


 CC-BY 4.0
CC-BY 4.0
1. Introduction
Science writer Fen Montaigne calls it “one of the greatest mysteries of our time… when humans made the first bold journey to the Americas” [Montaigne 2020]. The solution was once thought certain but now profoundly is not. The when, where, and how of this crucial migration continue to be hotly debated in archeology, anthropology, oceanography, and human geography. According to present consensus, any viable hypothesis must meet four criteria: (1) a source population in Asia at the start, (2) a reasonable pathway with sufficient sustenance for human survival, (3) documented settlement sites in North America soon after but not before the proposed date, and (4) a sanctuary where migrants could have sojourned long enough to develop a distinctive genetic profile with little sustained input from Asian forebears.
From the 1930s onward, the Clovis-first [Archaeological Notes 1932; Haynes and Huckell 2007] via interior Ice-free Corridor [Johnston 1933; Antevs 1935] hypotheses were believed to meet the first three conditions. Starting in 2007, however, researchers found pre-Clovis settlement sites in North America that challenged Clovis-first and its reliance on the deglaciation corridor [Waters and Stafford 2007]. Later, investigators questioned when the deglaciation corridor could have provided sufficient nutrition for human migrants and determined a date 300 years after the youngest Clovis sites [Pedersen et al. 2016]. Subsequently, the Kelp Highway hypothesis posited a maritime route sustained by coastal resources and nearshore kelp forests [Erlandson et al. 2007]. Then, Dobson et al. [2020] retrospectively mapped an ancient transitory archipelago that offers a third alternative: an island-rich maritime route like stepping stones across the Bering Sea. Scores of scattered, continuously evolving islands were available to maritime people at least as early as 30,000 BP to as late as 8000 BP. This route would have provided abundant freshwater; rich sources of food from vastly expanded shorelines, kelp forests, fisheries, and game; fuel from driftwood, island shrubs, and forests; protected waters for people foraging and traveling in boats; and frequent haul-outs. Key dates of inundation, which would have forced sequential evacuation of island populations, are consistent with early evidence of humans settling in North America. Our reconstructions are based on the solution of a topographically and gravitationally self-consistent sea-level equation that accounts for deformation and gravity variations in response to glacial melting. Details are given in our supplementary materials.
This paper builds on three previous ones that establish the global context of Beringia and reveal spatiotemporal opportunities for human crossings. First, Dobson [2014] emphasized the importance of aquaterra as a global feature and gave it a name. Aquaterra is the collective identity of all lands that were inundated and exposed repeatedly during the Late Pleistocene ice ages from the first appearance of modern humans through today. Second, Spada and Galassi [2017] refined the boundaries of aquaterra at coarse resolution for the whole earth and finer resolution for Doggerland and Beringia, using a Glacial Isostatic Adjustment (GIA) model. Third, Dobson et al. [2020] conducted GIA mapping and hypsometric analyses of nine global choke points—Bering Strait, Isthmus of Panama, Bosporus and Dardanelles, Strait of Gibraltar, Straits of Sicily and Messina, Isthmus of Suez, Bab al Mandab, Strait of Hormuz, and Strait of Malacca—to assess their status at the Last Glacial Maximum (LGM) ∼20,000 years ago. In the course of that global research, our paleotopographic reconstructions digitally discovered previously unnamed islands in a broad swath across the Bering Sea. The authors named them the Bering Transitory Archipelago and noted their relevance to the longstanding debate over when, where, and how the first humans crossed from Asia to North America. Here we further investigate the pathway and its implications for existing theories.
2. The Bering Transitory Archipelago
The sequence of island formation and inundation in the Bering Sea is captured in accompanying maps (Figures 1 and 2) selected from a series of 61 maps representing model results for each 500-year time step from 30,000 BP to today. Our GIA model and input data are described in supplementary material available online. The full animation of all time steps is available in our supplementary materials. Readers are encouraged to view the animation itself, which vividly portrays sea-level rise and fall with associated land changes, and read the verbal description accompanying it.
Reconstructed map of Beringia today and at 10,000 BP (soon after the Bering Strait reopened) Albers equal area conic projection.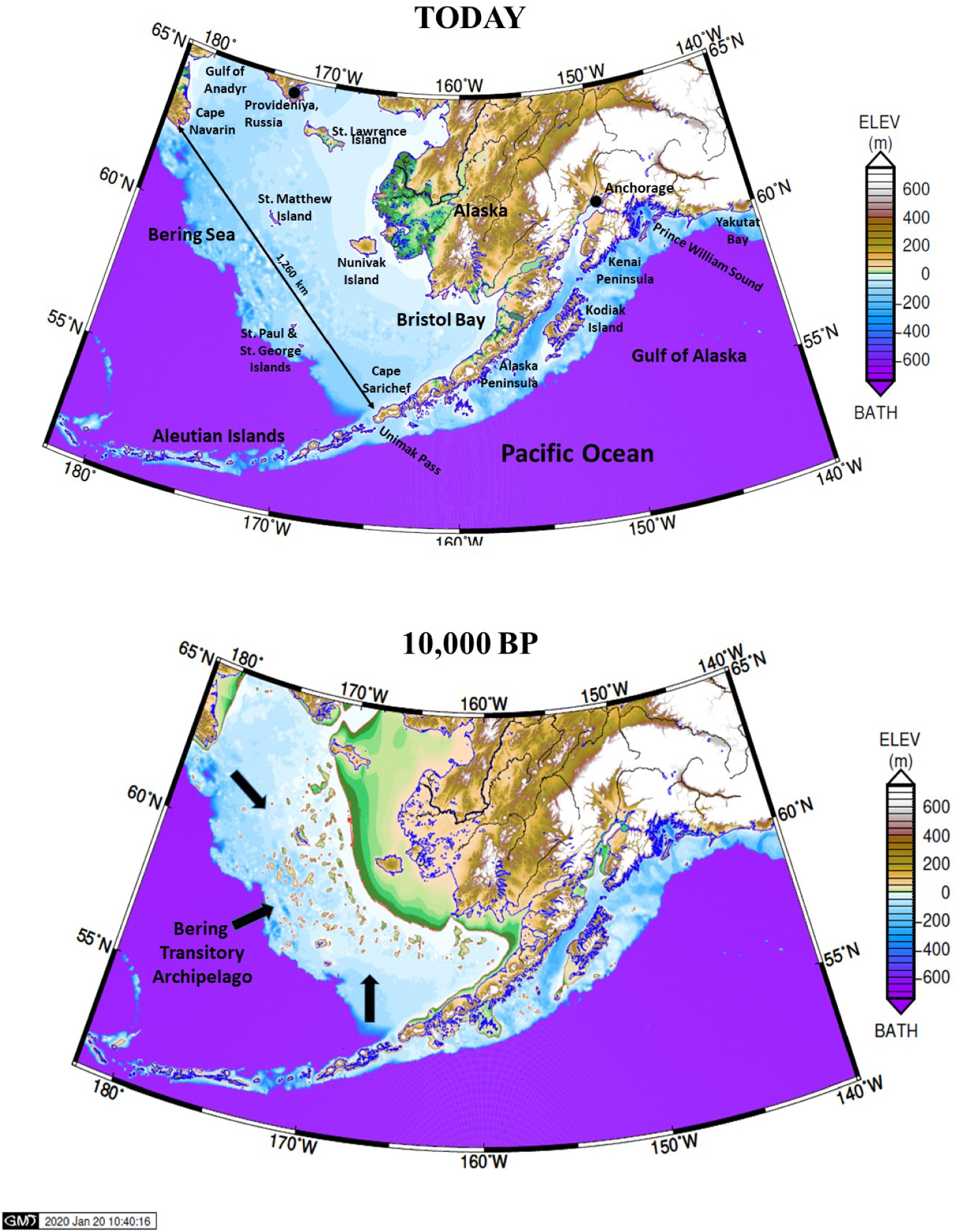
Glacial Isostatic Adjustment (GIA) reconstructions of Beringia at 30,000 BP (as early as ice history data go), 20,500 BP (Last Glacial Maximum), 10,500 BP (just before the Bering Strait opened), and 8000 BP (shortly before inundation was complete). Potential kelp habitat is highlighted in bright red at depths of 3 to 20 m which are suitable for bull kelp wherever rocky bottom may occur. Albers equal area conic projection.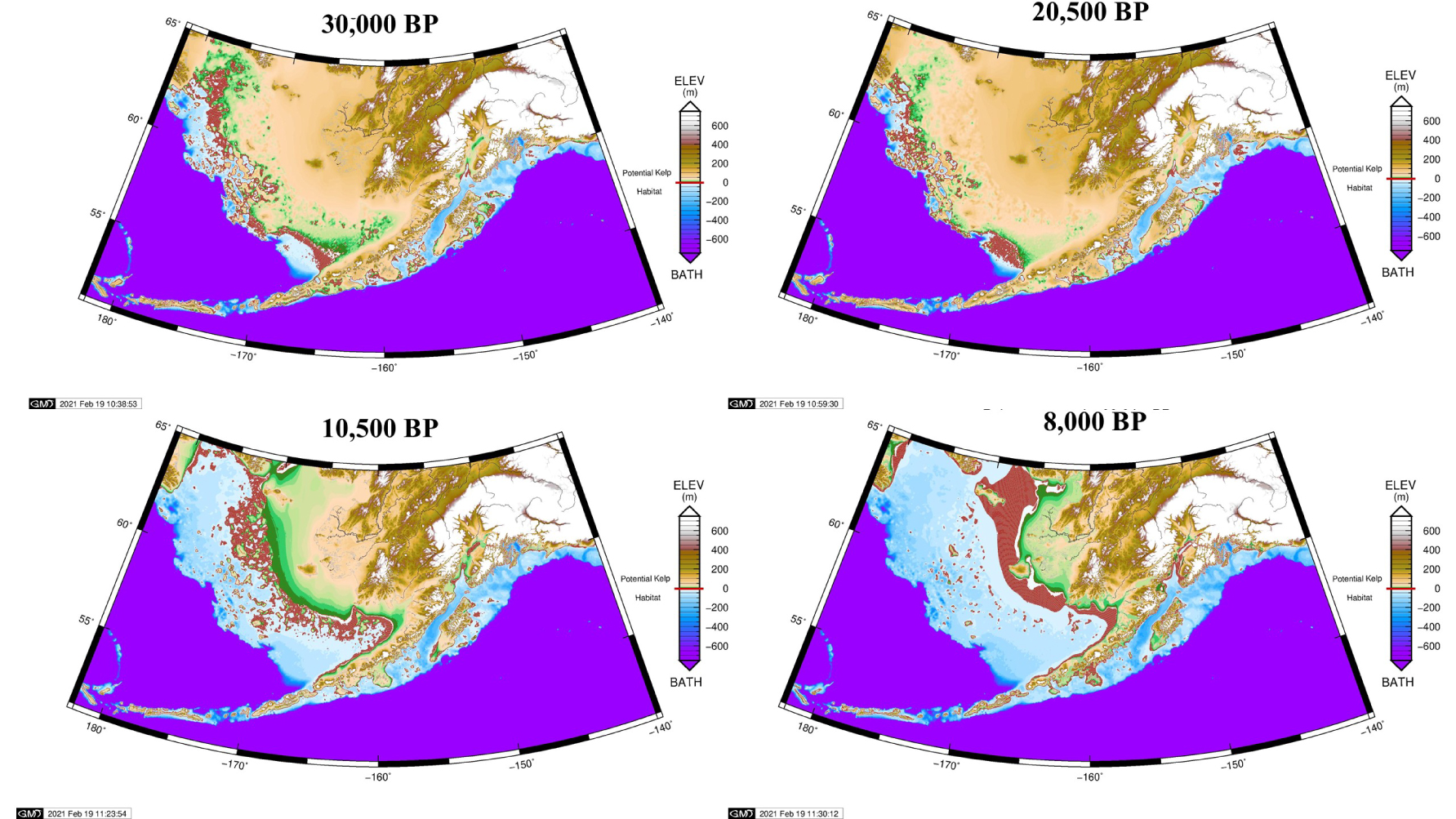
The first digital “sighting” of scattered islands in the ice-age Bering Sea appeared in a geovisualization of sea-level change from 21,000 BP to 2002 CE rendered by William F. Manley of the Institute of Arctic and Alpine Research (INSTAAR) in 2002 [Manley 2002]. A video of maps at 1000-year intervals was posted on the author’s institutional website, but the simulation was not documented in refereed literature. Soon afterward, Brigham-Grette et al. [2004] produced grayscale images of “Schematic sea level reconstructions at modern, −54 m, −64 m, −77 m, −88 m, and −120 m based on Manley [2002] without compensation for postglacial sedimentation or tectonic influences.” They observed (p. 36), “Detailed examination of [Manley’s] schematic paleogeography maps … implies that the southern shore of the Bering land bridge was geomorphically complex, with hundreds of islands located just off a coast riddled with bays and inlets. Such a coastline may have been a rich marine habitat for walrus and seals, both as haul-out spots and breeding localities. The possible abundance of such habitats has clear implications for the coastal migration theory.”
Dixon [2013] published grayscale images of Manley’s video in a peer-reviewed, 321-page book published by the US National Park Service. Brigham-Grette’s islands appear to be fewer and larger than Manley’s and Dixon’s. We are not able to verify individual island matches between ours and theirs due to differences in resolution and documentation. Later, Dixon and Monteleone [2014] referred to the “southern Beringian archipelago” (in lower case) and astutely observed, “…this long-lived geographic feature possibly offered a complex system of coves, bays, and intertidal features that may have been biotically productive and provided maritime subsistence resources and sheltered channels suitable for coastal navigation.” That same article, however, introduced a bibliographic error that muddles the precedence of discovery even today. Dixon [2013] correctly cited Manley’s video, but Dixon and Monteleone [2014] accidentally transposed the end of one entry (journal, volume, number, pages) with the beginning of another (author, year, title) which made it appear that Manley’s finding was documented in Science. We were the first to catch this bibliographic error, and this paragraph is the first public notice of it.
Neither Manley, Brigham-Grette, nor Dixon proposed a name for the archipelago or islands within it. We are the first to produce GIA reconstructions of the Bering Transitory Archipelago.
In Finkl’s [Finkl 2004], standardized classification of all coastal landform types the Bering Transitory Archipelago’s geomorphology and causality fit within a category of “lithospheric modulation through isostatic and eustatic movements tens of thousands of years ago (Type 4. a. & b. Time Sequence Order 2. c.).”
3. Transitory islands in the Gulf of Alaska
After clearing Unimak Pass, ice-age travelers faced arduous voyages eastward to the nearest ice-free haul-outs with access to lands beyond the ice sheets. From 30,000 BP to 11,000 BP, there were closely spaced islands in the Gulf of Alaska as far east as Middleton Island (146 W), then a gap of about 200 km that would have to be navigated alongside the present coast, next a pair of islands close together, and then another 200 km gap with a stretch alongside today’s coast as far as Yakutat Bay. Ideally this study should continue southeast to the Columbia River, but our mapping stops at 140° W due to practical limits on how much territory we can present cartographically with spatial resolution adequate for analysis.
A major test lies in those 200 km gaps. Royer and Finney say paddlers in skin boats could manage 24 to 36 hours between haul-outs [Royer and Finney 2020]. Hence, when transitory islands were exposed, paddlers would need to average 5.6 to 8.3 kph in order to cross each gap. Royer believes a voyage from Middleton Island to Yakutat Bay would have been feasible. From 24,000 BP to 18,000 BP, conditions would have favored boat journeys to the Pacific Northwest while the opposing Alaska Coastal Current (ACC) was suppressed. In the Bølling–Allerød warming period (15,000 BP to 14,000 BP), the ACC may have hindered travel along the coast of Alaska and British Columbia sufficiently to cause a migration hiatus until the onset of the Younger Dryas cold period (12,800 BP to 11,600 BP) [Royer and Finney 2020]. The Stepping-Stones hypothesis may serve well but with variable rates of advance from fast to slow to halted or nearly halted travel.
4. The Clovis-first via Ice-free (deglaciation) Corridor hypotheses
The Clovis culture was discovered through excavations near Roswell, New Mexico–Burnet Cave, Blackwater Draw, and Clovis—from 1932 to 1937 [Archaeological Notes 1932; Boldurian and Cotter 1999; Haynes and Huckell 2007; Boldurian 2017]. Almost simultaneously, a deglaciation corridor was found to have existed from Beringia to the southern terminus of the North American ice sheet [Johnston 1933; Antevs 1935]. Dates for the Clovis culture—13,250 to 12,800 BP—[Waters and Stafford 2007] and the deglaciation corridor—initially estimated as 20,000 to 15,000 BP [Antevs 1935] and later revised to about 13,000 BP [Jackson Jr and Duk-Rodkin 1996] coincided in a window lasting at least 200 years. Thus, most researchers assumed the first Americans crossed from Asia to Alaska when Beringia was dry land and a deglaciation corridor allowed overland travel. For at least half a century, the Clovis-first and Ice-free (deglaciation) Corridor hypotheses were a matched pair.
Then, pre-Clovis sites in the Americas, many of them previously proposed, became widely accepted. Initially they were only 550 to 1250 years earlier, including a pre-Clovis mastodon hunting site in Washington (13,800 BP) [Waters et al. 2011], Paisley Caves in Oregon (14,500 BP) [Gilbert et al. 2008], and Monte Verde (14,220 to 13,980 BP) in Chile [Dillehay et al. 2008]. Genetic-based estimates suggest that an isolated population moved south of the North American ice sheets sometime after ∼19,500 BP [Llamas et al. 2016; Davis et al. 2019; Pinotti et al. 2019], about 6000 years before Clovis appeared. Now, approximately 1900 stone artifacts excavated in Chiquihuite cave in northwest Mexico suggest human presence 26,000 years ago and perhaps as early as 33,000 to 31,000 years ago [Ardelean et al. 2020].
Ultimately, Pedersen et al. [2016] concluded that the deglaciation corridor would not have been biologically viable for human survival until about 12,600 BP, approximately 200 years after the Clovis culture ended. Ultimately the Clovis-first via Ice-free Corridor hypotheses were no longer viable.
5. The Kelp Highway hypothesis
For more than a century, scientists debated the likelihood of a coastal migration route crossing the southern fringe of Beringia [Mason 1896; Fladmark 1979]. From the 1890s to 1935, that question stood alone because the deglaciation corridor had not yet been discovered. The challenge for advocates was to prove that beaches and nearshore waters could have provided adequate shelter and sufficient resources to ensure survival. Once the deglaciation corridor was widely accepted, coastal routes drew little scientific interest except for the occasional Clovis-first via deglaciation corridor skeptic [Fladmark 1979].
After distinctly pre-Clovis sites were finally accepted, attention returned to the coasts. The first proposal to address the new timing was the Kelp Highway hypothesis [Erlandson et al. 2007]. The notion of a “kelp highway” captured public attention, but proponents understood it to be far more than just seaweed [Braje et al. 2019]. Diversity and complexity characterize north Pacific Rim ecosystems with kelp playing an important role in shallow seas. This seaborne mass of flora and fauna bolstered the larder envisioned for travel on or near the coast as well as for subsistence economies wherever they settled at the ends of their journeys [Moss 2011]. The concept encompasses littoral and nearshore biomes, some not associated with kelp. There is also a possibility that Pleistocene terrestrial fauna may have persisted on some islands for quite some time after they were severed from mainland Beringia. Mid-Holocene age mammoth remains have been found and dated on Wrangel and St Paul Islands [Dixon and Monteleone 2014].
It is quite possible that a multi-island feature as vast as the Bering Transitory Archipelago could have hosted several diverse ecosystems. It is not yet feasible to discern precisely what distinctive patterns of flora and fauna existed where, but a landmark symposium and subsequent anthology advanced the field of study from 1979 onward [Hopkins et al. 2013]. Kelp forests exhibit a quick ecological response, appearing after favorable conditions, and disappearing after unfavorable ones. So, they may have represented an available food source during most of the period considered here, while colonization by other kinds of flora is difficult to speculate. Kelp likely was an important food source but certainly not the only one.
The Kelp Highway itself is a coastal route, envisioned with plentiful food supply due to nearshore kelp forests and estuaries which are rich in shellfish, fish, marine mammals, birds, and seaweeds that substantially augment terrestrial resources onshore. We hypothesize that the Bering Transitory Archipelago would have improved geographical access to kelp forest and other marine ecosystems, ensuring abundant nutrition while broadening the travel corridor itself. The presence of numerous low islands along Beringia’s southern coast—like the convoluted archipelagos of Southeast Alaska and British Columbia today—magnify the linear distance and productivity of coastline and nearshore habitats for maritime foragers, as well as provide protected and semi-protected waters for people in boats.
Kelp thrives today in a vast swath extending across the North Pacific Ocean from Northern China to Baja California [Erlandson et al. 2007] and other oceans in both hemispheres. Kelp forests occupy an ecological niche consisting of cool ocean waters near shore. Like coral, kelp must have a solid substrate on which to attach its anchors, called holdfasts; otherwise, it cannot live. Like terrestrial forests, kelp forests have multiple canopies hosting diverse species—invertebrates, fish, marine mammals, and even birds. Like mangrove forests, kelp forests serve as refugia for fauna that attach themselves to or hide beneath blades (leaves) for nourishment and protection.
Kelp forests are among the most productive biomes on earth [Rodgers and Shears 2016]. Bull kelp (Nereocystis luetkeana), ranging as far north as the Aleutian Islands, grows to a maximum of 36 m long with multiple blades up to 3 m long. They grow off rocky coasts in nutrient-rich tidal and intertidal waters to 20 m depth. Blades often reach the surface and then flow outward horizontally.
The Kelp Highway is not a static feature. It moves seaward as sea-level falls and landward as sea-level rises. Potential bull kelp habitat is mapped here, based on a depth range of 3 to 20 m but without geologic data to ascertain where the rocky bottom would support growth. Four maps are selected from a series of 61 maps representing model results for each 500-year time step from 30,000 BP to today (Figure 2). The full animation is available in our supplementary materials. From 30,000 BP to 14,500 BP, a smooth arc ran directly from Cape Navarin to Cape Sarichef. Subsequently, it diverted northward from both capes, grew increasingly sinuous, and eventually concentrated where it is today with little change after 6000 BP.
Henceforth, we will speak not of “kelp forests” but of nearshore biomes associated with coasts, islands, and shallow seas. Clearly, these geographical features satisfactorily address the longstanding concern about adequate nourishment for early migrants traveling on or near the coast. Yet, a major question remains: where on the route could travelers have stalled for thousands of years without frequent genetic input from Asia?
6. The Beringian Standstill hypothesis
The Beringian Standstill hypothesis [Hulten 1972; Tamm et al. 2007], also known as the Beringian Incubation Model (BIM), addresses the crucial question of migration pace regardless of route or arrival date. Did migrants trek quickly and directly across Beringia, as the term “land bridge” might imply, or was there a long sojourn in Beringia? A strong argument for the standstill lies in genetic differences between the first Americans and their Asian forebears [Tamm et al. 2007].
One challenge for advocates of the Beringian Standstill hypothesis is to find a time and place where migrants could have remained isolated from Asian populations long enough to develop a separate and distinctive genetic heritage. Moreno-Mayar et al. [2018] “infer that the Ancient Beringian population and ancestors of other Native Americans descended from a single founding population that initially split from East Asians around 36 ± 1.5 ka, with gene flow persisting until around 25 ± 1.1 ka.” Raghavan et al. [2015] favor “an upper bound of ∼23 ka on the initial divergence of ancestral Native Americans from their East Asian ancestors, followed by a short isolation period of no more than ∼8000 years.” Tamm et al. [2007] agree with the Raghavan et al. termination date (15,000 BP) based on a longer isolation period, “the initial founders of the Americas emerged from a single source ancestral population that evolved in isolation, likely in Beringia… The finding that humans were present at the Yana Rhinoceros Horn Site [in Siberia dated 30,000 BP] suggests that the isolation in Beringia might have lasted up to 15,000 years.”
John Hoffecker asks [Montaigne 2020, 40], “Where do you put these people where they cannot be exchanging genes with the rest of the Northeast Asian population?” Hoffecker et al. [2016] add, “Wherever it was located, the genetic diversity of the standstill population suggests that it may have been distributed across a large area and perhaps even subdivided among several regions.” They proposed that such isolation might be maintained by geographic patterns of varying biological productivity across mainland Beringia. Similarly, Sikora et al. [2019] propose that climatic factors may have caused abandonment in Siberia and created a refugium across southern Beringia. We do not dispute either possibility. However, when it comes to isolation over large areas and long time periods, nothing beats islands scattered across oceans. Indeed, the internally connected yet outwardly isolated maritime culture of Polynesia from 6000 BCE to 1595 CE offers a striking parallel to the pattern of isolation proposed above. The Bering Transitory Archipelago offers a previously unsuspected refugium with adequate land area, resources, and separation, which offers the tantalizing possibility of even earlier populations closer to the early split proposed by Moreno-Mayar et al. [2018].
7. Stepping-Stones hypothesis
Retrospective sea-level modeling affirmed a third, stunning alternative when Dobson et al. [2020] documented scores of islands that could have served as stepping stones for maritime travelers. This archipelago would have been an ideal staging area for LGM travelers heading east. Maritime travelers would have needed food, shelter, and maritime facilities such as fish traps and haul-outs for umiak and kayak. Its islands would have provided a maritime homeland in which to hone the precise skills needed—kayaking, hunting, sheltering, and surviving—to reach and settle North American landfalls. Its shores would have abounded in iodine, Docosahexaenoic acid (DHA), and other nutrients essential for human health and brain development.
This transitory, aquaterran archipelago existed intact from at least 30,000 BP to 20,000 BP. Soon afterward, a rapid inundation of the western islands began. The loss of islands after 20,000 BP matches the population movement genetically detected in North America by Llamas et al. [2016], Davis et al. [2019], and Pinotti et al. [2019]. From 10,000 BP to 8000 BP, the number and size of islands in the east reduced rapidly from (in 500-year time steps) fifty-seven to fifty-three to thirty-nine to seventeen then six, after which it remained steady at only a few. As these inundations occurred first in the west and then in the east, residents would have had no choice but to evacuate, however dangerously, choosing among Asia, the Aleutian Islands, and North America. The archipelago’s eastern shift after 10,000 BP would have favored migrating to North America by the seaward route. We urge underwater archaeologists to search for landfall settlements at depths of −90 m to present sea level off the coast of Alaska [Dobson et al. 2020].
In essence, the Stepping-Stones hypothesis visualizes a gigantic conveyor belt drawing migrants from Asia, gradually nudging them eastward, and forcefully dumping them wherever settlement sites are found from Bristol Bay to Yakutat Bay. A source area could have been on the Yana River of Siberia, where coastal hunters are known to have existed 27,000 years ago [Pitulko et al. 2004], or offshore islands in northern Japan where maritime people are known to have exploited obsidian deposits by 30,000 BP [Izuho and Sato 2007]. The “conveyor belt” would have lured coastal people from Siberia or maritime people arriving from farther south. In the beginning it would have facilitated sea travel and island settlement in the west. Soon after LGM and more dramatically at 16,000 BP, mainland Beringia receded from the westernmost islands, and the remaining islands became more isolated from shore. At about 16,000 BP, island densities began to decline in the west. At 15,000 BP, they were quite scattered. By 10,000 BP, when the Bering Strait finally opened, only three tiny islands remained west of the new strait, and the eastern islands grew profoundly isolated from Asian source areas. For more on island distributions and sea distances, see our supplementary materials.
Surveying all options for crossing from Asia to the Americas, Mason [1896, 277] speculated, “The greater part of the earth’s surface was sterile and repellent to primitive man or to the living forms upon which he depended.” He named six zones he thought would have been impassable from that standpoint; his first was “The deep sea, out of sight of coastal plains and meadows.” He was, of course, unaware of the Bering Transitory Archipelago, which provides islands and nearshore biomes in more abundance than anyone imagined, a path across the sea, and a comfortable place to remain isolated for thousands of years.
A stepping stone is “a stone on which to step (as in crossing a stream)” or “a means of progress or advancement.” [Merriam-Webster 2004] Both meanings are fitting analogies for the dual roles the Bering Transitory Archipelago potentially played in the Beringian Standstill and the peopling of the Americas.
The Stepping-Stones hypothesis can be tested only by reconstructing palaeolandscapes and conducting archaeological surveys. Resurgence in this type of research around the world (Europe, North America, Arabia, Australia, and South Africa) makes such vertically integrated investigations a realistic prospect [Bailey et al. 2017]. Duncan McLaren and colleagues advocate a stepwise process: (1) create localized sea-level curves, (2) generate detailed bare earth digital elevation models, (3) create archaeological predictive models, (4) verify the models using archaeological prospection, and (5) demonstrate that archaeological materials found date to the late Pleistocene [McLaren et al. 2020]. In British Colombia, for example, they mapped a sea floor, identified a submerged paleo-lake basin at −50 m, selected a promising target for archaeological survey, deployed a dredge, and retrieved indisputable worked stone and wooden artifacts.
8. Seaworthy craft, ice history, and migration
Three additional criteria are necessary for the Stepping-Stones pathway to work: seaworthy craft, an unimpeded maritime corridor, and accessible landing sites on the North American shore. We reviewed literature for the earliest evidence of seaworthy craft globally [Dobson et al. 2020]. Royer and Finney [2020] focused on the Bering crossing, and concluded that skin boats propelled by paddles likely would have been available. They say and we concur that “Early migrants to the Americas were likely seaworthy.” They further conclude, “Any migration during the Last Glacial Maximum and early deglaciation period may have been facilitated by altered coastal currents, which would have favored such travel for only a few millennia.”
How might ice sheets have impacted migration and landing? According to a model progressively developed at the Australian National University by Kurt Lambeck and collaborators [Nakada and Lambeck 1987; Lambeck et al. 2003] and associated ice-history maps which underlie our paleotopographic reconstruction, the area was free of thick ice sheets for 30,000 years. However, the southern coast of Alaska including the Alaska Peninsula was ice capped from 27,500 BP through 19,500 BP. Before 27,500 BP, the coast of Alaska was unimpeded by ice from Yakutat Bay westward. From 19,000 BP to the present, the bulk of Alaska’s southern coast was accessible. See “Ice History” in our supplementary materials.
The pattern of ice coverage on the Alaska Peninsula from 27,500 through 19,500 BP is about the same as Peltier’s map [Peltier et al. 2015], meaning the worst case is that migrants would have to navigate around Cape Sarichef through Unimak Pass to reach a passage unimpeded by ice before turning east again. The difference is that Lambeck shows no blockage after 19,500, while Peltier shows the blockage continuing until 11,000 BP. Taken together, these three conditions favor human coastal dispersals from 30,000 to ∼15,000 BP and after 12,800 BP. See our supplementary material.
Seasonal and perennial sea ice are quite another matter. As we said earlier [Dobson et al. 2020], “These islands, sounds, and bays would have been the active interface among ice, land, and ocean. [At times] The entire archipelago was backed on the north by a broad swath of tundra [Ray and Adams 2001], meaning the area likely would have been as habitable as the north shore of Alaska today.” Brigham-Grette et al. [2004] state, “sea ice was present on the margins of Beringia for nine months or more each year during the LGM, making coastal migration from northeast Asia difficult, if not impossible. This barrier likely did not disappear until after a rapid post-LGM rise in sea temperatures.” Some ice may have been shore-fast, producing land-like connections among islands and the mainland as well as restrictions on subsistence and travel by watercraft. Information from marine sediment cores such as diatom species and biochemical markers can provide such insights, but we are not able to acquire adequate data at this time.
9. Conclusions
The theoretical implications of the Bering Transitory Archipelago are profound, including an even better pathway to America than overland passage across the Bering landmass itself. The Stepping-Stones hypothesis satisfies all four requirements for a viable hypothesis about the first human crossing to America: a source population in Asia, a pathway with abundant sustenance, settlements in North America soon after but not before, and an isolated sanctuary. Literally, from 30,000 to 8000 BP a facilitative corridor did exist, and it was far better than the interior deglaciation corridor that dominated scientific debate for eight decades.
The very existence of a previously unappreciated archipelago enhances both the Kelp Highway and Beringian Standstill hypotheses. The highway was originally conceived as a linear pattern of nearshore biomes associated with coasts, islands, and shallow seas stretching beside the advancing and retreating shores. An archipelago would imply far greater expanses of such biomes surrounding scores of islands while still clinging to mainland coasts. It would have advanced seaward from 30,000 to 20,500 BP, then retreated landward from 20,000 to today.
Simultaneously, the Beringian Standstill hypothesis gains a vast new platform to nurture migrants for thousands of years before they moved on to North America. How better to describe an archipelago than Hoffecker’s “distributed across a large area and perhaps even subdivided among several regions” (1998). Clearly, it would provide a sanctuary where Beringians could exchange DNA among themselves for thousands of years with little contribution from Asia while they became a separate people.
Acknowledgments
We sincerely thank Jon Erlandson, Geoff Bailey, E. James Dixon, Thomas Royer, Daniele Melini, and Charles Finkl for reviewing early drafts of our manuscript and offering helpful, even essential, advice. We deeply appreciate the cordial and professional manner in which François Chabaux, co-editor-in-chief of Comptes Rendus Geoscience, and three anonymous reviewers evaluated our research and improved it tremendously with their expert knowledge of the diverse topics necessarily involved in age-old mysteries of place, time, and human behavior. The figures have been drawn using the Generic Mapping Tools (GMT) of Wessel and Smith (1998). The Sea-Level Equation solver can be downloaded from: https://gmd.copernicus.org/articles/12/5055/2019/gmd-12-5055-2019.html.
GS is funded by a FFABR (Finanziamento delle Attività Base di Ricerca) grant of MIUR (Ministero dell’Istruzione, dell’Università e della Ricerca) and by a DiSPeA research grant.