


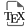
 CC-BY 4.0
CC-BY 4.0
1. Introduction
The mid-Cretaceous Blue Marls Formation of the Vocontian Basin (SE-France) is a thick sequence (650–800 m cumulated thickness) dominated by grey-colored pelagic to hemipelagic marls that were deposited in a fast subsiding basin [Bréhéret 1995, Friès 1986]. These deposits very well recorded both local and global environmental changes [Bréhéret 1995, Friès 1986]. Consequently, two Global Boundary Stratotype Sections and Points (GSSPs), namely the Col de Pré-Guittard section for the base of the Albian Stage [Kennedy et al. 2017] and the Mont Risou section for the base of the Cenomanian stage [Kennedy et al. 2004] are located in the Blue Marls Formation. One of the most remarkable features of the Blue Marls Formation is the recurrence of pluridecimetric dark colored layers, which have been collectively termed “organic levels” (OL), though the organic matter (OM) enrichment in these levels is highly variable [Bréhéret 1995, Caillaud et al. 2022]. All Cretaceous oceanic anoxic events (OAEs) were recorded in the Vocontian Basin [Leckie et al. 2002, Föllmi 2012], but in particular OAEs 1a to 1d and OAE2 are characterized by organic-rich deposits within the Blue Marls Formation. Nevertheless, additional OLs not related to global events are also observed.
Using sedimentation rate, grain size analysis, Rock-Eval analysis, trace metal content, and preliminary biomarker data, Caillaud et al. [2022] compared six of these OLs, namely the Goguel Level, Niveau Noir, Fallot Interval, Jacob Level, Kilian Level, and Paquier Level (Supplementary Figure 1). They concluded that the OLs were formed under variable conditions, but that in the absence of high primary productivity and bottom water anoxia, the supposedly “favorable” factors (e.g. rapid burial, sediment condensation, dominance of refractory terrestrial OM, …), only caused modest OM enrichments in the sediment [Caillaud et al. 2022]. We here present the continuation of the work by Caillaud et al. [2022] with detailed molecular fossil results of the same six OLs. These data complement the previous dataset and allow to reconsider the depositional model of some of these OLs.
2. Materials and methods
2.1. Sampling and bulk analysis
For bulk analysis, more than 130 samples were collected from 8 sections located in the Vocontian Basin (Figure 1, Supplementary Figure 2). Detailed information on the different sampled OLs and sections can be found in Supplementary text 1 and in Caillaud et al. [2022]. For each sample, 500 g–1 kg of rock were sampled. The surface sediment was removed on a thickness of 30–50 cm, in order to sample material not affected by weathering. The rock samples were kept in aluminum foils to avoid contamination. All samples were characterized by Rock-Eval analysis at Sorbonne University (ISTeP) using a RE6 device and the bulk rock method [Caillaud et al. 2022].
(a) Paleogeographic map of Late Aptian [simplified from Golonka 2007] showing the location of the Vocontian Basin (VB) and the main other areas cited in the text. Central Atlantic Ocean: ODP Sites 1049 and 1276, DSDP Site 545; UMB: Umbria-Marche Basin; IB: Ionian Basin. (b) Map of the geological context of the Vocontian Basin during the Aptian times. Modified from Friès and Parize [2003]. AB: l’Arboudeysse; Br: les Briers; GL: Glaise; ND: Notre Dame; PG: Pré-Guittard; Ra: Ravel; SA: les Sauzeries; SC: Serre Chaitieu; SJ: Saint Jaume; St: Saint-André-les-Alpes; TA: Tarendol.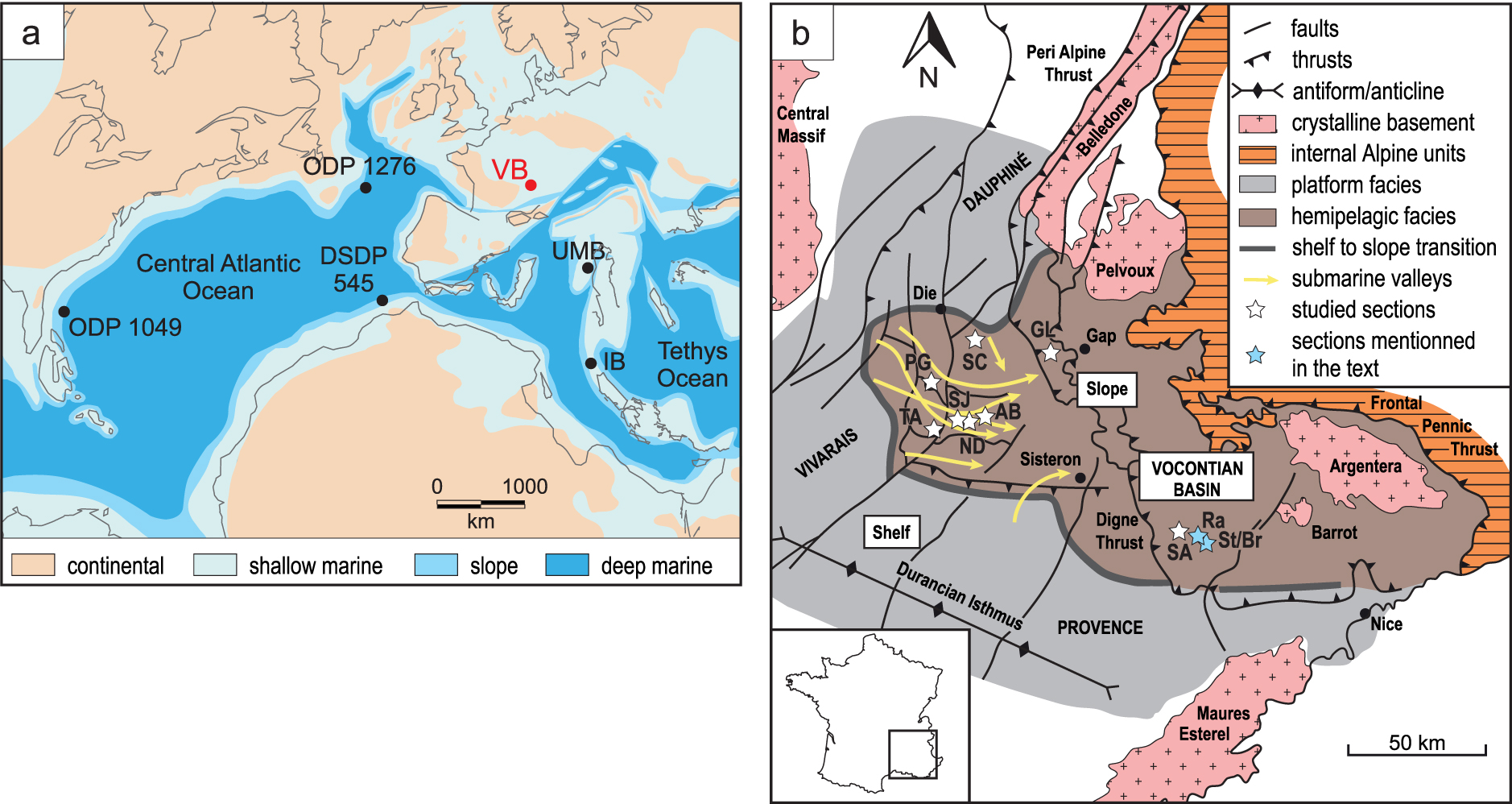
2.2. Biomarker analysis
64 samples, corresponding to black shales, dark-colored marls and bioturbated marls (Supplementary Figure 2), were selected for molecular fossil analysis. The procedure for lipid biomarker analysis is detailed in Supplementary text 2. Briefly, between 50 and 70 g of sediments were extracted using a mixture of dichloromethane and methanol 2:1 v/v with an accelerated solvent extractor. Maltenes-like fractions were recovered after reducing the total extracts to dryness by rotary evaporation and solubilisation in cyclohexane. The maltenes-like were separated in three fractions over an activated silica column using solvents of increasing polarity. The aliphatic and aromatic fractions were analyzed by gas chromatography–mass spectrometry (Supplementary text 2).
3. Results
3.1. Bulk OM
The total organic carbon (TOC) and results of Rock-Eval analysis of the total sample set were previously described [Caillaud et al. 2022]. Only the most important features of the samples selected for the present study are briefly recalled here. The highest TOC contents are observed in the samples from the Goguel Level at les Sauzeries section (av. 3.3%), followed by the Paquier Level (av. 2.7%). The Goguel Level at Saint Jaume and Notre Dame sections, part of the Fallot Interval, the Jacob Level, and the Kilian Level show TOC contents between 1 and 2%. The Goguel Level at Glaise section, the upper part of the Fallot Interval, and the Niveau Noir show TOC contents lower than 1%, which are comparable to the TOC content of the surrounding marls. Generally, the Hydrogen Indices (HI) are positively correlated with the TOC contents (Figure 2; Supplementary Figure 3). The highest HI are observed in the Goguel Level at les Sauzeries (387 mgHC/gTOC on average) and Paquier Level (290 mgHC/gTOC), indicating type II kerogen. The other sections are characterized by low to very low HI (67–202 mgHC/gTOC), reflecting type III to type II–III kerogen. The majority of samples bears Tmax values below 435 °C suggesting thermal immaturity. However, the Tmax values from Glaise and les Sauzeries sections are generally higher (435 and 439 °C, respectively), indicating that the OM has reached early oil-generating window.
Cross plot of the total organic carbon (TOC) content and Hydrogen Index (HI) for the rock samples of the Blue Marls Formation. The two trends depicted by the arrows are discussed in the text. See electronic version for colors.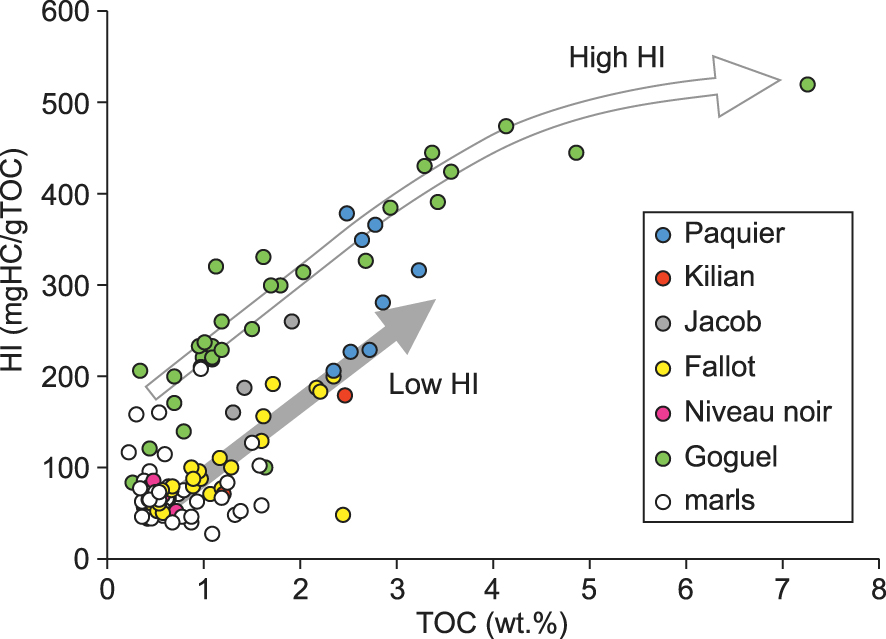
Two parallel trends are observed for the positive correlation between TOC content and HI (Figure 2; Supplementary Figure 3). They are termed as “high HI” and “low HI” for the sake of simplicity. The high HI trend includes samples from the Goguel Level at all sections plus a few samples from the Paquier Level. The low HI trend includes samples from the Kilian Level, Fallot Interval, Niveau Noir, part of the Paquier Level, and some samples from the Goguel Level at Saint Jaume section. The few samples from the Jacob Level are located in between these two trends. These two trends are also visible when Rock-Eval data from other studies of the Blue Marls Formation [Bréhéret 1994, Giraud et al. 2018, Westermann et al. 2013] are included.
3.2. Molecular fossils
A large variety of lipid biomarkers was found in the saturated and aromatic fractions of the studied samples. The molecular fossil content is similar in all samples. However, a few samples contain additional compounds, as will be described below. A detailed description of compounds distributions is presented in Supplementary text 3.
3.2.1. Linear and branched alkanes
Linear alkanes (n-alkanes) generally dominate the saturated fractions. n-Alkanes comprise short chain compounds from C13 to C22, with a maximum in C16 or C17 (Figure 3), which are generally ascribed to phytoplanktic sources [Blumer et al. 1971, Giger et al. 1980], and long chain compounds from C23 to C33, presenting a predominance of odd-carbon-numbered compounds. Though odd-carbon-numbered long-chain n-alkanes may originate from micro- or macroalgae [e.g. Aichner et al. 2010, Allard and Templier 2000], these compounds mainly derive from the epicuticular waxes of terrestrial plants [Eglinton and Hamilton 1967]. The terrestrial vs. aquatic ratio [TAR; Bourbonniere and Meyers 1996] is generally lower than 0.4, reflecting a low contribution of long chain n-alkanes (Supplementary Table 1). Marl samples at Pré-Guittard have an average TAR of 5, and TAR > 0.4 are observed in the Fallot Interval and Paquier Level. The lowest TAR values are observed in the Goguel Level at les Sauzeries and Saint Jaume. The predominance of odd-carbon-numbered long n-alkanes, as indicated by OEP27 [Scalan and Smith 1970], is present in the majority of samples (Supplementary Table 1). The highest OEP27 are observed in the Fallot Interval (av. 1.9) and Paquier Level (av. 1.8). In the samples of the Goguel Level at les Sauzeries and Glaise, the OEP27 ∼1 is ascribed to the high thermal maturity of the OM in these sections.
Mass chromatogram m∕z 57 of selected aliphatic fractions showing the distribution of acyclic compounds. (a) Sample TAFE2OG003—Kilian Level; (b) sample ARBOG002A—Paquier Level. Pr: pristane; Ph: pyhtane; TMI: 2,6,15,19-tetramethylicosane; PMI: 2,6,10,15,19-pentamethylicosane; ETMI: 10-ethyl-2,6,15,19-tetramethylicosane; PMH: pentamethylhenicosane, Cx = number of carbon atoms.
Series of 2-methyl- and 3-methyl-alkanes, ranging from C15 to C21, maximizing around C17, are observed in most of the samples. These compounds generally derive from branched iso- and anteiso- fatty acids of gram positive bacteria [Goossens et al. 1986, Kaneda 1991]. The highest relative abundances of branched alkanes are observed in the Niveau Noir while low abundances are observed in the Fallot Interval (Supplementary Table 1).
3.2.2. Acyclic and monocyclic isoprenoids
Acyclic isoprenoids mainly consist of a series of head-to-tail linked (regular) compounds ranging from C14 to C21 and are dominated by pristane (Pr) and phytane (Ph). Regular isoprenoids are generally less abundant than short n-alkanes (Supplementary Table 1; Figure 3a). In the Paquier and Jacob Levels, however, the relative abundance of regular isoprenoids is higher (Figure 3, Supplementary Table 1) and 2,6,10-trimethyltetradecane (C16) is the dominant regular isoprenoid in the samples of the Jacob Level. The samples from the Paquier Level also contain a series of tail-to-tail linked C24–C26 irregular isoprenoids (TMI, PMI and ETMI) which were previously described by Vink et al. [1998, Figure 3] in the same interval. An additional compound belonging to this series, pentamethylhenicosane (PMH), is also tentatively identified (Supplementary Figure 4). Three isoprenoid compounds with a cyclohexyl ring ranging from C17 to C19 previously described by Vink et al. [1998] are also present with significant contents in the samples of the Paquier Level. Numerous other compounds showing comparable structures were tentatively identified based on their mass spectra (Supplementary Figure 5). The latter compounds, ranging from C14 to C20, are also present in one sample of the Jacob Level.
Pristane and phytane are often related to chlorophyll [Didyk et al. 1978], which is present in many photosynthetic organisms, but can also originate from the degradation of tocopherols, carotenoids or biphytane structures [Goossens et al. 1984, Li et al. 1995, ten Haven et al. 1987], so they are not source-specific. PMI derives from the membrane lipids of archaea [Holzer et al. 1979, Brassell et al. 1981]. PMI and other archaea-derived isoprenoids, such as biphytanes, presenting a highly depleted carbon isotopic signature point to methanotrophic archaea as the source of these compounds [e.g. Elvert et al. 1999, Birgel et al. 2006]. Nevertheless, in deposits coeval to the Paquier Level, phytane, PMI and biphytanes have a heavy carbon isotopic signature [Kuypers et al. 2002]. Isotopically heavy PMI is present in the deep part of the chemocline of the Black Sea and Cariaco Basin, and is related to chemoautotrophic archaea [Wakeham et al. 2007, 2012]. Vink et al. [1998] suggested that TMI and ETMI originated from the same source as PMI. Similarly, we consider likely that the other isoprenoids, including monocyclic compounds, present in abundance in the samples of the Paquier and Jacob Levels, also originate from chemoautotrophic archaea.
Most of the samples have a Pr/Ph ratio higher than 1 (Figure 4, Supplementary Table 1). The highest values of Pr/Ph, up to 8.6, are observed in the marl samples at Pré-Guittard, followed by the Fallot Interval. Only some samples from the Jacob and Paquier Levels have Pr/Ph close to or lower than 1 (Figure 4, Supplementary Table 1).
Pristane/nC17 vs. phytane/nC18 cross plot of samples from the Vocontian Basin. Partially filled symbols correspond to marls collected close to the organic levels.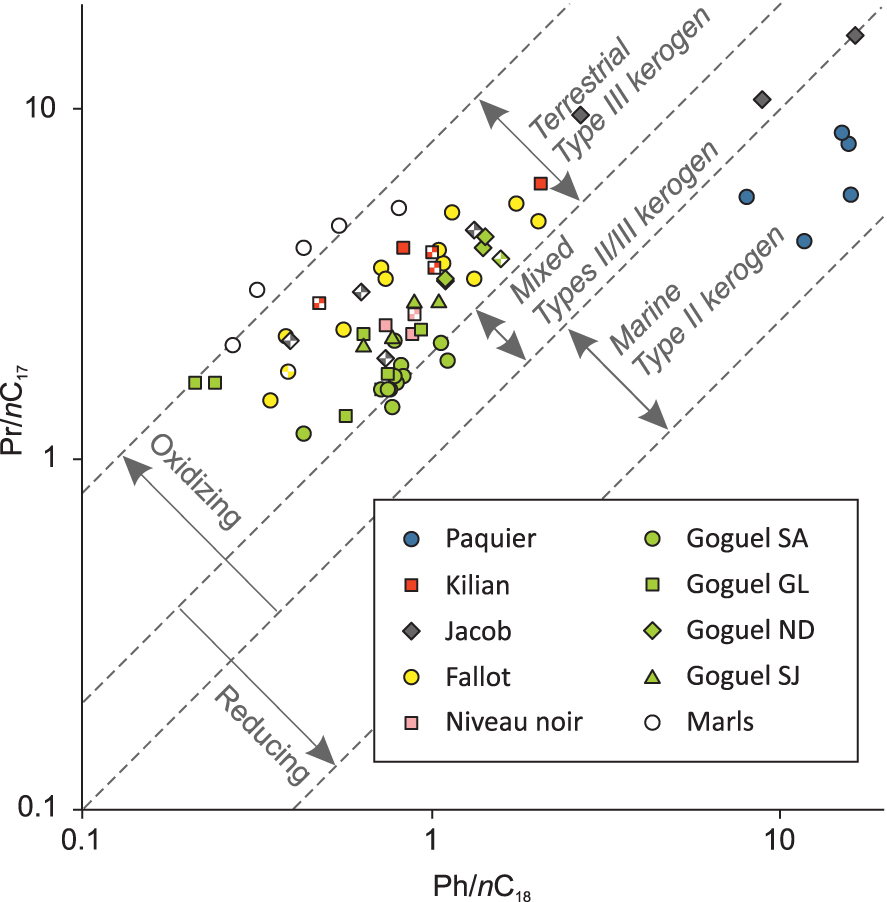
3.2.3. Steroids
A wide variety of steroids is observed in most of the samples. The compounds include diaster-13(17)-enes, diasteranes, regular steranes, ring-C monoaromatic steroids and triaromatic steroids, as well as methylated counterparts. The highest relative abundances of steroids are observed in the Paquier Level and the Goguel Level at Notre Dame. Low relative abundances of steroids are generally observed in the marls.
Regular steranes are generally dominated by C29 compounds (Figure 5, Supplementary Figure 6). In the Paquier Level, however, C27 isomers are dominating regular steranes. The relative abundance of C27, C28 and C29 regular steranes [Huang and Meinschein 1979] suggests a mixed contribution of algal and terrestrial OM for all samples (Figure 5). Nevertheless, 24-n-propylcholestanes are observed in significant proportion in most of the samples (Supplementary Table 1). Precursors of 24-n-propylcholestane have been observed in marine algae [Raederstorff and Rohmer 1984], desmosponges [Zumberge 2019] and a foraminifer [Grabenstatter et al. 2013]. 24-n-Propylcholestane could also result from diagenetic methylation of C29 sterols [Bobrovskiy et al. 2021]. Nevertheless, 24-n-propylcholestanes are generally considered as indicative of deposition under marine conditions [Moldowan et al. 1985]. The highest abundances of 24-n-propylcholestanes are observed in the Paquier Level (Supplementary Table 1).
Ternary plot showing the distribution of C27, C28, and C29 regular steranes in the aliphatic fractions. Legend of symbols as in Figure 4.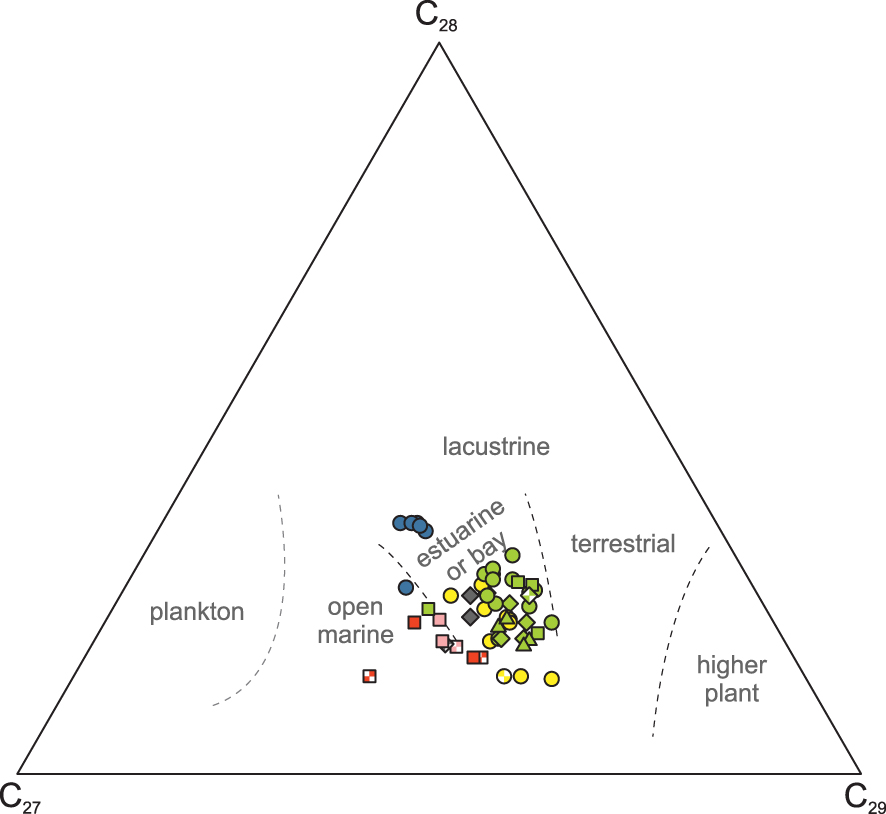
Methylsteroids comprise saturated and aromatic compounds methylated at position 2-, 3-, and 4-, as well as dinosteroids (based on the 4,23,24-trimethylcholestane skeleton; Supplementary Figures 6, 10). 2-Methyl- and 3-methyl-steroids originate from diagenetic rearrangements in the sediment [Summons and Capon 1988, 1991] and seem present in relatively stable proportion in the samples. Conversely, 4-methylsteroids and dinosteroids mainly originate from dinoflagellates [Summons et al. 1987], and are present in variable proportion in the samples. The highest proportions of 4α-methyl-steranes, dinosteranes and triaromatic dinosteroids are observed in the Paquier and Goguel Levels at Notre Dame and Saint Jaume.
3.2.4. Hopanoids
A wide variety of hopanoids is observed in the samples (Supplementary text 3). The dominant hopanoids are saturated αβ-hopanes ranging from C27 to C35 (C28 absent) with a maximum in C30 and a progressive decrease of the abundance of homohopanoids with increasing chain length (Figure 6). The low abundance of C34 and C35 hopanes is indicative of a deposition in non-hypersaline conditions [ten Haven et al. 1985]. The distribution of hopanoids isomers is comparable in most of the samples and reflects the thermal immaturity of the OM; however, the distribution differs at les Sauzeries and Glaise sections, reflecting the higher thermal maturity at these locations (Supplementary Table 1).
Partial mass chromatograms showing the distribution of (a) hopanes and moretanes (m∕z 191) and (b) methylhopanes (m∕z 205). Sample NDOG003—Goguel Level.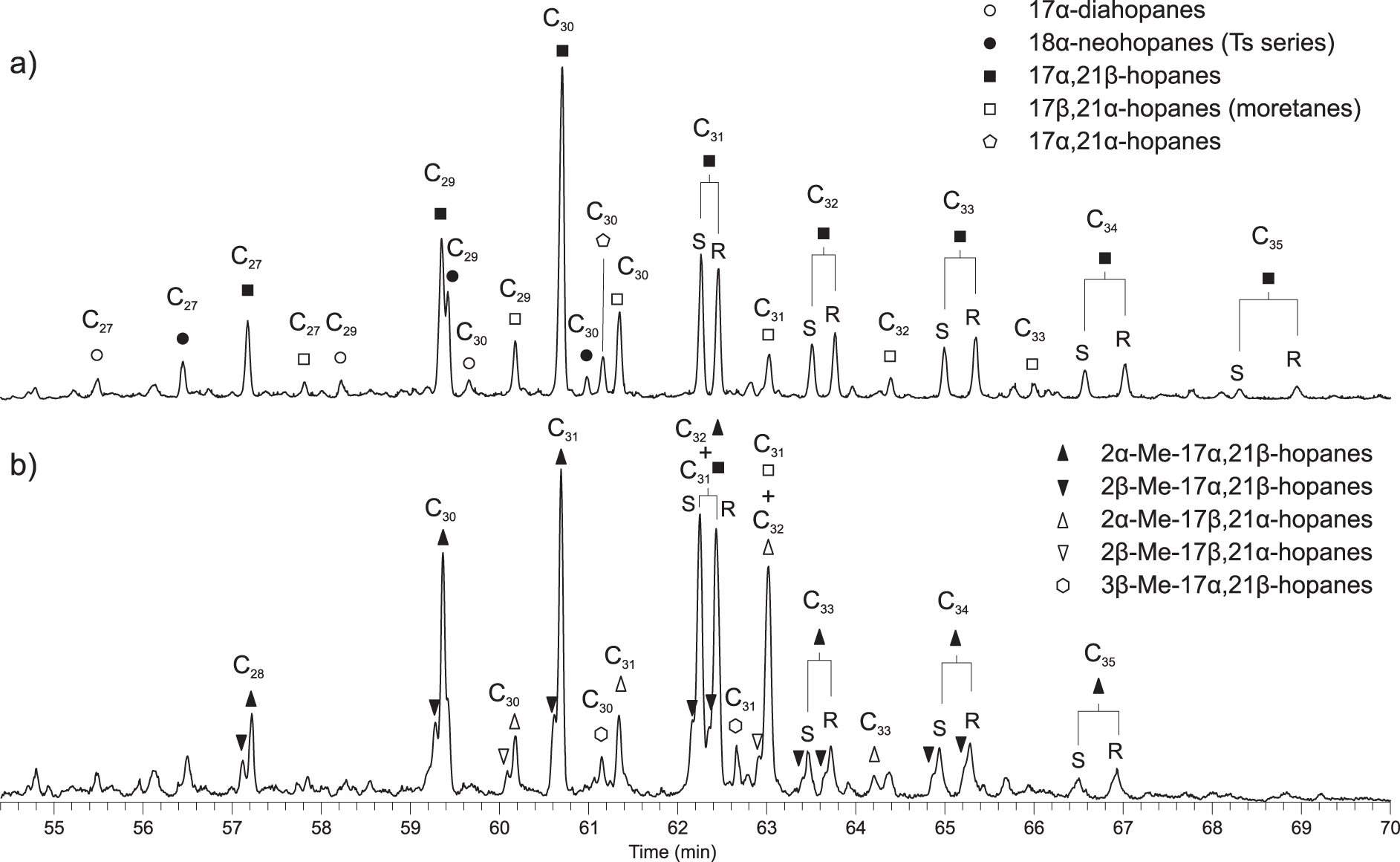
2-Methyl hopanes ranging from C28 to C35, with a maximum in C31, are present in significant proportion in most of the studied samples from the Goguel Level, with 2-methyl hopane index [2-MHI; Ando et al. 2022] ranging between 6 and 23 (Figure 6; Supplementary Table 1). If present in the other OLs, the abundance of 2-methyl hopanes is very low. 2-Methyl hopanes are absent from the extract of marl samples (Supplementary Table 1). Traces of 3-methyl-hopanes are detected in many samples (Figure 6).
Desmethyl hopanoids mostly originate from biohopanoids present in the cell walls of many bacteria, including cyanobacteria [Ourisson and Rohmer 1992, Talbot et al. 2008, Kusch and Rush 2022], so desmethyl hopanes are poorly specific bacterial markers. In contrast, methyl-hopanes are more specific: though their origin is still discussed, 2-methyl-hopanes mainly originate from cyanobacteria and alphaproteobacteria [Summons et al. 1999, Naafs et al. 2022, Kusch and Rush 2022], while 3-methylhopanes originate from aerobic methanothrophs [Neunlist and Rohmer 1985, Cordova-Gonzalez et al. 2020].
3.2.5. Aryl isoprenoids
Aryl isoprenoids are detected in most of the OLs, though generally in low abundance. This series ranges from C13 to C20, with the C24 and C29 compounds also often present in low proportion. Values of the aryl isoprenoid ratio [AIR; Schwark and Frimmel 2004] are generally higher than 1, reflecting the dominance of short aryl isoprenoids (C13–C15); nevertheless, AIR are lower than 1 in the Paquier Level (Supplementary Table 1). The highest proportions of arylisoprenoids are observed in the Paquier Level, as well as the Goguel Level at Glaise and Notre Dame.
Aryl isoprenoids are often considered to derive from carotenoids of anoxygenic photosynthetic bacteria [Summons and Powell 1987, Brocks and Schaeffer 2008]. Nevertheless, other sources, in particular the aromatisation of carotene, are also likely when these compounds are observed in rocks deposited in well oxygenated environments [Koopmans et al. 1996b]. From the very low abundance of aryl isoprenoids longer than C20, the absence of C40 aryl isoprenoids as well as of commonly observed isorenieratene derivatives [Koopmans et al. 1996a], the arylisoprenoids observed in the sediments of the Vocontian Basin likely do not originate from anoxygenic photosynthetic bacteria. In particular, the abundant aryl isoprenoids and substituted alkylbenzenes (Supplementary text 3) observed in the samples from the Paquier Level could originate from the degradation and aromatisation of archaea-derived biphytanyl compounds.
3.2.6. Other terpenoids
Terpenoids of microbial origin, namely dammar-13(17)-enes and 13β,17α(H)-dammaranes [Meunier-Christmann et al. 1991] are observed in the samples of the Paquier Level only, while cheilanthanes [Peters et al. 2005] are widespread in the samples from the Goguel Level, Niveau Noir, and the Paquier Level.
The aromatic terpenoids retene, cadalene, and 6-isopropyl-1-isohexyl-2-methylnaphthalene are detected in most of the samples. These three compounds mainly derive from higher plant terpenoids and are thus generally related to a terrestrial source [van Aarssen et al. 2000]. Their relative abundance is maximum in the marl samples from Pré-Guittard and minimum in the Goguel Level at Glaise and les Sauzeries. High relative abundances are also observed in the Paquier Level (Supplementary Table 1). Cadalene, a non-specific terrestrial plant biomarker [van Aarssen et al. 2000] is largely dominant in all the samples, though its relative abundance is slightly lower in the Goguel Level at les Sauzeries section.
Series of methylated 2-methyl-(trimethyltridecyl)-chromans (MTTC) are present in most of the samples, except those from Pré-Guittard, les Sauzeries and Glaise sections. These compounds likely originate from the rearrangement of the phytol side chain of chlorophyll [Li et al. 1995] but are widely used as paleosalinity indicators. The dominance of the 5,7,8-trimethyl-isomer in the samples from the Vocontian Basin indicates normal salinity conditions [Sinninghe Damsté et al. 1993].
3.2.7. Polycyclic aromatic hydrocarbons
Condensed polycyclic aromatic hydrocarbons ranging from triaromatic (phenanthrene and anthracene) to heptaaromatic (coronene) are detected in most of the samples (Supplementary Table 1, Supplementary Figure 13). These compounds generally have a pyrogenic origin [Wakeham et al. 1980, bin Abas et al. 1995] and can be used as tracers of terrigenous inputs. The highest relative abundances of PAHs are associated to a predominance of benzo[ghi]perylene and coronene, and are observed in the marl samples from Pré-Guittard section, Tarandol (Kilian and Jacob Levels), and Serre Chaitieu sections (Fallot Interval). Low relative abundances of PAHs are observed in the samples of the Goguel Level, Niveau Noir and Paquier Level.
Dibenzofuran is detected in most of the samples, except the marls from Pré-Guittard. Its highest abundance is observed in the Niveau Noir. Dibenzofuran and its alkylated counterparts have been related to lichens and can be used as tracers of terrigenous inputs [Radke et al. 2000]. Nevertheless, our data suggest that dibenzofuran here results from the degradation of an algal or microbial biomass (see Section 4).
3.2.8. Sulfur containing compounds
Organo-sulfur compounds (OSCs) are present in all the samples from the Paquier Level. They correspond to a C20 isoprenoid thiophene and several isomers of C20 isoprenoid benzothiophenes. OSCs are not observed in the other samples.
4. Discussion
4.1. Thermal maturity
Based on Rock-Eval analysis and molecular indicators (Supplementary Table 1), the majority of the studied outcrops are thermally immature, allowing to compare the biomarker content with each other in these different settings. Nevertheless, consistent with previous studies [Bréhéret 1995], our data indicate that the oil-generating window was reached at les Sauzeries and Glaise sections (Supplementary Table 1). The biomarker content of the Goguel Level at these two outcrops must therefore be considered in the light of this higher thermal maturity.
4.2. Comparison of the different OLs
Previous characterisations of the OM in the Blue Marls Formation based on palynofacies, palynology or organic geochemistry, showed that the OM was mainly of marine origin, with variable contributions of terrestrial plant debris [Bodin et al. 2023, Bréhéret 1994, Friedrich et al. 2003, Heimhofer et al. 2006, Herrle et al. 2010, 2003b, Kuypers et al. 2002, Okano et al. 2008b, Tribovillard and Gorin 1991]. The biomarkers identified in the present study confirm the mainly autochthonous origin of the OM, while the contributions from higher plants are present, but are low in most of the samples. According to previous studies (op. cit.), the variations of the organic content reflect several factors: sea-level evolution during the Aptian to Albian interval, variations of primary productivity, orbitally induced climate fluctuations, and oceanic anoxic events. In addition, turbiditic corridors are well documented in the western part of the Vocontian Basin [Bréhéret 1995, Friès 1986, Friès and Parize 2003], so that the location within the basin might also influence the organic content.
The Blue Marls Formation is characterized by alternations of ligh and dark levels related to orbitally induced climate oscillations [Ait-Itto et al. 2023, Charbonnier et al. 2023, Gale et al. 2011]. These orbitally induced oscillations are also visible in the OLs, as the latter frequently present alternations of black shales or dark-colored laminated marls and bioturbated marls (Supplementary Figure 2). Since the sampling for molecular fossil analyses was mainly focussed on the darkest sub-levels, the discussion below concentrates on the conditions most prone to OM deposition.
4.2.1. Hemipelagic marls
Though located in the west part of the Vocontian Basin, the Pré-Guittard section does not show major turbidites in the sampled interval (Supplementary Figure 2) and can therefore be considered as representative of the hemipelagic sedimentation. The very low TOC content and HI, as well as high OI [Caillaud et al. 2022, Ait-Itto et al. 2023], are typical for highly degraded OM (type IV). Such observation is consistent with the high Pr/Ph ratio and the absence of C35 homohopanes, indicative of well oxygenated conditions in the sediment [Didyk et al. 1978, Peters and Moldowan 1991]. The very low abundance of steroids, the notable contribution of resistant compounds, such as long n-alkanes from terrestrial plant waxes and fire-derived PAHs, and the CPI of n-alkanes close to 1, are also consistent with extensive biodegradation [Hoefs et al. 2002, Peters et al. 2005]. Dibenzofuran is hardly observed at Pré-Guittard, while other land-sourced compounds, e.g. long-chain n-alkanes, cadalene and PAHs are relatively abundant. The latter compounds are mainly delivered to the marine environment as aerosols [Simoneit and Mazurek 1982, Simoneit 1984], while dibenzofuran could be delivered by rivers, thus explaining this discrepancy. Nevertheless, an alternative source for dibenzofuran is suggested by our data (see below). It should be noted that in the marl samples from the Pré-Guittard section, the terrestrial signature is higher than in the marl samples from the other sections (Supplementary Table 1).
4.2.2. Goguel Level
The Goguel Level is recording the Oceanic Anoxic Event (OAE) 1a in the Vocontian Basin. The OAE1a is a global event characterized by a carbonate crisis, deposition of organic-rich sediments in marine settings and a marked negative carbon isotope excursion in both carbonates and OM [see Föllmi 2012, for a review].
The Goguel Level was sampled at four different outcrops (Figure 1, Supplementary Figure 2). Les Sauzeries was in a distal setting, far from detrital influences [Bréhéret 1995]. Nevertheless, the OM is thermally mature. Glaise section is as well supposed as a distal setting, but was influenced by detrital inputs, as indicated by the fine turbidites. The OM is thermally mature, as for les Sauzeries. Saint Jaume and Notre Dame sections were located in a more proximal setting and contain more detrital deposits, as indicated by the numerous turbidites, though the turbidites are thin at Saint Jaume and relatively thick à Notre Dame. As a result of the detrital influence, lower TOC and HI are observed at Glaise, Notre Dame and Saint Jaume compared to les Sauzeries (Supplementary Figure 3); still, the four outcrops all belong to the “High HI” trend, the same trend as observed in the coeval Selli Level, in the Umbria-Marche Basin [Baudin et al. 1998].
Consistent with previous molecular fossil studies [Heimhofer et al. 2004, Okano et al. 2008a, b, Ando et al. 2022, 2017], the molecular signature of the Goguel Level is marked by the predominance of autochthonous algal and microbial OM, without marked difference in the four studied sections (Supplementary Table 1). In particular, all four sections show a similar low abundance of land-derived inputs. The moderate amounts of dibenzofurans in these sections suggest that this compound is not land-derived. The high sterane/hopane ratio in the least thermally mature sections reflects an important contribution of eukaryotes and in particular a significant contribution of dinoflagellates, which can be related to the observation of dinocysts and acritarchs in the palynofacies [Heimhofer et al. 2006, Ando et al. 2022]. A high abundance of dinosteroids was similarly observed in the coeval mid-Pacific organic-rich deposits of Shatsky Rise [Dumitrescu and Brassell 2005].
Though indicators of severe water column anoxia were described in the coeval deposits from northern Tethys [Pancost et al. 2004, van Breugel et al. 2007], and sediment anoxia at Shatsky Rise [Dumitrescu and Brassell 2005], molecular fossils of euxinia or anoxia, like aryl isoprenoids are not observed in the Vocontian Basin. The Goguel Level presents a “paper shale” texture, and the benthic fauna is overall rare [Friès 1986, Bréhéret 1995, Giraud et al. 2018]. Moreover, the dominance of fluorescing amorphous OM in the palynofacies and the high HI indicate a good degree of OM preservation [Heimhofer et al. 2004, 2006, Ando et al. 2022]. These data point to anoxic conditions at the sediment water interface (Figure 7a), consistent with the high values of the C35 homohopane ratio [Peters and Moldowan 1991]. Episodic oxygenation is indicated by the intermittent presence of benthic foraminifera or bioturbation in the Goguel Level [Friès 1986, Bréhéret 1995, Giraud et al. 2018], but the low enrichments of redox-sensitive trace elements [Westermann et al. 2013, Caillaud et al. 2020] could reflect a basin reservoir effect [Caillaud et al. 2020]. Despite the dominance of anoxia in the sediment, high Pr/Ph are observed in the Goguel Level [this study; Okano et al. 2008b, Ando et al. 2017]; it thus appears that the Pr/Ph ratio is not a good indicator of oxygenation conditions in this level. The high Pr/Ph ratio could be related to an abundance of pristane precursors, such as tocopherols, in the organic flux to the sediment [Goossens et al. 1984], or to peculiar diagenetic conditions, as also suggested by the abundance of MTTC [Li et al. 1995].
Depositional model for the different OLs.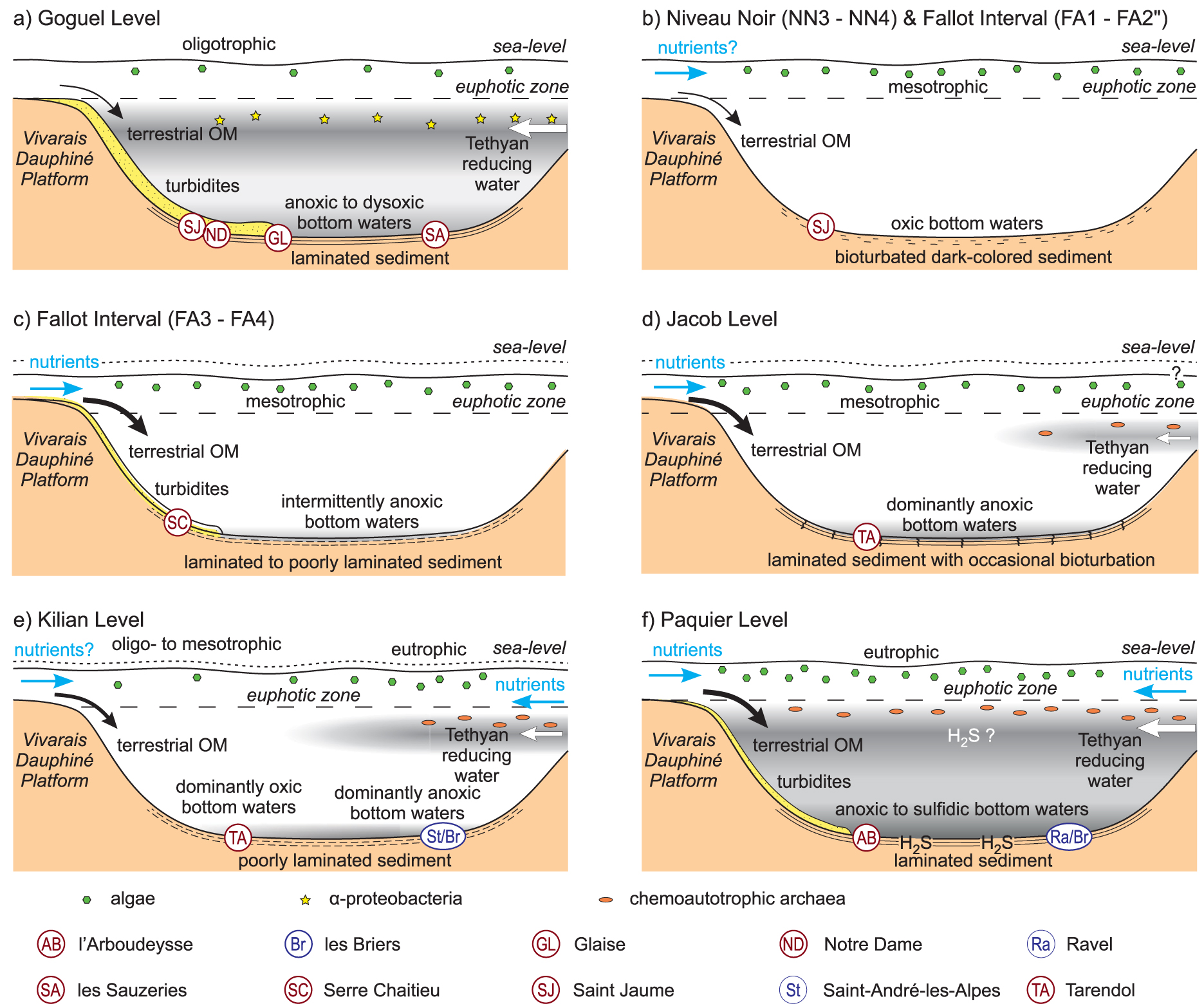
The abundance of 2-methylhopanoids at the four studied sections is distinctive of the Goguel Level compared to the other OLs of the Vocontian Basin. The abundance of 2-methylhopanoids has been previously described in different organic-rich deposits of OAE1a [Kuypers et al. 2004b, Dumitrescu and Brassell 2005, 2006, Karakitsios et al. 2018, Castro et al. 2019], but also in the deposits of the Toarcian OAE [Farrimond et al. 1994, Blumenberg and Wiese 2012, Ruebsam et al. 2018, Ajuaba et al. 2022], and OAE2 [Farrimond et al. 1990, Kuypers et al. 2004a, Forster et al. 2008]. The significance of elevated concentrations of 2-methyl hopanoids is still highly discussed [Kusch and Rush 2022]. Nevertheless, a recent review suggests that the abundance of 2-methylhopanoids during these events reflects an expansion of marine anoxygenic photoautotrophic α-proteobacteria as a response to water column deoxygenation [Naafs et al. 2022]. A high contribution of α-proteobacteria during deposition of the Goguel Level is consistent with the oligothrophic conditions indicated by the nannofossil content [Herrle and Mutterlose 2003, Giraud et al. 2018]. Oligotrophic conditions are generally not prone to favor deoxygenation of the water column or sediment. Conversely, high productivity is documented at the beginning of OAE1a in the Tethys Ocean [Aguado et al. 2014, Sabatino et al. 2015]. For this reason, it appears more likely that the deoxygenation in the Vocontian Basin resulted from the spreading of oxygen-poor waters originating from the Tethys Ocean (Figure 7a). The Goguel Level was deposited during a transgressive period [Supplementary Figure 14; Rubino 1989, Friès and Parize 2003, Ferry et al. 2022], which favored communications between the Tethys Ocean and the Vocontian Basin. The gradient of the phosphorus content in the sediment [Westermann et al. 2013], and the observation of maximum 2-MHI in the most basinal section at les Sauzeries, are consistent with this scenario. Sedimentary condensation likely also favored OM enrichment [Supplementary Figure 14; Caillaud et al. 2022, 2020].
4.2.3. Niveau noir
The Niveau Noir has not been correlated with other organic-rich levels outside the Vocontian Basin and is considered a local event. In the present study, only the two upper sub-levels of the Niveau Noir were analyzed (NN3 and NN4, Supplementary Figure 2). The biomarker content of these two samples indicate that the OM is mainly of algal to bacterial origin. The low abundances of continent-derived long-chain n-alkanes and cadalene, as well as fire-derived PAHs, are consistent with the absence of turbidites from the Niveau Noir, though turbidites are present in other parts of Saint Jaume section. Molecular indicators of the redox state of the water column and sediment (homohopane ratio, AIR, …) indicate oxygenated conditions, consistent with the absence of redox-sensitive trace metal enrichment [Caillaud et al. 2022]. The abundance of branched alkanes suggests efficient bacterial activity under aerobic conditions. Nevertheless, the sterane/hopane ratio indicates significant input of eukaryotic biomass, including algae producing 4-methyl-steranes. The chain length distribution of steranes in the Niveau Noir is characterized by a high proportion of the C27 compounds (Figure 5). C27 sterols are produced by a variety of marine algae as well as zooplankton [Volkman 1986], and thus are not specific biomarkers. Nevertheless, cholesterol dominance was observed in the upper sediment or settling particles of several high productivity environments [e.g. Volkman et al. 1987, Colombo et al. 1996]. The high proportion of the C27 steranes in the Niveau Noir could therefore reflect high primary productivity, despite the low enrichment of trace elements sensitive to productivity [Caillaud et al. 2022]. Consistent with our interpretation, the nannofossil content of the NN4 level indicates higher primary productivity than for the surrounding marls [Herrle et al. 2010].
To summarize, the Niveau Noir resulted from an increase in surface primary productivity (Figure 7b, Supplementary Figure 14), possibly resulting from orbitally induced increases of nutrient delivery associated to continental runoff [Herrle et al. 2010]. Nevertheless, this increase in primary productivity was not sufficient to significantly alter the concentration of oxygen in the water column and sediment (Figure 7b). As a result, OM preservation was ultimately low. The Niveau Noir was deposited during a high sea level interval [Rubino 1989, Friès and Parize 2003], sediment condensation likely also contributed to OM enrichment [Supplementary Figure 14; Caillaud et al. 2022].
4.2.4. Fallot interval
The Fallot Interval has no recognised lateral equivalent outside the Vocontian Basin and is considered a local event. Though the biomarker content of the Fallot Interval is dominated by an algo-bacterial signature, the influence of terrestrial OM is more pronounced than in the other OLs. A progressive increase of the terrestrial influence from the base to the top of the Fallot Interval is indicated by the increase of TAR and of the relative abundance of PAHs, mirroring a decrease of the proportions of the marine steroids and of dibenzofuran (Supplementary Table 1). Consistently, several authors consider that the Fallot Interval was deposited during a period of falling relative sea level [Supplementary Figure 14; Rubino 1989, Friès and Parize 2003]. The values of the AIR and C35 homohopane ratio, as well as the absence of isorenieratane [Reitner et al. 2015], point to dysoxic to well oxygenated conditions, consistent with the presence of bioturbations and benthic foraminifera in most of the Fallot Interval [Bréhéret 1995, Friedrich et al. 2003]. The low HI and low abundance of steroids consitently indicate efficient OM degradation. The moderate sterane/hopane ratio suggests a moderate input of eukaryotic biomass, consistent with the mesotrophic conditions indicated by planktic foraminifera [Friedrich et al. 2003]. The latter authors suggested that OM enrichment in the sediment was related to moderately high primary productivity for the lower part of the Fallot Interval, but was linked to water column stagnation in the upper part. The molecular fossil content of the lower part of the Fallot Interval is consistent with the “productivity” model of Friedrich et al. [2003] and points to depositional conditions comparable to the Niveau Noir, but with a higher input of terrestrial OM (Figure 7b). Nevertheless, for the second part of the Fallot Interval, the stagnation model [Friedrich et al. 2003] is not supported by the redox proxies. Taking account of the lower hydrocarbon extraction yields from leaves and wood of terrestrial plants compared to algae [e.g. Cranwell et al. 1990, Bush and McInerney 2013, Jambrina-Enríquez et al. 2018], the biomarker content likely underestimates the proportion of terrestrial OM in the sediment. Though a detailed palynofacies analysis is needed to further support this hypothesis, the more pronounced terrestrial signature suggests that the upper part of the Fallot Interval results from efficient burial of terrestrial OM (Figure 7b, Supplementary Figure 14), as proposed by Caillaud et al. [2022].
4.2.5. Jacob Level
The Jacob Level is the first organic-rich deposit associated to OAE1b in the Vocontian Basin [Leckie et al. 2002, Coccioni et al. 2014]. Lateral equivalents of the Jacob Level have been mostly recognised in the Tethys Ocean and in the Pacific Ocean; it could correspond to a global event [see Bodin et al. 2023, for a review].
Debris of higher plants have been described in the Jacob Level [Barale and Bréhéret 1995], as well as abundant spores and pollen grains in the palynofacies [Bréhéret 1995, Heimhofer et al. 2006]. Consistent, PAHs, retene and cadalene are relatively abundant. Nevertheless, only the highest sample from the Jacob Level shows a high TAR, and the molecular signature is overall dominated by the algal contribution (Supplementary Table 1). Low proportions of dinosterane and medium values of the triaromatic dinosteroid (TADS, Supplementary text 2) ratio suggest medium to low contribution of dinoflagellates. Average C27 triaromatic steroids (TAS, Supplementary text 2) values also suggest an average productivity of coccolithophorids or prasinophytes. Consistent, the microfaunal content suggests mesotrophic conditions [Erbacher et al. 1998, Heimhofer et al. 2006]. The samples of the Jacob Level plot between the “high HI” and “low HI” trends (Figure 2), which suggests intermediate OM preservation, and is consistent with the variable proportion of amorphous OM in the palynofacies [Heimhofer et al. 2006]. The same conclusion can be reached based on the molecular signature: steranes and hopanes are relatively abundant but the homohopane index is null, suggesting OM degradation in an oxygenated sediment. The presence of benthic foraminifera in the Jacob Level [Erbacher et al. 1998] gives support to this latter interpretation.
A peculiar feature in the extracts of the Jacob Level is the abundance of acyclic and monocyclic isoprenoids. Though PMI and TMI are not observed in the Jacob Level [this study; Ando et al. 2022, 2017], we consider that the abundant isoprenoids likely derive from lipids of planktic chemoautotrophic archaea. The archaeal biomarkers PMI and TMI have been described in Livello 113, a lateral equivalent of the Jacob Level in Umbria-Marche Basin [Coccioni et al. 2014, Ferraro 2017]. Livello 113 was deposited as a result of moderate to high primary productivity and anoxia to dysoxia of the bottom water [Coccioni et al. 2014, Sabatino et al. 2015, Ferraro et al. 2020]. Planktic chemoautotrophic archaea could have been driven in the Vocontian Basin through intrusions of anoxic intermediate waters originating from the northern Tethyan margin. The presence of the Tethyan taxon Nannoconus in the Jacob Level [Herrle and Mutterlose 2003], is consistent with this scenario. Nevertheless, the water column was oxygenated in the Vocontian Basin, which led to efficient degradation of the archaeal lipids (Figure 7d).
Previous models have proposed that OM enrichment in the Jacob Level was the result of sediment condensation, low sea level, elevated detrital input, or water column anoxia [Bréhéret 1995, Erbacher et al. 1998, Heimhofer et al. 2006, Caillaud et al. 2022]. A short episode of temperature increase during this interval is documented in the Umbria-Marche Basin [Ferraro et al. 2020]. Increased continental runoff associated to a more humid climate could have increased the delivery of terrestrial OM, but also favoured primary productivity and bottom water stagnation in the Vocontian Basin [Figure 7d; Heimhofer et al. 2006]. The Jacob Level was deposited during a period of low sea level, which also favoured the delivery of continental OM to the Vocontian Basin [Figure 7d, Supplementary Figure 14; Friès and Parize 2003]. Nevertheless, the surrection of the west margin of the Vocontian Basin during the late Aptian–early Albian [Ferry et al. 2022] could have further contributed to the increased detrital flux.
4.2.6. Kilian Level
The Kilian Level is the second organic-rich deposit associated to OAE1b in the Vocontian Basin [Leckie et al. 2002, Trabucho Alexandre et al. 2011, Coccioni et al. 2014]. The Kilian Level is associated to a negative carbon isotope excursion, which has been recognised in numerous settings [Trabucho Alexandre et al. 2011, Bodin et al. 2023]. It corresponds to a global event.
Two samples from the Kilian Level were analyzed for their biomarker content and they show contrasting results: the lower one is comparable to the marls below, with a marked terrestrial signature (TAR, PAHs) and abundant hopanes, while the second sample has a reduced terrestrial contribution and a more marine affinity, but high abundance of plant terpenes. The relative abundance of steroids in the Kilian Level is low, but is higher than in the surrounding marls. The sterane/hopane ratio is high in the upper sample. Based on TADS values, the dinoflagellate contribution is low in both samples, but higher than in the surrounding marls, and lower than in the Jacob Level (Supplementary Table 1). The C27TAS ratio indicates a contribution of coccolithophorid or prasinophyte comparable in the Kilian Level and surrounding marls. The sterane distribution in the upper sample is dominated by cholestane, interpreted as indicative of increased primary productivity. Despite the differences between the two studied samples, the biomarker content is consistent with the notable occurrence of terrestrial palynomorphs in the palynofacies [Herrle et al. 2003b]. It also suggests low to medium primary productivity, consistent with the oligo- to mesotrophic conditions indicated by nannofossils [Herrle et al. 2003b]. No molecular indicators of water column anoxia are observed, though the small and poorly diversified planktic foraminifera suggest water column stratification or deoxygenation [Bréhéret 1995, Kennedy et al. 2000]. Though the Kilian Level is often laminated, the low C35 homohopane ratio indicates a relatively oxygenated sediment, consistent with the presence of benthic foraminifera [Herrle et al. 2003b] and modest enrichments in redox-sensitive trace elements [Caillaud et al. 2022, Wang et al. 2022]. Our observations differ from previous molecular studies of the Kilian Level at Saint-André-les-Alpes, located to the east of the Vocontian Basin, in a more basinal setting (Figure 1). A higher contribution of dinoflagellates was observed [Ando et al. 2017], with a general increase from the Jacob to the Kilian and Paquier Levels. In addition, PMI and TMI were present in significant concentration [Okano et al. 2008b].
In the Umbria-Marche Basin and in the Central Atlantic Ocean, the equivalents of the Kilian Level are dominated by marine OM and resulted from increases in surface productivity [Trabucho Alexandre et al. 2011, Coccioni et al. 2014]. They were deposited under anoxic bottom waters, though trace elements are not systematically enriched in the rocks [Trabucho Alexandre et al. 2011, Sabatino et al. 2015]. The Italian equivalent of the Kilian Level contains TMI and PMI, but no isorenieratane [Ferraro 2017].
Recent studies of les Briers outcrop, located to the east of the Vocontian Basin, suggest that the Kilian Level resulted from an episode of increased primary productivity and OM preservation, associated to pulses of continental runoff in the Vocontian Basin [Bodin et al. 2023]. The molecular data do not conflict with this model, but the presence of archaeal biomarkers in the eastern part of the basin suggest that the Vocontian Basin was also influenced by intrusions of reducing water from the Tethyan margin (Figure 7d, Supplementary Figure 14). These intrusions led to higher surface productivity, high abundance of dinoflagellates, presence of planktic archaea and possibly more reducing conditions in the water column in the eastern part of the Vocontian Basin, but not to the west (Figure 7d). The east–west decrease of TOC content in the Kilian Level [Caillaud et al. 2022, Bodin et al. 2023] and the absence of Nannoconus in the Kilian Level at Pré-Guittard and Palluel [Bréhéret 1995], in the west part of the Vocontian Basin, give support to this scenario.
4.2.7. Paquier Level
The Paquier Level is the most prominent of the organic-rich deposits associated to OAE1b in the Vocontian Basin [Bréhéret 1995, Coccioni et al. 2014]. Equivalents of the Paquier Level are recognised worldwide, in particular by its marked negative carbon isotope excursion in carbonate [Trabucho Alexandre et al. 2011, Coccioni et al. 2014; also see Bodin et al. 2023 for a review]. The Paquier Level is the only level in the OAE1b that records global ocean deoxygenation [Wang et al. 2022].
The molecular fossil content of the Paquier Level is characterized by the abundance of archaea-derived acyclic and monocyclic isoprenoids (Supplementary text 3). This feature was previously observed in Ravel section, located in a more basinal part of the Vocontian Basin [Vink et al. 1998, Kuypers et al. 2002, Okano et al. 2008b]. The high abundance of archea-derived phytane in the samples of the Paquier Level explains the low Pr/Ph, which are the lowest of our dataset (Supplementary Table 1). The Paquier Level is also characterized by a high abundance of steranes, a high sterane/hopane ratio and the highest abundances of n-propyl cholestanes, indicative of a high flux of a diversified eukaryotic biomass. The relative abundance of dinosteroids is high, as indicated by the TADS values [Ando et al. 2017], which can be related to the abundance of dinocysts in the palynofacies [Tribovillard and Gorin 1991, Kennedy et al. 2000]. The chain length distribution of steranes in the Paquier Level is characterized by the dominance of the C27 compounds, which we interpret as indicating high primary productivity. This interpretation is consistent with the microfaunal content indicating eutrophic conditions [Erbacher et al. 1998, Kennedy et al. 2000, Herrle et al. 2003b]. Though the molecular signature of marine organisms is dominant, the terrestrial contribution, indicated by the TAR, abundance of PAHs and of higher plant aromatic terpenoids, is also notable (Supplementary Table 1). These molecular data can be related to the presence of abundant terrestrial palynomorphs in the palynofacies [Tribovillard and Gorin 1991, Herrle et al. 2003b], and of occasional plant remains in the rocks [Bréhéret 1995]. The contribution of terrestrial OM in the Paquier Level has been observed in different locations of the Vocontian Basin, and cannot be ascribed to the relatively proximal setting of the studied section.
The high homohopane ratio and the presence of OSCs indicate sulfidic conditions in the sediment, possibly associated with bottom water anoxia (Figure 7f). This is consistent with the rarity of the benthic fauna [Bréhéret 1995, Erbacher et al. 1998], abundance of amorphous OM in the palynofacies [Tribovillard and Gorin 1991], and trace metal enrichment in the sediment [Benamara et al. 2020, Caillaud et al. 2022, Wang et al. 2022]. Nevertheless, no indicator of photic zone anoxia was detected, consistent with previous data [Kuypers et al. 2002].
Molecular fossil analyses were performed in several lateral equivalents of the Paquier Level, in the Umbria-Marche Basin [Coccioni et al. 2014], in the Ionian Basin [Tsikos et al. 2004] and in the Central Atlantic Ocean [Kuypers et al. 2002]. All these levels are characterized by the abundance of biomarkers derived from archaeal lipids, including PMI and TMI, and by the absence of isorenieratane [Kuypers et al. 2002, Tsikos et al. 2004, Ferraro 2017]. While in the Central Atlantic Ocean samples the steranes are dominated by the C29 compounds and 2-methylhopanes are present [Kuypers et al. 2002], the biomarker content in the Umbria-Marche Basin is more comparable with that of the Paquier Level, with a dominance of C27 steranes and possible absence of 2-methylhopanes [Ferraro 2017]. These lateral equivalents of the Paquier Level are all characterized by heavy organic carbon isotopic ratios (𝛿13Corg = −22 to − 17‰), indicative of a high contribution of chemoautotrophic archaea [Kuypers et al. 2002, Tsikos et al. 2004, Sabatino et al. 2015]. In the Paquier Level, however, heavy organic carbon isotopic ratios are only observed sporadically [Benamara et al. 2020, Ait-Itto et al. 2023, Bodin et al. 2023], indicating that archaea were rarely dominant in the Vocontian Basin.
Because of the high proportion of terrestrial OM, several authors proposed that the Paquier Level deposited as a result of increased input of nutrient to the ocean associated to a warm, humid climate and continental runoff [Herrle et al. 2003a, b, Benamara et al. 2020, Bodin et al. 2023]. The highest levels of primary productivity and water stratification would have been reached under favourable orbital conditions, at the peak of a long period of progressive warming [Wang et al. 2022, Bodin et al. 2023]. The surrection of the west margin of the Vocontian Basin during the late Aptian–early Albian [Ferry et al. 2022] possibly also contributed to the increased inputs of continental material during this period. The molecular fossil content does not conflict with the influence of terrestrial runoff. Nevertheless, the similarity of biomarker composition in the northern Tethyan margin and in the Vocontian Basin, as well as the abundance of the Tethyan nannofossil Nannoconus in certain layers of the Paquier Level [Bréhéret 1995], are consistent with the establishment of good connections between the Tethys Ocean and the Vocontian Basin (Figure 7f). The Paquier Level is interpreted as a 3rd order maximum flooding surface [Figure 7f, Supplementary Figure 14; Rubino 1989, Friès and Parize 2003]. Sediment and bottom water anoxia in the Vocontian Basin could therefore have resulted from the high primary productivity, nevertheless, an expansion of the Tethyan oxygen minimum zone associated to the eustatic rise also seems possible.
5. Conclusion
Basing on sedimentation rate, grain size analysis, Rock-Eval analysis, trace metal content, and preliminary biomarker data, Caillaud et al. [2022] came to the conclusion that organic matter (OM) enrichment within the Blue Marls Formation (Aptian–Albian) was only modest because the conditions in the Vocontian Basin were overall not favorable to massive burial of OM. This detailed study of the biomarker content in the Organic Levels confirms that surface productivity fluctuated mainly between oligotrophic and mesotrophic and that bottom waters were rarely reducing. As a result, the OLs of regional extension only show modest OM enrichments, associated to small increases of productivity (Niveau Noir) or of terrestrial OM inputs (Fallot Interval).
Larger-scale events are fundamentally different: the Goguel Level (OAE1a) and Jacob, Kilian, and Paquier Levels (OAE1b) present the same molecular peculiarities (abundance of 2-methylhopanoids and of archaeal lipids, respectively) as coeval deposits located outside the Vocontian Basin. These are strong arguments in favor of good connections between the Tethys Ocean and the Vocontian Basin during the deposition of these levels. In particular, a lateral spreading of the Tethyan oxygen minimum zone into the Vocontian Basin could explain the low sediment oxygenation despite moderate increases of surface productivity. Sea-level variations were previously considered as an important factor influencing OM enrichment, through sediment condensation and water column stratification. The present biomarker study highlights how sea-level variations also impact OM enrichment in a given sedimentary basin, through connections with surrounding basins. Such feature, well known in modern settings, such as the Cariaco Basin off Venezuela, are more difficult to document in ancient sedimentary basins.
Declaration of interests
The authors do not work for, advise, own shares in, or receive funds from any organization that could benefit from this article, and have declared no affiliations other than their research organizations.
Acknowledgements
We thank TOTAL S.A. (now TotalEnergy) for funding this research, M. Gentric for administrative management, M. Delattre and R. Abraham for technical support. P. Adam and P. Schaeffer (Institut de Chimie de Strasbourg) are acknowledged for their help in identifying compounds.