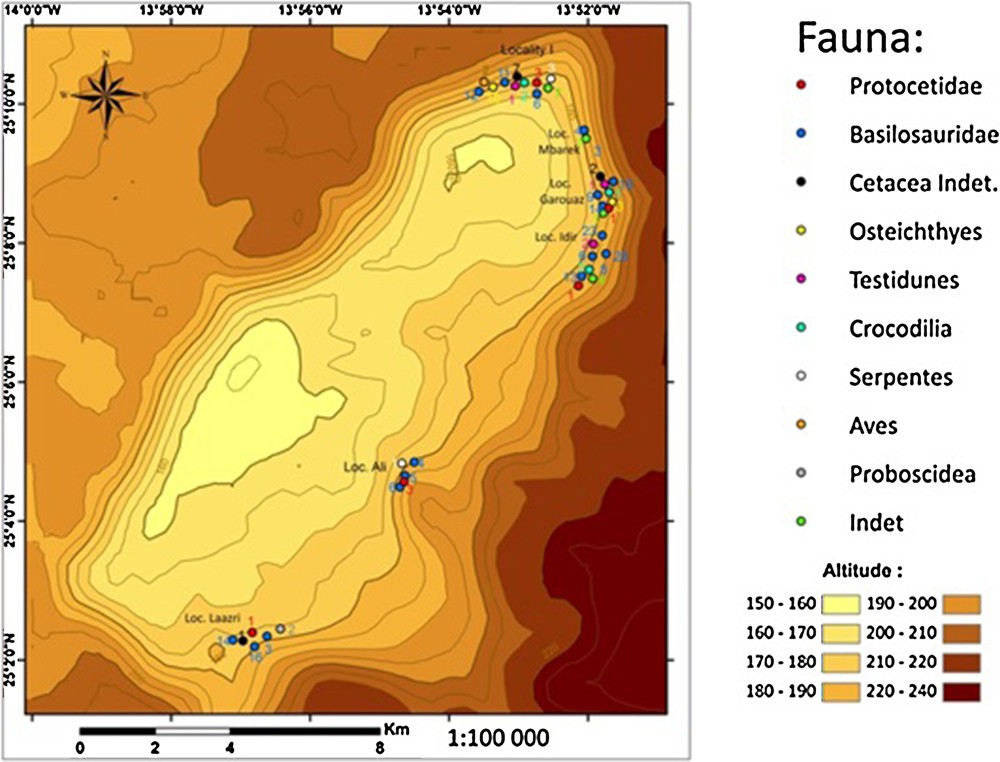
Map of Gueran Depression showing the global spatial distribution of fossil vertebrates. The numbers indicate the total number of specimens for a given taxon by location.
4 Results and discussion
4.1 Fossils distribution
Vertebrate fossils are not distributed uniformly at Gueran (Fig. 4). In some areas, such as the western and southwestern flanks of the sabkha, fossils are apparently totally absent. In the southeastern part, there is a low concentration of fossils (Ali site). On the other hand, a great concentration of fossils is observed in the northeastern part and particularly in the Mbarek, Garouaz, and Idir sites at the E–NE of the sabkha (see Fig. 4 for site locations). A high concentration is also recorded in the Laazri site, in the southern part of the depression.
When the distribution of the spatial position in the sabkha of each identified taxon is analyzed separately, the following distributions are observed.
Basilosauridae have been identified practically in all fossiliferous sites whereas Protocetidae seem to be confined to the north and northeast (Idir, Garouaz, M’barek, and Locality I sites). Among the Basilosauridae, Chrysocetus and Platyosphys, the two most common cetacean genera of Gueran, have the same distribution.
Fossil actinopterygians, turtles and crocodiles seem to have the same distribution as the Protocetidae. Two exceptions are seabirds and proboscideans, represented by a very small number of specimens. Birds are only known from Locality I in the North of Gueran, whereas proboscideans are found only in the Laazri locality in the South of Gueran.
All specimens described here are permanently housed in the Department of Geology, Faculty of Science Aïn Chock (FSAC), Hassan II University of Casablanca (Morocco).
4.2 Gueran fauna
As noted above (Fig. 3), the fauna from Gueran is composed of elasmobranchs, actinopterygians (perciforms, siluriforms, and Cylindracanthus), sauropsids (turtles, crocodilians, palaeophiid snakes and pelagornithid seabirds), and mammals (archaeocete whales and proboscideans).
Mammals are mostly represented by archaeocete whales. In a preliminary study, Gingerich and Zouhri (2015) recognized three species of Protocetidae and three species of Basilosauridae. Sirenians are currently absent from the locality, while eosirenids are known from Ad-Dakhla (Priabonian, Morocco) and Wadi el Hitan (Egypt). As in Ad-Dakhla, terrestrial mammals are represented by proboscidean tooth fragments, referred to as Proboscidea indet.
At least 12 species of elasmobranchs have been identified, including several new species currently under study. It should be noted that fossils were only collected by surface picking, which explains the absence of the small taxa. This elasmobranch fossil assemblage shows similarity with the fauna recovered in the Midawara Formation (Egypt), uppermost Lutetian–lowermost Bartonian (e.g., “Carcharias” koerti), even if some of their taxa (e.g., Tethylamna cf. twiggsensis; Hemipristis curvatus) are reminiscent of the younger fauna recovered in the Gehannam Formation in Egypt (around the Bartonian/Priabonian boundary).
The actinopterygian fauna is only known from incomplete and isolated remains, including rostra of Cylindracanthus, a siluriform spine possibly related to ariids, and vertebrae, fin spines, and jaw bones of perciforms (including scombrids). These taxa are common in Eocene marine paleoenvironments, in Africa (e.g., Egypt, Morocco, Namibia: Adnet et al., 2010; Böhm, 1926; Zalmout and Gingerich, 2012; Zouhri et al., 2017) and Europe (e.g., Belgium, England, Casier, 1966; Leriche, 1905). The taxonomic determinations are too imprecise to refine the biogeographical and biostratigraphical implications.
Turtles from Gueran are only represented by isolated and incomplete postcranial remains, attributed to a cheloniid and a dermochelyid. Both taxa correspond to deep-sea forms and have also been recorded in the Priabonian beds of Ad-Dakhla and Fayum (Zouhri et al., 2017). A probable littoral pleurodire is also present, possibly representing either one podocnemidid form from Ad-Dakhla and Fayum, or a bothremydid from another locality.
The crocodilian material belongs to at least two longirostrine species. Specimens from the same individual (anterior and mid-portion of a left maxilla; posterior portion of a left maxilla; mid-portion of a right maxilla) and a portion of a left dentary were assigned to a gavialoid. These remains from Gueran strongly resemble Eogavialis africanum (Andrews, 1901) from the Priabonian and Rupelian of Fayum (Egypt) (Müller, 1927). However, these specimens are considered as Gavialoidea indet because their fragmentary nature precludes precise taxonomic assignment. Compared to gavialoid remains from same age of Dur At-Talah (Bartonian of southern Libya) (Llinas Agrasar, 2004), no clear differences can be noted. Unfortunately, the remains are too poor to provide a deeper comparison. The few teeth found in younger levels in Ad-Dakhla (Morocco) with their robust shape resemble those found in the gavialoid from Gueran (Zouhri et al., 2017). Unfortunately, other remains from this locality are uniquely postcranial material that bears few diagnostic characters; clear comparisons are thus not possible.
The sabkha of Gueran is the second locality in the Moroccan Sahara to have yielded remains of the palaeophiid snake Pterosphenus. Indeed, given the limited nature of the material and its incompleteness, and despite the uncertainty regarding the attribution of isolated vertebrae to one of the species of the genus Pterosphenus, we cautiously attribute the material of Gueran to Pterosphenus cf. schweinfurthi. It should be recalled that the locality of Garitas, geographically close to Ad-Dakhla city, has given some vertebrae of this genus. The Bedbone 1 in the Ad-Dakhla area, from which the palaeophiid fossils are coming from, is slightly younger than the fossiliferous level of Gueran; Zouhri et al. (2014) assigned a Priabonian age to Bedbone 1, while the fossiliferous level at Gueran would be of Bartonian age (Gingerich and Zouhri, 2015).
Seabirds are represented in the sabkha of Gueran by a fragmentary maxillary rostrum attributed to the pseudo-toothed birds (Pelagornithidae). The Gueran specimen constitutes the earliest occurrence of the genus Pelagornis. The wingspan of the Gueran specimen was probably between 5 and 6 m, and ranked in the upper size range for pseudo-toothed birds, together with Pelagornis chilensis (Mayr and Rubilar-Rogers, 2010) and the largest known flying bird Pelagornis sandersi (Ksepka, 2014). Two morphotypes can be distinguished within the Pelagornithidae (Bourdon, 2011; Bourdon et al., 2010). The first morphotype corresponds to the genus Dasornis, which includes small, medium (albatross-sized) and large forms, and is characterized by the primitive morphology of the wing bones compared to Neogene pelagornithids (Bourdon et al., 2010). Dasornis occurs in late Paleocene/early Eocene deposits of Morocco (Bourdon et al., 2010) and in the early Eocene of England (Harrison and Walker, 1976; Mayr, 2008). In contrast, the Pelagornis morphotype only includes gigantic forms that were highly specialized for soaring flight, and appears in the late Oligocene (Ksepka, 2014). Pelagornithid remains of uncertain affinities have been described between the middle Eocene and the middle Oligocene, which show some, but not all, derived features of Pelagornis (e.g., Bourdon and Cappetta, 2012; Cenizo et al., 2015; Mayr and Smith, 2010; Mayr and Zvonok, 2012; Mayr et al., 2008). The late Eocene fragmentary remains from Ad-Dakhla (Morocco) could possibly fit in this intermediate morphology (Zouhri et al., 2017). The Gueran specimen is late middle Eocene, and provides evidence that the Pelagornis morphotype appears in the fossil record more than 10 million years earlier than previously thought. It also indicates that giant species of Pelagornis coexisted with less derived pelagornithids showing an intermediate morphology between Dasornis and Pelagornis.
4.3 Geological age of the Gueran fauna
Based on the archaeocete assemblage characterized by the presence of Protocetidae and Basilosauridae, Gingerich and Zouhri (2015) assigned a Bartonian age to the unique fossiliferous level of Gueran. The shark and ray assemblage from Gueran, even incomplete with lack of small and medium-sized elasmobranch species, is rather similar to the fauna recovered in the Midawara Formation of Wadi El Rayan, Egypt (Fig. 5; Underwood et al., 2011), dated as uppermost Lutetian–lowermost Bartonian. Most taxa are similar in the two faunas, except the presence in Gueran of a small unidentified Otodus (Carcharocles) species and a possible different medium-sized carcharhinid, both unknown in Egypt. The presence of “Carcharias” koerti, Galeocerdo cf. eaglesomi and the lack of large modern Carcharhinus and Misrichthys suggest that these fossiliferous deposits are older than the units A–C of the Gehannam Formation of Wadi el Hitan in Egypt (uppermost Bartonian).
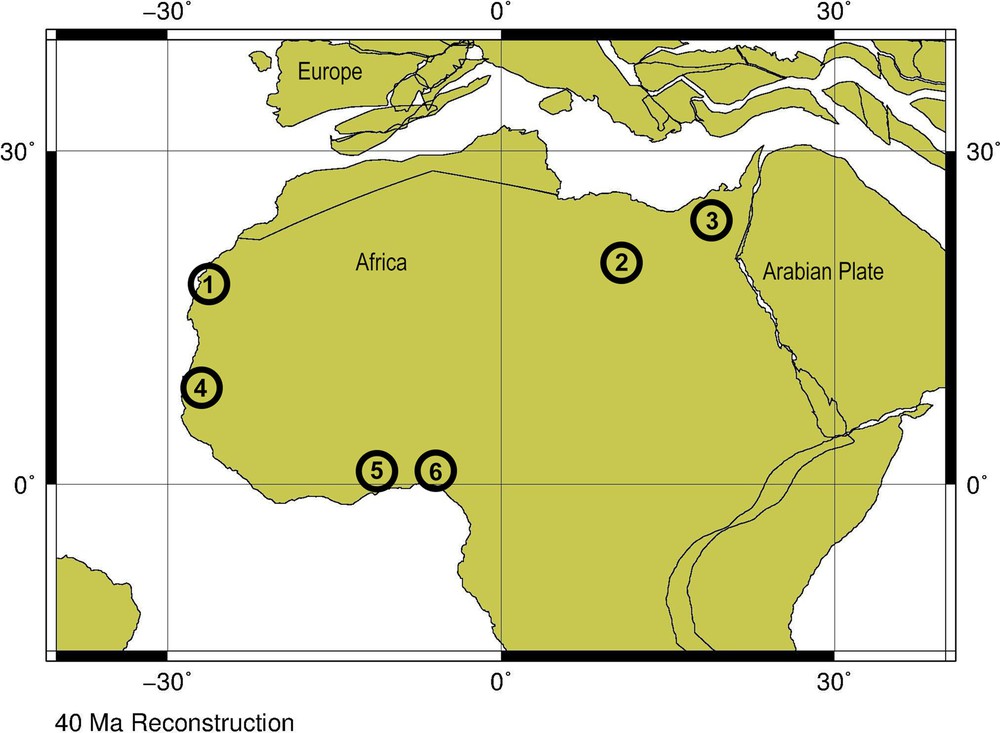
Bartonian palaeogeographic map of North Africa showing the position of (1) Gueran and Ad-Dakhla, (2) Dur at-Talah and (3) Fayum localities, (4) the Ndomor Diop site, and Tiavandou site in Senegal, (5) Kpogamé in Togo, (6) Ameke in southern Nigeria. ODSN Plate Tectonic Reconstruction Service (2011), modified.
It is therefore likely that the Gueran deposits are contemporaneous with the Midawara Formation thus uppermost Lutetian–lowermost Bartonian or slightly younger if we consider the greater abundance of Otodus (Carcharocles) cf. Sokolowi and Tethylamna cf. twiggsensis in the deposits analysed here. This age range is consistent with the Bartonian age suggested by Gingerich and Zouhri (2015) based on the study of the archaeocete fauna. The presence of the other taxa does not contradict this dating.
4.4 Paleobiogeography
Protocetid and basilosaurid archaeocetes had cosmopolitan distribution by the later part of the middle to late Eocene. The archaeocete assemblage of Gueran, which consists of species from these two groups, highlights the centrality of Gueran biogeographically for understanding the evolutionary transition from foot-powered swimming in Protocetidae to the tail-powered swimming of Basilosauridae and all later Cetaceans. The Eocene and Oligocene gavialoids are particularly scarce, and only known (Fig. 5) in the Priabonian and Rupelian Fayum beds (Egypt) (Müller, 1927), Priabonian of Ad-Dakhla (Morocco) (Zouhri et al., 2017), Bartonian of Dur At-Talah (southern Libya) (Llinas Agrasar, 2004), and possibly from the early–middle Eocene of England (Brochu, 2007). Late Eocene gavialoids are thus of particular interest, as the dispersal of the gavialoids to South America probably occurred during this period (Salas-Gismondi et al., 2016). The presence of gavialoids in the western part of the African continent could be an interesting element for a dispersal hypothesis of first Cenozoic South American gavialoids.
Pseudo-toothed birds of the genus Pelagornis were cosmopolitan during the Neogene, with the exception of Antarctica (see references above). Avian remains from Gueran provide evidence that the first appearance of Pelagornis is in the late middle Eocene (Bartonian) of North Africa. The second oldest record of this taxon comes from the late Oligocene (Chattian) of North America (Ksepka, 2014). These two records suggest that Pelagornis reached a wide distribution millions of years before the beginning of the Neogene. However, no precise biogeographical interpretations (e.g., an origin of Pelagornis in Africa) can be provided based on these records, because species of Pelagornis were highly efficient gliders that could disperse over thousands of kilometers (Ksepka, 2014).
4.5 Paleoenvironment
The elasmobranch and actinopterygian fauna is formed by marine taxa (e.g., lamniforms, carcharhiniforms, rhinopristiforms, Cylindracanthus, possible ariid siluriforms), usually encountered in shallow environments. Turtles from Gueran are represented by two forms of deep-sea and probably by a third littoral form. The unique extant gavialoid species, Gavialis gangeticus, lives in freshwater environments, but nearly all extinct species older than Miocene have been found in marine sediments. Therefore, Eocene–Oligocene gavialoids were probably marine, living in shallow water environments, likely in estuaries, deltas, and freshwater habitats. Palaeophiids were highly aquatic snakes. They lived mostly in shallow marine water, but they were capable of entering estuaries, deltas, and perhaps even coastal marshes (Parmley and DeVore, 2005; Rage, 1983). Mangroves were suggested as probable habitats for palaeophiids (Houssaye et al., 2013; Westgate and Gee, 1990). Therefore, palaeophiids were able to live in marine, brackish, and freshwater environments. However, if nearshore habitats were clearly preferred by palaeophiids, Pterosphenus cf. schucherti was reported from a locality corresponding to an open marine environment (Hutchinson, 1985). Pelagic seabirds of the genus Pelagornis were able of highly efficient gliding flight, and exploited a long-range soaring strategy similar to that of extant albatrosses (Diomedeidae) (Ksepka, 2014). They were unable of sustained flapping flight and practiced either dynamic soaring using the vertical gradient of wind velocity, or wave-slope soaring using updrafts over waves, like extant albatrosses (Richardson, 2011). The occurrence of pseudo-toothed birds in coastal marine sediments is not surprising, however. Abundant pelagornithid remains are usually found in deposits that correspond to shallow marine environments, because of the proximity of the breeding areas (Bourdon et al., 2010).
5 Conclusions
The Eocene marine fossil record is relatively well documented in the eastern part of the Tethys (India and Pakistan Subcontinent) and Near East (Egypt) in contrast with the southwestern margin of the Tethys, including the extant Maghreb (Tunisia, Algeria, and Morocco) and the adjacent West African Atlantic margin. The Middle-upper Eocene faunas discovered in the Ad-Dakhla area and the Bartonian middle Eocene fauna recently discovered in Gueran and reported here allow filling this gap. Even though taxa from Gueran are not accurately identified, the fauna shows biogeographic affinities with middle Eocene levels of Egypt and Libya, especially the Midawara Formation (Egypt) and this finding supports a close biogeographical connection between faunas from southeastern and southwestern coasts of the Mediterranean Sea.
This study will provide an appropriate basis for systematic, paleoecological and taphonomic studies as well as a possible conservation of the paleontological heritage of this important fossil site.
Acknowledgements
The authors have a special thought for our colleague Jean-Claude Rage who recently passed away. We thank Anne-Lise Folie, Cécilia Cousin (Institut royal des sciences naturelles de Belgique, Brussels), Philippe Béarez (UMR 7209, Muséum national d’histoire naturelle, Paris), Gaël Clément (UMR 7207, MNHN), Patrice Pruvost and Zora Gabsi (UMR 7208, MNHN) for access to collections. We are grateful to Philippe Béarez and François Meunier (UMR 7208, MNHN) for fruitful discussions, to Hélène Bourget (UMR 7207, MNHN) for the preparation of the material, and to Philippe Loubry (UMR 7207, MNHN) for taking photographs. We thank Mouloud Benammi, André Michard, and Isabelle Manighetti for reviews improving the text. Fieldwork research was supported by GRANT #9765-15 from the National Geographic Society, and by the Museum of Paleontology, University of Michigan, Ann Arbor, and by Hassan II University of Casablanca, Morocco.