



 CC-BY 4.0
CC-BY 4.0
1. Introduction
After oxygen, silicon is the second most abundant element on Earth’s surface, and its importance in global geochemical cycles is considerable. Its geological cycle is closely intertwined with that of other elements. This is particularly true for carbon through the supergene weathering of silicate minerals, which leads to the capture of atmospheric carbon in the form of carbonates that are then stored at the bottom of the ocean.
At the oceanic scale, silicon is absorbed by microorganisms such as diatoms, which play a major role in biogeochemical cycles. On land, the terrestrial biogeochemical cycle of silicon remained largely understudied until the early 1990s. The pioneering work of Bartoli (Bartoli, 1983, and references herein) proposed a steady-state cycle model with four reservoirs (Figure 1): soil water, living biomass, dead biomass, and soil minerals. This system also included atmospheric inputs and outputs through dissolved silicon and particles exported to rivers and groundwater. Measurements of certain fluxes, such as litterfall, allowed Bartoli (1983) to demonstrate that silicon recycling by vegetation could regulate the silicon cycle in ecosystems, such as the deciduous forest of Housseras (eastern France). In other ecosystems, such as the coniferous forest of Biffontaine (also in eastern France), vegetation recycles very little silicon, and the silicon found in drainage water mainly originates from the dissolution of soil minerals.
The terrestrial biogeochemical cycle of Si according to Bartoli (1983).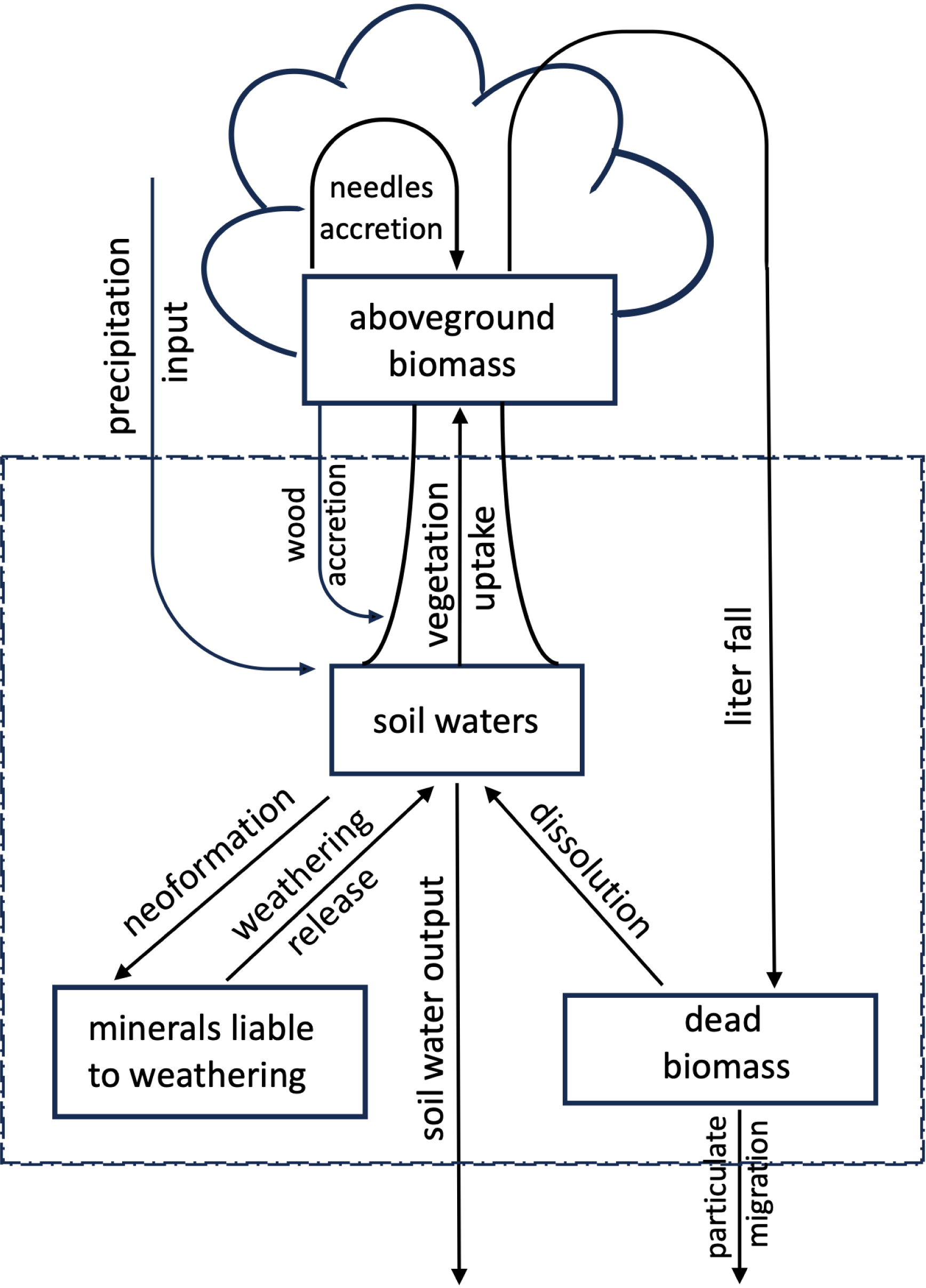
In plants, silicon is present in the form of amorphous silica particles, which range in size from a few micrometers to several tens of micrometers. These particles are called phytoliths. They have been known since the 19th century (Piperno, 2006), but a WOS (Web of Science) analysis shows that the number of publications with the keyword “phytolith” has increased significantly since the early 1990s, thus showing a momentum of interest in the scientific community (total counts = 3053; Figure 2). Although a great number of publications concern the application of phytoliths in the archaeology and paleoenvironmental disciplines, approximately 30% (883) are counted if the keyword “soil science” is added to “phytolith”. Additionally, a greater number of publications appear with the terms “silicon” and “agriculture” (8011; Figure 2b), reflecting the growing interest in Si in the fields of biology and agronomy.
Counts of article citations from Web of Science (up to June 2025); (a) using the keyword “phytoliths”; (b) using the keywords “silicon” and “agriculture”.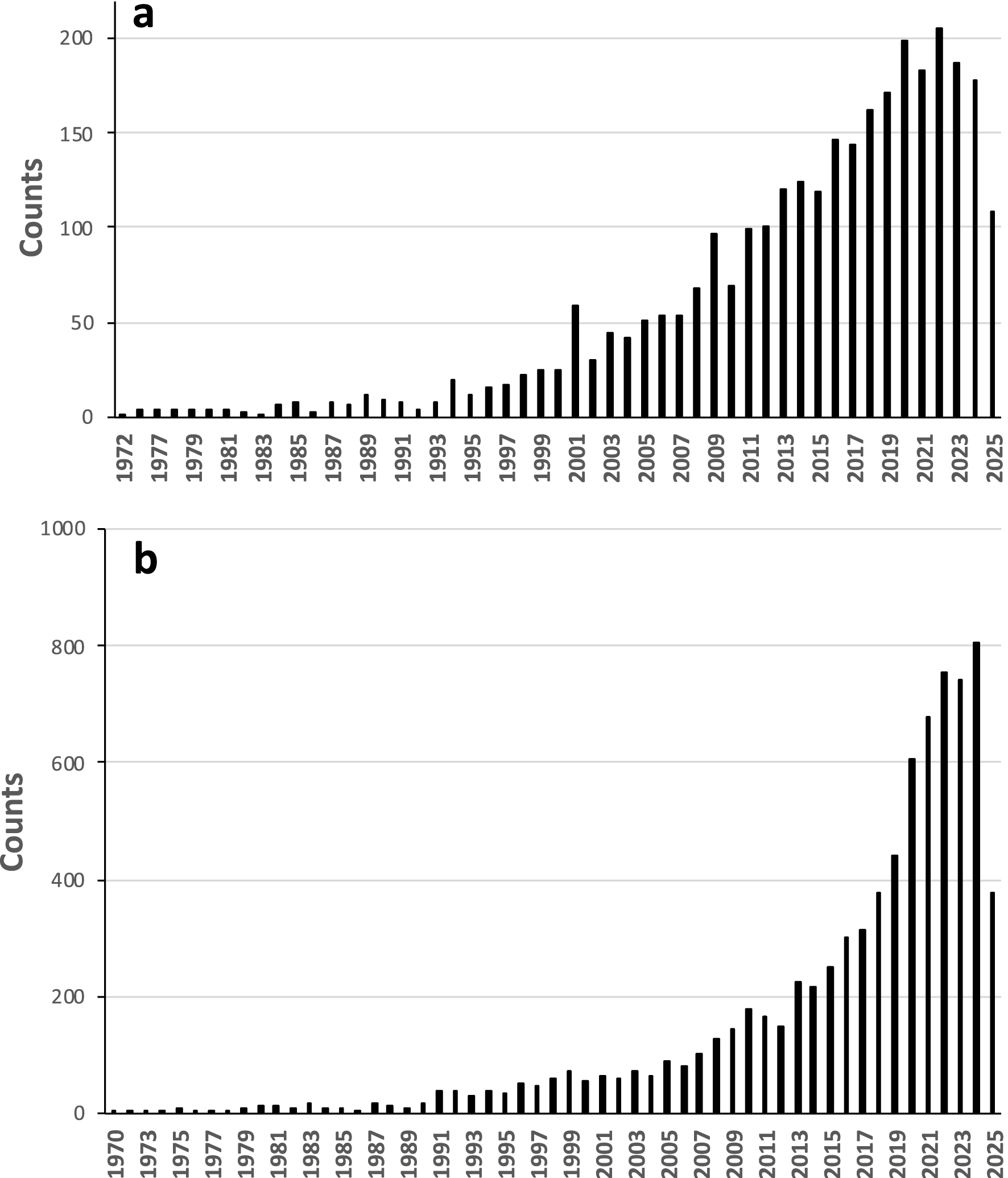
The objective of this review is to present the progress made in our understanding of the silicon cycle in ecosystems since Bartoli’s (1983) model was introduced and to highlight the significance of these findings for understanding the global silicon cycle and its potential implications for agricultural practices.
2. Drivers of Si mobility
2.1. Phytoliths as major players
At the core of Bartoli’s model (1983) is soil water, which contains dissolved silicon (DSi) (Figure 1). DSi results from the chemical weathering of silicates, a process driven by water and carbon dioxide that releases silicon into the soil solution, as well as into rivers and oceans. The concentration of DSi depends on the intrinsic properties of minerals (such as solubility) and environmental parameters (e.g., pH or temperature). In 1993, Lucas et al. (1993) changed the strictly abiotic view of soil formation in tropical latitudes, which considered a highly leached surface horizon rich in gibbsite, a more siliceous horizon below, and at the base, a smectitic accumulation horizon. The presence of kaolinite above the gibbsitic horizon could not be explained by this abiotic model; a model involving a Si contribution from plants that recombines with Al to form kaolinite then appeared more coherent. Other studies followed, showing the importance of plants in weathering balances. Alexandre et al. (1997) analyzed the phytoliths of a lateritic soil profile in the equatorial forest of the Congo and used Bartoli’s model (1983). They revealed two important points: the Si cycle was mainly controlled by plants, which reinforced the studies of Lucas and Bartoli, and some of the phytoliths (approximately 10%) were not rapidly dissolved and thus constituted a stable pool of amorphous silica in the soil. Meunier and Colin (1999) analyzed a soil rich in amorphous silica located on the volcanic island of Réunion (Indian Ocean). They demonstrated that this 15 cm thick layer was composed of Poaceae phytoliths. These phytoliths accumulated because of the convergence of 3 factors: soil developed on volcanic ash rich in Si, Si-accumulating plants (bamboo), and, finally, acidic soil pH, which is unfavorable for the rapid solubilization of amorphous silica. The Bartoli model was thus enriched with a new pool with stable phytoliths in soils.
However, very few studies on the physicochemical properties of phytoliths were available before the mid-1990s. It was therefore assumed that the phytoliths had the same properties as the other amorphous silica phases did. This hypothesis was the subject of the work of Fraysse et al. (2006) and Fraysse et al. (2009), who analyzed the solubility and dissolution rate of phytoliths in the laboratory. Their results confirmed that the solubility of phytoliths was identical to that of other forms of amorphous silica but that their dissolution rate was lower than that of vitreous silica. The dissolution rates increase with increasing pH and thus explain why phytoliths accumulated on Réunion (Meunier and Colin, 1999). Fraysse et al. (2009) reported that, beyond pH 4, the dissolution rates of phytoliths were higher than those of aluminosilicates, which explained why phytoliths can control the Si cycle in Bartoli’s model (1983), despite their low proportion in soils (generally less than 1 wt%).
By measuring germanium and silicon in stream waters in Hawaii, Derry et al. (2005) provided new evidence of the importance of phytoliths. Their results revealed that stream waters were distributed along a hyperbolic curve between a pole characterized by low dissolved Si (DSi) values and high Ge/Si values (clays) and a phytolith pole characterized by high DSi values and low Ge/Si ratios. They showed that contribution of phytoliths predominates in the stream waters. At the global scale, Conley (2002) evaluated the role of plant recycling by comparing the annual production of amorphous silica (ASi) in the form of phytoliths to that of diatoms in the ocean. The results revealed that the annual production of silica from phytoliths was between 60 and 200 Tmol⋅yr−1 and of the same order of magnitude as that of silica fixed in the oceanic biogeochemical cycle (240 Tmol⋅yr−1). Thus, for approximately thirty years, phytoliths have become major players in the Si cycle at the local and global scale.
2.2. Biological factors other than plants
Several studies have provided new insights into the role of living organisms other than plants (see review in de Tombeur, Rouw, et al., 2021). Sommer, Jochheim, et al. (2013) quantified the pool of testate amoebae in a German forest. Although the Si pool of testate amoebae tests was small, approximately 100 times less than that of phytoliths, the authors calculated an annual production of 17 kg of Si ha−1⋅yr−1 by testate amoebae, comparable to that of phytoliths. Consequently, testate amoebae can be considered a temporary pool of Si on a monthly scale, considering their relatively short generation times. However, the total protozoic Si pool and its dissolution rates remain poorly understood (Puppe, 2020).
Soil microfauna, such as earthworms and termites, can also play a role in the dynamics of Si. Bityutskii et al. (2016) analyzed water-extracted DSi in Russian earthworm casts and reported concentrations that were 2–12 times higher than those in surrounding soils. The authors suggested that passage through the earthworm digestive created finer particles and accelerated the dissolution of silicate minerals. Empirical indicators of plant-available-silicon (PAS) have been developed, which have been shown to be proportional to the silicon concentration in plants. The concept of PAS will be further discussed in Section 5. Jouquet et al. (2024) analyzed the role of earthworms in Vietnamese rice paddies with PAS measurements using 0.5 M acetic acid (SiAA). They reported, in agreement with the results of Bityutskii et al. (2016), an increase in the SiAA concentration in the casts. The authors also reported an increase in phytoliths in the casts, which could explain their higher SiAA values. Another explanation for the high SiAA values was proposed by Hu et al. (2018), who isolated silicate-solubilizing bacteria in the digestive tract of earthworms. These authors also inoculated these bacteria into pots filled with Chinese soils and reported that they had a positive effect on DSi and the amount of Si absorbed by maize.
The effects of termites on the PAS have also been documented (Albert et al., 2025). These authors reported higher SiCaCl2 (PAS using CaCl2 extractant) concentrations in non-fungus-growing termite mounds than in surrounding soils and a positive correlation between PAS and phytolith concentrations. This type of termite accumulates residues of fecal material rich in poorly degraded leaf and stem plant debris, which explains the richness of the phytoliths that control SiCaCl2.
Large animals can also impact the silicon cycle. Vandevenne, Barão, Schoelynck, et al. (2013) studied the impact of large herbivores on the solubility of phytoliths. They reported that compared with phytoliths extracted from hay, phytoliths from horse, cow, and sheep feces dissolve 3–5 times faster. Schoelynck et al. (2019) quantified the effect of hippopotamuses (in Kenya), which have the ability to ingest large quantities of food from the grassland savanna at night and rest during the day in the hippo pools of river meanders, where they evacuate their fecal matter. The authors were able to quantify the Si transferred from soils to rivers and reported that 76% of the total Si flux was affected by hippopotamuses. Large herbivores thus have a significant effect on the turnover of Si in soils and on the balance of Si delivery to aquatic systems.
2.3. Physicochemical factors
Since the 1960s, the adsorption of Si on the surfaces of soil minerals has been known to increase with increasing pH (Beckwith and Reeve, 1963). Laboratory analyses have shown that the amount of desorbed Si decreases over time (Haynes and Zhou, 2020), suggesting that over time, a fraction of adsorbed DSi becomes immobile. Although the mechanisms are not yet well understood, these results could lead to a decrease in DSi in alkaline soils.
The effect of aggregation on the solubilization of phytoliths was evaluated by Li et al. (2022) on mixtures of materials composed of organic matter, aluminosilicates, oxides, and phytoliths extracted from rice leaves. Their results revealed that aggregation can decrease the release of DSi from phytoliths by up to 3.9 times depending on the type of material used. Other factors can influence the dissolution of phytoliths, such as morphology (Cabanes et al., 2011), age (Liu et al., 2023), redox conditions (Koebernick et al., 2022), and fire (Cabanes et al., 2011).
2.4. Si isotopes as tracers of the sources of dissolved silicon (DSi)
In addition to the use of the Ge/Si ratio for tracing the source of DSi, the use of Si isotopes has been developed. Si has three stable isotopes: 28Si (92.23%), 29Si (4.67%), and 30Si (3.1%). Measurements of isotopic ratios are generally expressed in δ notation:
| \begin {eqnarray*} \updelta {}^{30}\mathrm {Si} &=& [(\mbox {}^{30}\mathrm {Si}/\mbox {}^{28}\mathrm {Si~sample})/ (\mbox {}^{30}\mathrm {Si}/\mbox {}^{28}\mathrm {Si~standard}) - 1] \times 1000 \end {eqnarray*} |
In nature, δ30Si values range from −5.7‰ for silcretes (Basile-Doelsch et al., 2005) to +6.1‰ for rice grains (Ding et al., 2005). Parent rocks, including crustal, magmatic, and sedimentary rocks, are characterized by values of approximately 0 (Frings et al., 2016). Fractionation in soils can be explained by water–rock interaction mechanisms or biological processes (Opfergelt and Delmelle, 2012). Thus, the Si isotopic composition of clay minerals formed during weathering is lower than that of parent minerals. In soils, low δ30Si values are indicators of strong weathering (Ziegler et al., 2005). Furthermore, this phenomenon is amplified by the adsorption of Si on secondary Fe minerals, which is accompanied by the preferential retention of light isotopes. Si uptake by plants is accompanied by an enrichment in light isotopes, and during the transfer to the shoots, heavy isotopes are preferentially incorporated into the phytoliths. Depending on their location in the plant, the δ30Si values of phytoliths range from −2.3 to +6.1‰ (Opfergelt and Delmelle, 2012). In an analysis of the different compartments of a tropical forest ecosystem in South India, Riotte et al. (2018) reported that the heavy δ30Si values of water infiltrating through the soil were similar to those of phytoliths, thus revealing that the latter controlled DSi. However, in acidic brown soils in France (Cornelis, Delvaux, et al., 2010), the results differed, with the light δ30Si values of forest floor leachates being controlled more by the dissolution of clays than by phytoliths. In the temperate Black Forest (Germany), Steinhoefer et al. (2017) reported that δ30Si values in streams were lower in spring than in late summer. During spring, the increase in dissolved organic carbon triggers a decrease in the pH and preferential dissolution of clays and a decrease in δ30Si values. During summer, the formation of secondary minerals and Si uptake by plants lead to positive δ30Si values. The use of silicon isotopes remains a promising tool for constraining the dynamics of silicon in soil–plant interactions.
3. Improving the quantification of Si pools and fluxes
Progress in quantifying plant–soil silicon transfers has been achieved through the publication of data on Si concentrations in plants, absorbed Si fluxes, and concentrations and fluxes of phytoliths in soils. Hodson et al. (2005) compiled data on 735 species from 125 publications and reported that, in general, ferns, gymnosperms, and angiosperms contain less Si than nonvascular plants and horsetails do. Furthermore, compared with other monocot clades, Poales and Arecales accumulate more Si in their shoots. The fluxes of absorbed Si, expressed in kg Si ha−1⋅yr−1, have been estimated in most terrestrial ecosystems, both natural and anthropogenic. In a bibliographic synthesis, Vander Linden and Delvaux (2019) reported a considerable range of values, between 0.7 and 1470 kg Si ha−1⋅yr−1. Using such analyses and the net primary production of terrestrial biomes, Carey and Fulweiler (2012a) reported that the annual amount of Si fixed depends on biome types and was higher in tropical woodland savanna and temperate steppe than in tropical forest, 22.19, 13.26, and 4.48 Tmol Si yr−1, respectively, thus highlighting the importance of grasses in the global Si cycle. The Si production of cultivated land reaches almost 30 Tmol Si yr−1, which is 35% of the production of all the biomes combined. The high proportion of cultivated land is explained by the fact that 8 of the 10 most cultivated crops in the world are Si-rich plants: sugarcane, maize, rice, wheat, soybeans, sugar beet, barley, and tomatoes (Guntzer, Keller and Meunier, 2012). Given the predictable increase in cultivated areas to feed future populations, Carey and Fulweiler (2012a) hypothesized an increase in the transfer of bioavailable Si to aquatic environments, with possible consequences for the development of diatoms.
The fluxes of absorbed Si depend not only on the types of plants but also on the types of soils and the soil weathering stage (Cornelis and Delvaux, 2016; Vander Linden and Delvaux, 2019). Phytolith pools have been measured in many ecosystems (Haynes, 2017b). Natural grassland ecosystems contain a higher reservoir of biogenic silica (BSi) than forest ecosystems, with values ranging from 20 to 690 T Si ha−1 (for soil depths between 0.2 and 1 m) and 0.6–17 T Si ha−1 (for soil depths between 0.2 and 0.85 m), respectively, which is in good agreement with the absorbed Si fluxes presented earlier.
Finally, the contribution of atmospheric silicon (Si) inputs to soils remains poorly documented. It is generally assumed that Si inputs are neglected due to their low values (Sommer, Kaczorek, et al., 2006). For instance in a temperate forest ecosystem, Turpault et al. (2018) reported values of dust and DSi respectively of 0.2 and 6 kg Si Ha−1⋅yr−1 while uptake by plant represented 95–157 kg⋅yr−1. Xi et al. (2022) reported values of approximately 1 kg Si ha−1⋅yr−1 in Chinese terrestrial ecosystems from 2010–2020. These values are approximately 200 times lower than the Si uptake by rice shoots (219 kg ha−1⋅yr−1). Therefore, the input of DSi from rainfall does not appear to be negligible, as it would take only 4–10 years to meet the needs of rice if the straw is exported. These results warrant further validation in other regions worldwide.
4. Impact of human activities
The question of the impact of anthropogenic activities, such as intensive farming, deforestation, or urbanization, on soil silicon (Si) dynamics has been the subject of innovative research since the 2000s. With respect to deforestation, Conley et al. (2008) documented the effects on dissolved silicon (DSi) in stream discharges at the Hubbard Brook experimental site (USA). They reported that DSi exports were greater in deforested areas where biomass was left in place. According to the authors, the excess DSi was due to the dissolution of excess phytoliths from the biomass in place and not to an increase in the chemical weathering of primary minerals. Deforestation followed by the establishment of crops is known to disrupt soils and promote erosion, but no work has focused on phytoliths. Struyf et al. (2010) analyzed the fluxes of DSi and amorphous silica (ASi) in waters from 51 watersheds in Belgium. Their conclusion differed from that of Conley et al. (2008), as they reported that exports of reactive silica, TSi = DSi + ASi, were higher in forested areas than in agricultural areas. The authors suggested that the decrease in TSi in cultivated areas was due to the long-term depletion of phytoliths in the soils, whereas the results of Conley et al. (ibid.) applied to the short term (<20 years). Several studies have sought to verify this hypothesis in the 2010s.
Clymans et al. (2011) measured ASi stocks along a gradient of soils from forestland–pasture–arable land in Sweden. ASi was estimated by destructive alkaline extraction, called ASia. The results revealed that the total ASia values were greater in forests. These results were confirmed in another study along a gradient of land use change in temperate zones in Belgium using continuous extraction measurements of Si and Al by NaOH (Vandevenne, Barão, Ronchi, et al., 2015). In natural environments, such as forests, the quantity of phytoliths decreases with depth, as is the case for carbon (Alexandre et al., 1997). In a cultivated field with biomass export, a decrease in the stock of phytoliths in the surface horizon was expected. This hypothesis was documented at Hubbard Brook (Saccone et al., 2008). These authors reported that after deforestation, ASi stocks in the O horizon decreased by 34%. At Rothamsted Research, Guntzer, Keller, Poulton, et al. (2012) documented a decrease in ASi in a cultivated field with partial or total export of straw after 1844. It has thus been clearly established that soil phytolith stocks can decrease when litterfall is disturbed by human interventions in the field.
Another aspect of the perturbations of the Si cycle by human activities has been evaluated: the role of urban systems. Indeed, natural cycles are strongly disrupted by urbanization, and this also concerns silicon, as several studies have shown. The export and transformation of plant materials can occur in urban areas and they can be released into rivers through wastewater (Vandevenne, Struyf, et al., 2011). Sferratore et al. (2006) analyzed the impact of urban discharge on the DSi balance in the Seine River (France). Their study revealed that the concentration of DSi in urban runoff was at least twice as high as that in rainwater, thus indicating that impervious urban surfaces contain alterable siliceous materials. Carey and Fulweiler (2012b) analyzed DSi exports in a gradient of watersheds ranging from forested to urbanized areas in the USA and reported lower values as forest cover increased. These authors also analyzed the effect of seasonality and reported a decline in DSi in forested areas between February and April, which was lacking in urban areas. The authors attributed this decline not to volumetric dilution or to the development of river diatoms but to trees preferentially absorbing DSi during this period. Maguire and Fulweiler (2016) specifically estimated DSi release in urban areas in Boston (USA) and reported that DSi release was greater at the outlets of wastewater treatment plants than in infiltration water and runoff. These results confirmed the importance of domestic consumption on DSi fluxes in urban areas.
5. Si in agroecosystems
The beneficial role of silicon in the growth of cereals has been known for more than a century (see review in Epstein, 1999). Over the past thirty years, a significant number of studies have documented the positive impact of Si not only on yields but also when the plant is subjected to biotic and abiotic stresses (de Mello Prado, 2023; Liang, Nikolic, et al., 2015). The mechanisms of these beneficial effects are still poorly understood, but since the work of Ma et al. (2006), it has been established that uptake can be active, meaning triggered by genes, in Si-accumulating plants (for instance, rice, barley, maize, wheat, and horsetail). Proteins from the aquaporin family are involved. The silicon accumulated in plants can also be explained by passive absorption controlled by water flux (transpiration). In cultivated plants such as rice and maize, both mechanisms coexist (Liang, Hua, et al., 2006).
The accumulation of Si in plants depends on the pool of bioavailable Si in the soils that supply DSi. However, measurements of DSi in solutions are rare and difficult to implement; thus, another approach is used, namely, chemical extractants as plant-available silicon indicators or PAS (Sauer et al., 2006). For example, Haysom and Chapman (1975) extracted Si from Australian soils using a 0.02 N CaCl2 solution and reported that the concentrations were proportional to the sugarcane yield. This method has since been confirmed on other soils and plants, such as Indian soils cultivated with rice (Narayanaswamy and Prakash, 2010). The CaCl2 method is used to characterize the easily soluble Si pool and is thus directly mobilizable by plants. Other more powerful extractants capable of desorbing Si, such as acetic acid (see the review in Tubana et al., 2016), have also been proposed. Measurements of DSi by different extractants are correlated with each other (Narayanaswamy and Prakash, 2010). The practical interest of these techniques is that they can be used to determine the critical thresholds of Si for a given cultivated plant. Below the critical threshold, an input of Si fertilizer to the soil would ensure optimal yield (Paye et al., 2018).
However, these PAS do not characterize the real pool of bioavailable Si, which remains poorly understood. By combining Si analyses with other soil properties (pH, particle size, amorphous phases, CEC, etc.), several studies have attempted to characterize the mineral phases of soils that control PAS. There is a positive correlation between the PAS and pH. In highly weathered acidic soils of South Africa and South India, the PAS are lower than those in soils close to neutrality, which are richer in alterable silicate minerals (Miles et al., 2014; Meunier, Sandhya, et al., 2018). Klotzbücher et al. (2018) also reported such a correlation in chernozems in Germany, suggesting the role of phytoliths and sorption as factors controlling PAS. Indeed, the solubility of phytoliths as well as adsorption increases with increasing pH. Two studies on more than 2000 soil samples in France (Landré et al., 2018; Caubet et al., 2020) helped to specify the role of pedological factors on PAS using the CaCl2 method (SiCaCl2). Landré et al. (2018) reported that SiCaCl2 values were negatively correlated with total Si concentrations when all soil types were combined (r = −0.32, p < 0.001). In regard to podzols, this negative correlation was stronger (r = −0.75, p < 0.001) and was explained by the presence of quartz, a mineral that is recalcitrant to dissolution. In contrast, the authors observed a slightly positive correlation between these same parameters (r = 0.12, p < 0.001) for carbonate soils, which are known to be poor in quartz.
Caubet et al. (2020) reported that SiCaCl2 concentrations were positively correlated with clay content (<2 μm fraction) and pH. Caubet et al. (ibid.) also reported higher SiCaCl2 concentrations in cultivated soils than in forest soils. This result can be explained by the liming commonly practiced in cultivated soils, which not only increases the soil pH but also soil PAS. To better explain these results, another study by the same team focused on characterizing Si pools in pairs of cultivated soils over two centuries and their neighboring forest soils (Cornu et al., 2022). The authors showed that allophanes (short-range order aluminosilicates), included in the <2 μm fraction, control PAS and not the phytoliths. The role of clays as a source of Si for plants was the subject of a pot study with wheat and rice (Keller et al., 2021). The authors reported that a soil consisting of 5% amorphous silica (in the form of diatomite) provided wheat with the same amount of Si as a soil consisting of 25% vermiculite would. The results showed that the identification of the source of DSi requires good characterization of the soil properties.
In regions of the world where soils are highly weathered and desilicified, phytoliths can constitute the main source of dissolved silica (DSi) (Cornelis and Delvaux, 2016; de Tombeur, Turner, et al., 2020). As previously mentioned, intensive cereal cultivation combined with straw removal can deplete soils of phytoliths. The same is true for PAS, as demonstrated by a study conducted in rice paddies in China (Yang, Song, Van Zwieten, et al., 2021). The authors compared soils sampled in the 1990s with those collected between 2016 and 2018 and reported an approximately 14% decrease in NaAC-extractable Si. They also reported that 66% of China’s paddy fields are deficient in PAS.
Since the 1980s, a decline in rice yields has been documented across Asia. This phenomenon could result from multiple factors, one of which is a deficiency in PAS (Savant et al., 1997). This hypothesis is supported by numerous experimental studies—conducted under hydroponic, pot, and field conditions—that have tested the effects of silicon fertilization on soils (Barao, 2022; Liang, Nikolic, et al., 2015; Haynes, 2017a; Tubana et al., 2016). Unlike nitrogen, the beneficial effects of silicon on crop plants are not systematic. A relevant example was provided by Sandhya et al. (2018), who conducted pot experiments using acidic, neutral, and alkaline soils from southern India under both field capacity and submergence conditions, with silicon fertilizer applied in the form of diatomaceous earth (DE). Under submerged conditions, DE application improved both Si uptake and grain yield in acidic soils. In contrast, for neutral soils, no significant effect on Si uptake was observed, and a yield increase was noted only at high application rates (600 kg/ha DE). Acidic soils, which are generally low in PAS, benefit from silicon inputs that improve yield. Conversely, neutral soils, which are richer in PAS, are less responsive. The observed yield increase without a corresponding increase in Si uptake may be due to the presence of other nutrients in the amendment. In alkaline soils, Si supplementation helps plants better cope with stress caused by alkalinity and salinity, thereby increasing rice growth (de Mello Prado, 2023). Furthermore, under field capacity conditions, Si addition did not significantly increase Si uptake or grain yield in acidic soils. This study underscores the importance of water availability in controlling the release of DSi from fertilizers, highlighting water as a key limiting factor for optimal Si uptake by plants. Many studies have also used materials derived from mineral extraction—such as the diatomaceous earth mentioned earlier—as well as industrial by products (Liang, Nikolic, et al., 2015). The effectiveness of Si fertilization on crop yields therefore requires a thorough understanding of the soil’s physicochemical and agronomic parameters.
Another strategy to increase the soil PAS concentration is to retain crop residues (straw) onsite. This hypothesis was tested in a long-term field experiment in Germany (Puppe et al., 2021). The authors compared the PAS levels as SiCaCl2 in soils with and without straw application from 1976 to 2018. The results revealed a significant increase in SiCaCl2, as well as an accumulation of soil phytoliths, over time. Another example of long-term analysis was provided by a 36-year study in China (Yang, Song, Qin, et al., 2020), where the authors showed that the so-called “labile” fraction of Si—measured as the sum of extractions using CaCl2, acetic acid, H2O2, oxalate, and Na2CO3—was higher in fields that received phytolith-rich straw. Additional evidence of the positive effects of plant-derived Si inputs on PAS was provided by Song et al. (2014), who used Si-rich manure at experimental sites in China. After five years of application, a significant increase in Si extracted using the acid-Na acetate method was observed down to a depth of 30 cm.
Although the direct application of silicon fertilizers has proven effective at improving crop yields, its use remains limited in agricultural practices. This can be attributed to several factors, including insufficient knowledge about the physiological role of Si and a lack of local data on PAS levels in soils (Zellner et al., 2021).
6. Conclusions
The continental biogeochemical cycle of silicon has become more complex since the work of Bartoli (1983) (Figure 3). Phytoliths, which reflect the recycling of soil silicon by plants, are now considered major drivers of this cycle, as they dissolve more rapidly than most other aluminosilicate minerals in soils do.
A modern picture of the biogeochemical cycle of Si in natural and cultivated terrestrial ecosystems (modified from Meunier, Cornu, et al., 2022).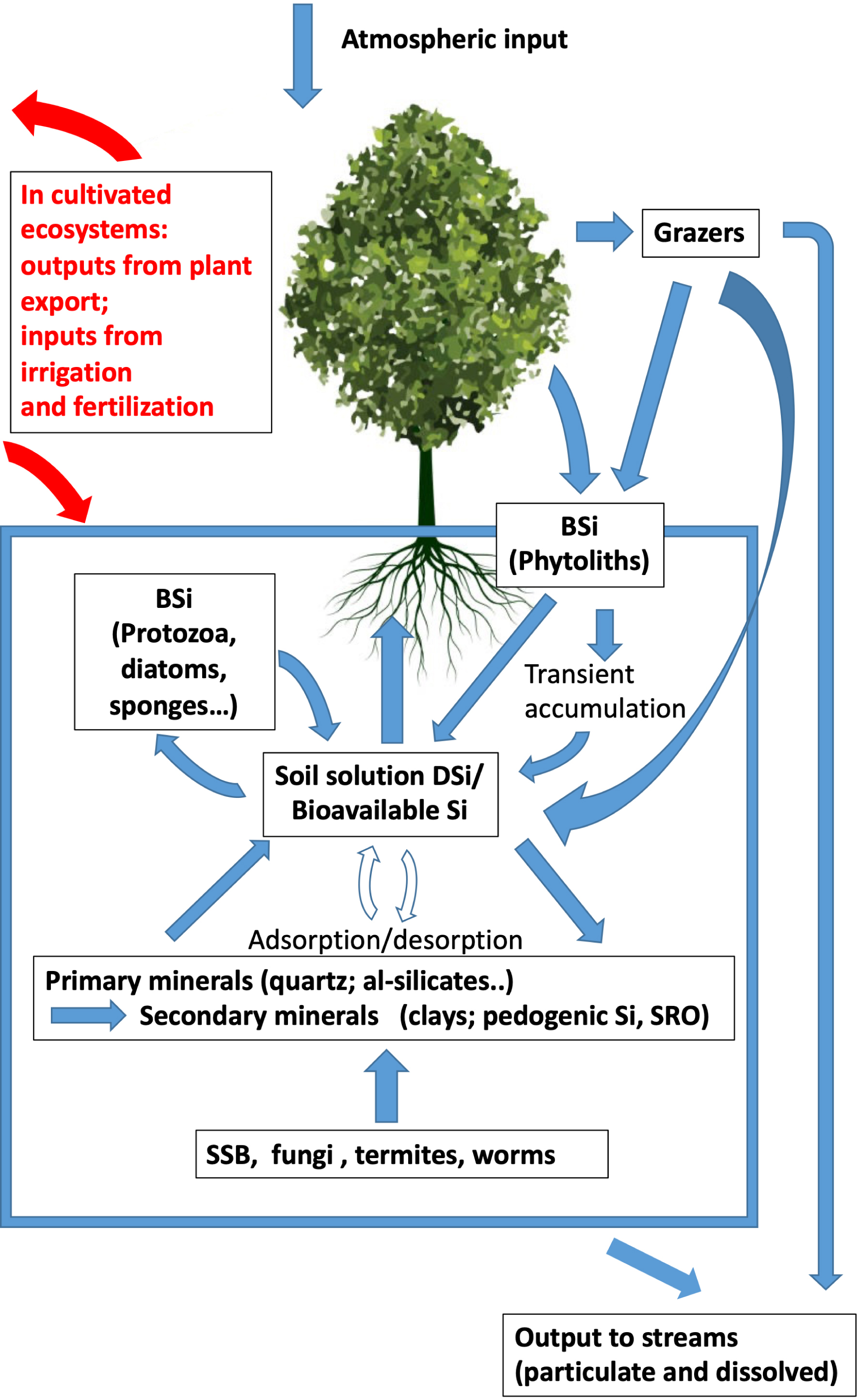
In addition to plants, other biological factors have been shown to influence the cycle, ranging from microorganisms (e.g., testate amoebae) to large herbivores and soil engineers. Human activities disrupt the cycle, with the most well-documented impact being the decline in phytoliths in the surface horizons of soils.
The hypothesis that silicon may be a limiting nutrient for crop growth has spurred research on the factors governing the mobilization of silicon in solution. Indicators of bioavailable silicon (PAS) have become more widespread, with practical applications in agronomy, as threshold values are now available.
The use of silicon isotopes and Ge/Si ratios makes it possible to constrain the silicon cycle at the ecosystem level, even though the results of such studies are not always straightforward.
To predict the appropriate doses of silicon fertilizers to apply to soils to improve crop yields or to forecast changes in the global silicon cycle, it would be desirable to develop mechanistic models that consider the multiple factors that have been documented in recent years.
Acknowledgements
I would like to thank the anonymous reviewers whose comments and suggestions helped improve both the content and the form of the manuscript.
Declaration of interests
The author does not work for, advise, own shares in, or receive funds from any organization that could benefit from this article, and has declared no affiliations other than their research organization.