

Abridged English version
1. The growth of a cell population structured according to three cellular states is first considered. Its properties are expressed within the framework of Delattre's transformation systems theory, which allows an accurate formalization of the autocatalysis process:
- – occurrence of five classes of functional equivalence: one source y1, one transient variable at the stationary state x, three cell classes: juvenile y2, mature y3 (the only cells that are able to divide) and senescent y4 cells;
- – native dissymmetry of the two daughter cells at the time of any mitosis: y3→y2+y3;
- – transformations governing transitions between classes:
Model I is characterized by a non-limiting source (autocatalysis only). Population growth is of the exponential type with a rapid change towards a stable structure (constancy of the relative numbers of the three cell states).
Model II is developed for a source in limited amount (autocatalysis + competition). In this case, Y growth is of the sigmoidal type. This model is used for putting forward an interpretation of the ‘logistic law’ applied to Y(t) kinetics.
2. Interpretation of the logistic function. Richards–Nelder's generalized logistics, with the adjunction of a lower asymptote Y≠0, fits very satisfactorily the data simulated by model II. A biological interpretation of the logistics is then put forward. For that purpose, the concept of growth potential (which is distinct from the classical absolute growth rate dY/dt) is introduced. The growth potential is defined, with reference to the basic process, as the instantaneous capacity of autocatalysis of the population. At any instant, the occurrence and the amount of mitoses are determined by the ‘present mitotic resources’, i.e. concomitantly source + available (non senescing) mature cells, that is k1(1−k4)y1(t)y3(t).
The time dependence of the gradient of this potential parallels the growth acceleration variations d2Y/dt2 of the total population.
The definition of the temporal structure of a logistic growth is recalled, with the discretization of the cursus Y(t) into a series of four phases depending on whether the acceleration is positive or negative, increasing or decreasing. The occurrence of these four phases and their delimitation correspond exactly to the temporal variations of the growth potential. Therefore, the logistic law can be interpreted in terms of a formal model based on the time dependence of the instantaneous mitotic resources of a population that is structured according to a criterion of state.
3. An extension to five (three juveniles, one mature, one senescent) cellular states with other transition rules corroborates the preceding results. The generalization of this type of model to other (dimensional or ponderal) growth types is finally considered. With this change in the physical nature of the variables, the choice of the structuring criterion of the population implies the principle of a spatial discretization of a continuous medium into ‘growth elements’.
1 Introduction
La grande généralité des courbes sigmoı̈des représentatives de la croissance de systèmes biologiques de type densité-dépendant dans le cas monotone rend délicat le problème de leur interprétation. Cette sorte d'ubiquité, au regard de la diversité de la nature physique des systèmes concernés, rend en effet difficile la recherche d'une base théorique précise pouvant justifier le modèle apte à générer de telles courbes. De fait, à l'exception de certains modèles de grandissement cellulaire fondés sur les propriétés mécaniques de la paroi et du cytoplasme ([1,2] par exemple), peu de travaux se sont attachés à cette question ([3], pour un cas particulier de la fonction de Gompertz, par exemple). La plupart visent plus la généralité formelle du modèle et sa flexibilité (avec souvent une augmentation du nombre de paramètres) que son explication en termes biologiques [4–6]. Dans ce but, nous avons proposé [7], dans le cadre des systèmes de transformation de Delattre [8,9], un modèle autocatalytique permettant de rendre compte de courbes sigmoı̈des de dissymétrie variée dans le cas d'une seule variable y(t). Précisons toutefois que ce modèle se rapporte au formalisme général de Bertalanffy [5], plutôt qu'à la forme classique de la logistique qui sera discutée ici.
Rappelons que, selon la théorie logistique de la croissance, les variations temporelles de la variable étudiée y(t) suivent la fonction dite logistique (selon la terminologie de Verhulst [10]) considérée ici selon l'expression de Richards–Nelder [11]. Plaçons-nous ici dans un cas plus général où nous supposons l'existence d'une asymptote inférieure y=K0≠0. Dans ces conditions, la théorie logistique repose sur l'hypothèse suivante :
| (1) |
| (2) |
La présente note élargit le problème aux cas de populations structurées afin de tenir compte de la dépendance de l'activité de croissance vis-à-vis de l'âge ou de l'état des éléments constitutifs de la population. D'une part, cela permet de lever le caractère parfois peu réaliste des modèles usuels de croissance comme la logistique classique, bien que le recours à un modèle multilogistique puisse être parfois une alternative intéressante en décomposant le processus en deux ou plusieurs composantes logistiques [12]. D'autre part, cette approche devrait permettre de mettre en rapport cinétique et structure, qui sont deux caractères fondamentaux de toute dynamique biologique.
2 Système à trois états cellulaires
Prenons pour exemple une population cellulaire dont nous considérons la croissance en nombre d'éléments, et qui est caractérisée par les propriétés suivantes.
- – Tout cellule passe par une suite de trois états successifs, dits juvénile, mature et sénescent. Seul l'état mature est apte à la division cellulaire. L'état sénescent constitue un état terminal.
- – Toute mitose est dissymétrique : elle génère deux cellules-filles d'état initial différent. Par état initial, nous entendons le « degré de juvénilité », ou temps nécessaire pour acquérir la maturité de division.
- – La croissance de la population (en nombre de cellules) est subordonnée à l'existence d'une source nécessaire au déclenchement des mitoses. Sans préjuger de sa nature, ce terme de source est phénoménologiquement l'équivalent des notions écologique de capacité biotique d'un milieu (carrying capacity) ou génétique de capacité mitotique d'une lignée cellulaire (nombre maximum de divisions).
Exprimons les propriétés biologiques élémentaires énoncées ci-dessus à l'aide de la théorie de Delattre dont le formalisme s'applique notamment au processus d'autocatalyse en mettant en jeu une variable transitoire à l'état stationnaire.
2.1 Existence de cinq classes d'équivalence fonctionnelle
- E1 : source y1 déterminant la possibilité des cellules matures à se diviser (son épuisement implique l'arrêt de toute mitose).
- E2 : variable x de transition pré-mitotique que l'on suppose à l'état stationnaire.
- E3 : cellules juvéniles y2.
- E4 : cellules matures y3 (seules aptes à se diviser).
- E5 : cellules sénescentes y4.
Ce critère « état cellulaire » structure la population cellulaire en trois classes d'état correspondantes (juvénile, mature, sénescent). À tout instant, elle est ainsi caractérisée par sa distribution dans ces classes E3, E4 et E5, dont les fréquences relatives explicitent la structure de la population.
2.2 Transformations (Fig. 1)
- – Toute cellule est soumise aux transitions temporelles de classes : E3 → E4 → E5 (changements d'état) définies par les coefficients de vitesse ki.
- – La division des cellules matures s'opère via la variable transitoire x, dont la formation requiert conjointement les variables y1 (source) et y3 (autocatalyse).
- –
Pour toute mitose, la règle de production cellulaire est : y3→y2+y3. La cellule-fille y2 doit subir une maturation y2→y3 avant de pouvoir se diviser. La cellule-fille y3 n'est apte à se diviser que si la source est présente, mais compte tenu d'une probabilité de sénescence (coefficient k4). Soit :
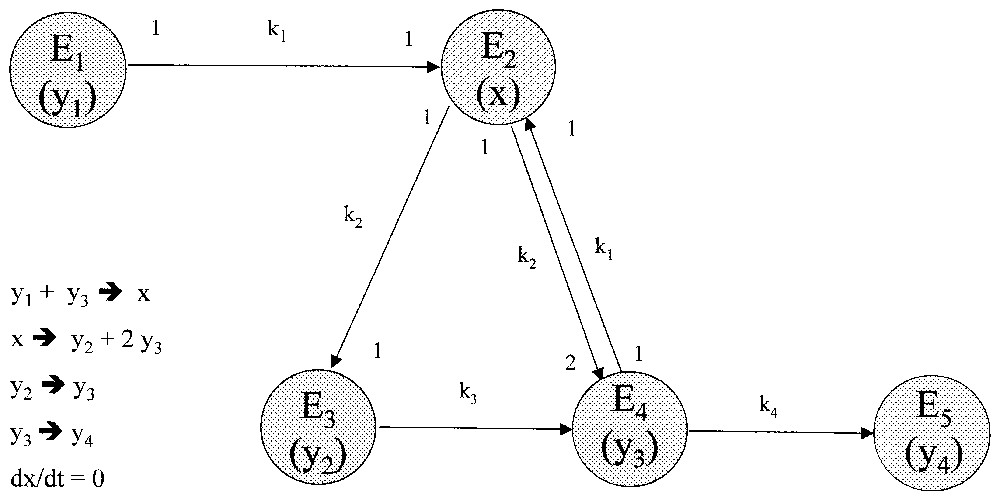
Système à trois états cellulaires : y2 (juvénile), y3 (mature), y4 (sénescent). y1 : source ; x : variable transitoire stationnaire.
Ces transformations sont représentées par un graphe orienté (Fig. 1) et s'expriment par les équations suivantes après élimination de la variable x (sachant que dx/dt=0) :
| (3) |
Deux cas sont à considérer selon que la source y1 est ou non en quantité limitée.
2.2.1 Modèle I : autocatalyse avec source non limitante
Supposant y1=S=Cte, (3) se réduit au système linéaire :
| (4) |
Système à trois états cellulaires (modèle I) : évolution de la structure (proportions des classes cellulaires). S=5 ; ki=0,1. Les différentes courbes correspondent à diverses conditions initiales. Quelles que soient celles-ci, la population tend vers une structure stable {0,41 ;0,49 ;0,10}.
Au système (4) correspond, par discrétisation du temps, un modèle matriciel dont la matrice de transition (différente des matrices de Leslie stricto sensu) est :
Soit
Exemple : avec S=5 et les ki=0.1 (Fig. 2), λ1=1.49 et l'on a une croissance exponentielle (λ1>1). La structure stable (proportions) est : {0.41,0.49,0.10}.
2.2.2 Modèle II : autocatalyse + compétition (source limitée)
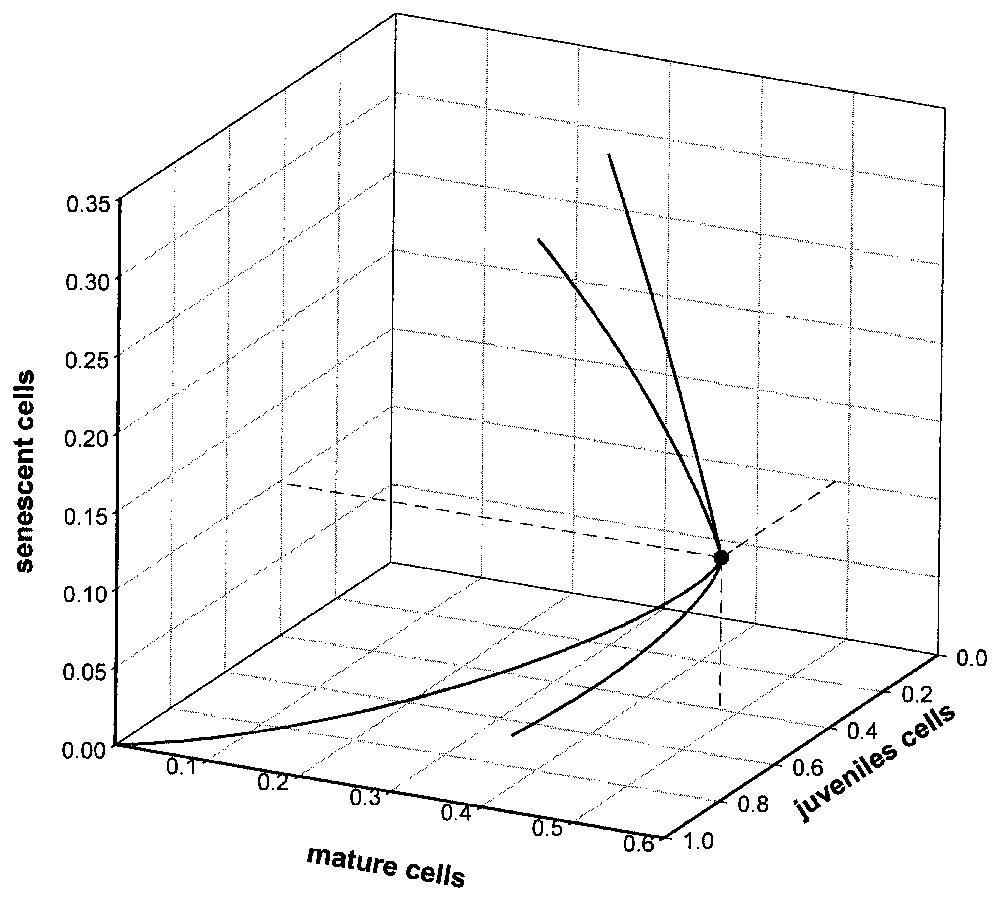
La résolution numérique de (3) montre une croissance de type sigmoı̈de pour y4 ainsi que pour l'effectif cellulaire total Y=y2+y3+y4, alors que y2 et y3 passent par un maximum avant épuisement de la source y1 (Fig. 3). La structure est différente du modèle précédent (Fig. 4), puisque, par défaut de renouvellement (source limitée), la population évolue vers un état homogène stable, toutes les cellules parvenant en E5.

Système à trois états cellulaires (modèle II) : cinétique des différentes classes et de la population totale. ki=0,1 ; y(0)={10,1,0,0}.
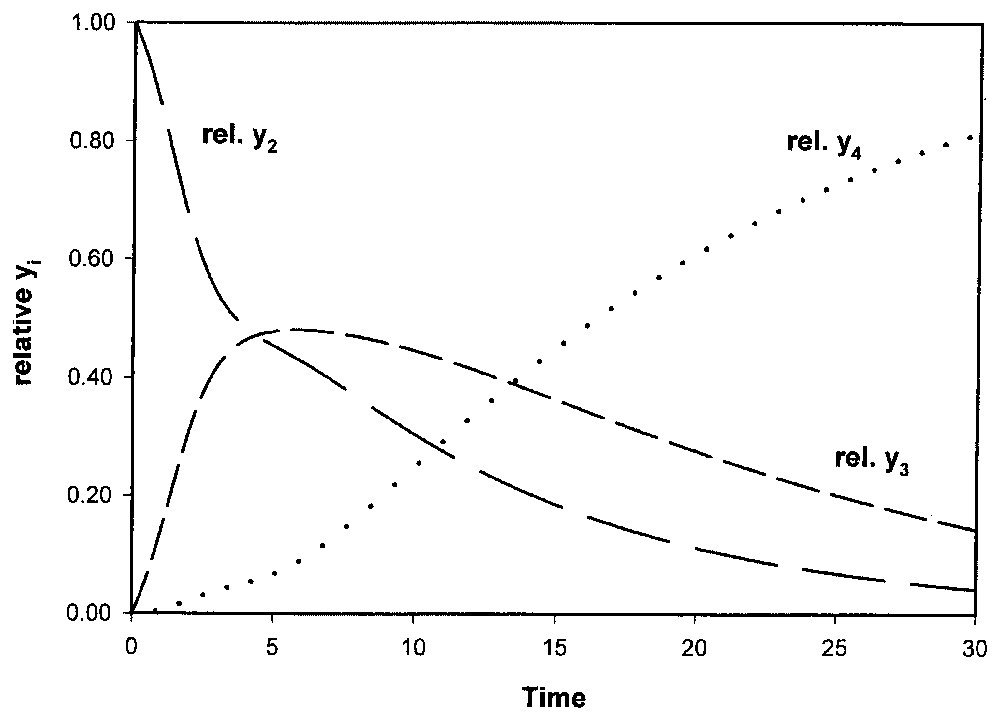
Système à trois états cellulaires (modèle II) : structure de la population (yi relatifs). Conditions de la Fig. 3.
Le modèle matriciel correspondant est évidemment très différent du précédent car la matrice M (où intervient la source qui est ici une variable) n'est pas constante mais densité-dépendante. On peut l'écrire sous la forme réductible suivante en explicitant la variable y3 sujette à l'autocatalyse :
L'analyse de stabilité par approximation linéaire [14, (p. 238)] montre l'existence d'un état stationnaire stable des fréquences qui est en réalité un état homogène {0,0,1}.
Détaillons la dynamique du modèle II. Une fonction logistique généralisée du type (2) s'ajuste correctement aux données de Y telles que simulées par le modèle (3) (Fig. 5), en posant y=Y. Ce modèle génère des courbes de croissance de dissymétrie variée par rapport au point d'inflexion. Dans le cas particulier des logistiques avec K0=0 (qui est le plus souvent exploité), le paramètre n mesure cette dissymétrie (selon que n<1 ou n>1). Mais, dans le cas général traité ici, où K0≠0, la symétrie exacte a lieu pour n racine de (1+n)1/n=2K1/(K1−K0). La dissymétrie des courbes Y(t), notée par le rapport Yinflexion/(Ymax/2), dépend des valeurs relatives des coefficients de vitesse ki. Ce rapport vaut approximativement 1 si
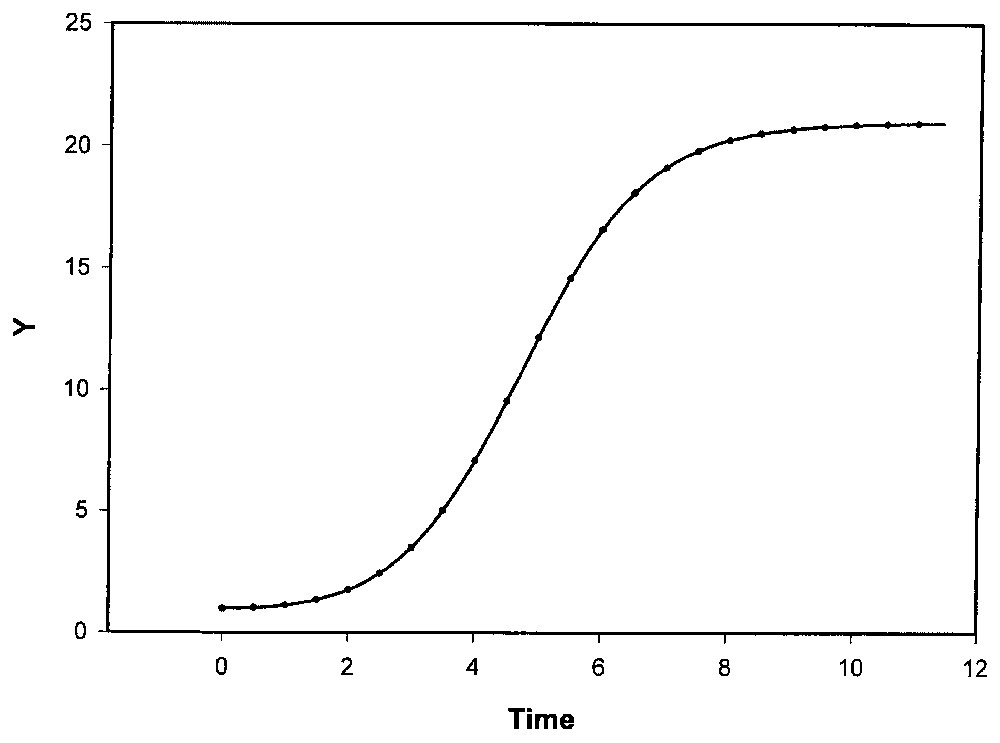
Système à trois états cellulaires (modèle II) : données simulées par le modèle II (•) (conditions de la Fig. 3) et ajustement d'une logistique généralisée. K0=0,8393; K1=20,1557 ; a=0,9998 ; n=0,8591. R2ajust. = 0,999997.
2.3 Structure temporelle de la croissance globale Y(t)
Rappelons que la cinétique logistique comporte comme points singuliers de l'activité de croissance les deux extremums de la dérivée seconde (accélération Γ) d2y/dt2 et son annulation d2y/dt2=0 [5,15]). Ces valeurs délimitent quatre phases de croissance dénommées {G1,G2,G3,G4} définies par les valeurs du couple (V,Γ), c'est-à-dire, selon que l'accélération est positive ou négative, croissante ou décroissante. Les durées et quantités de croissance (accroissements) pour chacune de ces phases, en valeurs relatives, caractérisent la structure temporelle de croissance. Ce schéma est donc indépendant de l'ordre de grandeur de la variable en croissance, permettant ainsi de caractériser le phénomène de croissance d'un point de vue strictement cinétique et non dimensionnel. Il montre en particulier l'existence d'une éventuelle phase prédominante, soit en durée, soit en quantité de croissance. Nous l'avons utilisé pour discriminer par exemple différents types cellulaires d'un même organisme [12]. Soulignons que la structure temporelle de Y (discrétisation du cursus de Y(t) en quatre phases) et la structure instantanée de la population {yi=1,…,k classes cellulaires} sont deux notions distinctes.
La structure temporelle de Y(t) dépend des coefficients ki. Elle analyse le cursus de croissance plus finement que ne le permet la prise en compte usuelle de la position du point d'inflexion comme indice de dissymétrie. Le Tableau 1 en donne quelques exemples. On voit que la flexibilité du modèle relève de l'ensemble des coefficients ki, c'est-à-dire de l'ensemble des transitions de classes, alors que l'usage habituel de la fonction (2) se réfère au seul paramètre n. La structure temporelle caractéristique de la logistique simple de Verhulst (K0=0 et n=1), c'est-à-dire l'égalité G1 et G4 d'une part, et G2 et G3 d'autre part [5], s'observe très sensiblement dans le cas d'une égalité de k1 et k4. Lorsqu'il y a nette dissymétrie, la prédominance de la phase G1 (resp. G4) en durée relative est déterminée par un rapport k1/k4>1 (resp. <1), en même temps que k3 contribue fortement à ce que la phase G3 soit prédominante en quantité de croissance réalisée. De la simple description de la cinétique logistique en une suite de phases Gi, on passe ainsi à son explication par des transitions de classes.
Structure temporelle de croissance en rapport avec les coefficients de vitesse ki
| Coefficients ki | Durée des phases (%) | Quantité de croissance par phase (%) | ||||||
| G 1 | G 2 | G 3 | G 4 | G 1 | G 2 | G 3 | G 4 | |
| 0,5,0,1,0,5,0,1 | 41 | 12 | 18 | 29 | 23 | 26 | 36 | 15 |
| 0,05,0,1,0,3,0,2 | 19 | 12 | 12 | 57 | 16 | 30 | 30 | 24 |
| 0,1,0,1,0,1,0,1 | 36 | 14 | 14 | 36 | 19 | 30 | 30 | 21 |
3 Interprétation du modèle logistique de croissance
La bonne adéquation d'une fonction logistique (2) à la croissance globale d'une population structurée (Fig. 5) permet de rechercher la signification biologique de cette fonction classique. L'interprétation proposée ne se situe pas au niveau de la variable globale Y. À ce niveau, en effet, cela se bornerait à reprendre l'hypothèse de base, purement formelle, rappelée plus haut.
Recherchons plutôt le rapport existant entre les points singuliers notés ci-dessus pour la cinétique globale Y(t), et l'évolution de la population à partir des variations chronologiques (i) du nombre de cellules en division, (ii) de la capacité mitotique de la population. Utilisons pour cela les notions suivantes.
Capacité totale de croissance. La taille maximale de la population en fin de croissance dépend exactement des conditions initiales puisque Ymax=2y1(0)+y2(0)+y3(0)+y4(0) (les ki influant sur la vitesse et la durée de croissance). À tout instant, l'écart entre cette grandeur et Y(t) exprime la capacité actuelle de croissance. Ceci reprend la notion classique de carrying capacity rappelée précédemment. Elle n'a ici que peu d'intérêt, car elle ne peut expliquer les variations de la vitesse du processus.
Activité instantanée de croissance. Le recours classique à la vitesse spécifique R n'a pas de sens ici, puisqu'elle ne tient pas compte de sa dépendance vis-à-vis de l'état cellulaire. Définissons l'activité de croissance par la possibilité d'occurrence de nouvelles mitoses, formalisée par la transformation :
Notons que cette « capacité d'autocatalyse » ne dépend pas seulement de la source disponible, ce qui dénue d'intérêt la grandeur dy1 (qui ne pourrait servir que de mesure du degré d'avancement de la croissance). Elle dépend aussi, conjointement, de la quantité de cellules matures disponibles (celles n'entrant pas en sénescence) y3∘. Soit f(y1(t),y3∘(t)). Adoptons pour f, en tenant compte des coefficients ki, la grandeur suivante, appelée potentiel de division :
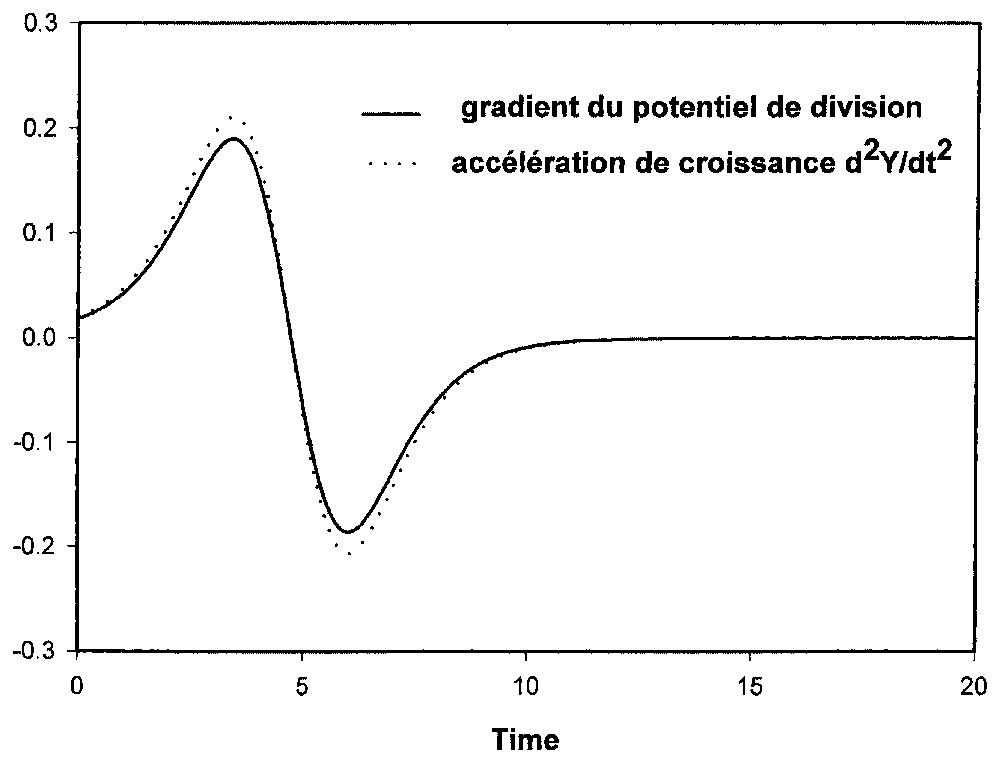
Système à trois états cellulaires (modèle II) : gradient du potentiel de division et accélération de croissance (conditions de la Fig. 3).
4 Généralisation
Ce système « minimal » à trois classes d'état peut être aisément étendu par ajout de processus de mortalité et/ou augmentation du nombre d'états cellulaires ainsi que par changement de l'état initial des deux cellules-filles. Ces modifications n'altèrent nullement le principe de l'analyse et de l'interprétation proposée.
Exemple : soit un système avec source en quantité limitée (y1) et cinq états cellulaires (trois juvéniles y2,y3,y4, un mature y5 et un sénescent y6). Toute mitose de cellule mature non engagée vers la sénescence donne deux cellules juvéniles différentes : y5→y2+y4. Le graphe correspondant résume les différentes transformations (Fig. 7). Comme précédemment, avec une source limitée, la croissance de la population totale Y=y2+y3+y4+y5 est de type sigmoı̈de. Avec le modèle matriciel correspondant, l'analyse de stabilité de la matrice de transition (densité-dépendante et réductible) conduit aux mêmes conclusions que précédemment.
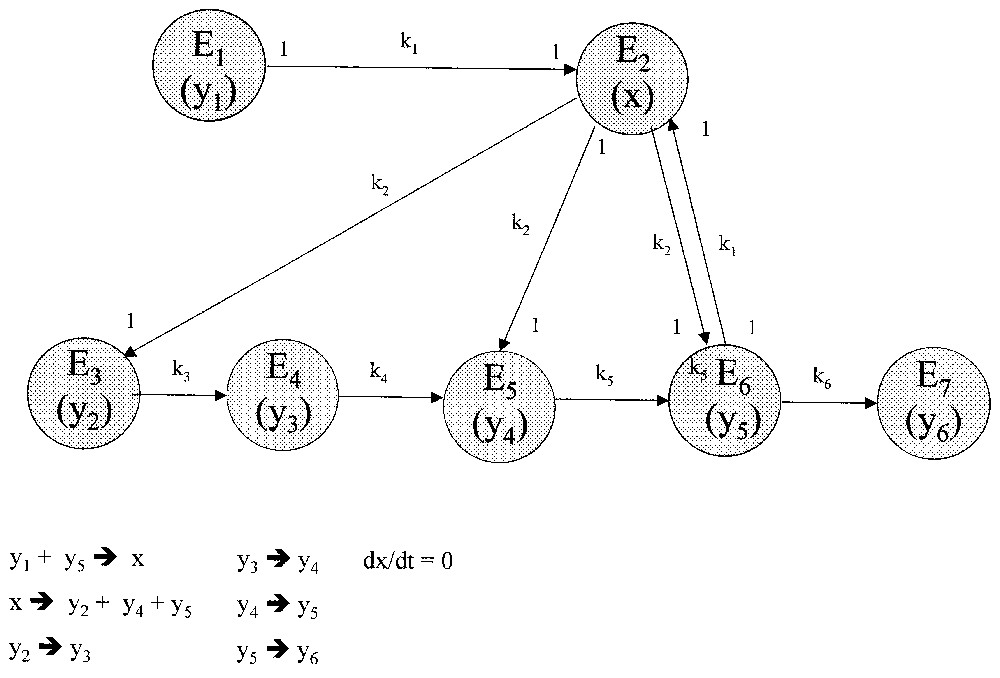
Système à cinq états cellulaires : trois états juvéniles (y2,y3,y4), un état mature (y5) et un état sénescent (y6).
D'autre part, les points singuliers de la structure temporelle de Y(t)(calculés après ajustement par une logistique généralisée) correspondent, comme avec le système précédent, à ceux du gradient du potentiel de division défini par les ressources mitotiques actuelles, soit : pot.div.(t)=k1(1−k6)y1(t)y5(t).
5 Discussion
Le type de modèle présenté concerne le cas particulier d'une croissance d'une population en nombre d'éléments constitutifs. Il peut être étendu formellement à d'autres cas de croissance de population (individus par exemple). En outre, son application à d'autres types de croissance – dimensionnelle ou pondérale – (grandissement cellulaire, croissance d'organes ou d'organismes) peut également être envisagée. Dans ce cas, on ne traite pas des entités discrètes et dénombrables comme des effectifs cellulaires, mais des « éléments », de nature topologique, d'un ensemble physique continu. Rappelons le cas des modèles discrets fondés sur les grammaires formelles (comme les L-systèmes). Outre la multiplication cellulaire qui constitue leur champ principal d'investigation, le principe de ces modèles s'applique également, avec discrétisation spatiale, au grandissement cellulaire lui-même [16].
Notons brièvement une comparaison entre les L-systèmes et le modèle proposé ici. Le type de critère de structuration de la population est analogue (existence d'un nombre limité d'états cellulaires). Mais les transitions entre états sont formalisées différemment : « règles de production » avec discrétisation du temps pour les L-systèmes, ou équations différentielles de transformations pour notre modèle. Ces systèmes discrets ont fait l'objet de diverses applications (croissance d'algues filamenteuses et de méristèmes, par exemple [17]). Par rapport à ces « automates cellulaires », la particularité de notre modèle réside d'abord dans la prise en compte explicite du processus d'autocatalyse que nous couplons à un processus de compétition. Mais surtout notre modèle permet : (i) de quantifier cette notion de « potentiel de croissance » défini par les ressources mitotiques instantanées de la population ; (ii) de relier la cinétique « globale » d'une population au concept fondamental de « structure temporelle » de croissance.
Un autre aspect important est la flexibilité des courbes de croissance générées par notre modèle, correspondant à la diversité des sigmoı̈des dissymétriques observées (voir, par exemple, la croissance de l'hypocotyle de Lupinus albus [5, (p. 305, Fig. 4)]. Notre modèle met en relation ce caractère (souvent négligé quoique caractéristique de certaines croissances) avec les valeurs relatives des coefficients de transformations (Tableau 1).
L'introduction d'une source limitée de nutriments comme variable (sujette à déplétion) dans le cadre d'un système dynamique a été souvent exploitée pour modéliser une cinétique de croissance [18,19]. Elle rend compte évidemment du caractère sigmoı̈de variable de nombreuses courbes de population cellulaires, mais sans fournir, par elle-même, d'explication aux points singuliers du cursus. De leur côté, les diverses études de populations structurées [20] ignorent généralement la notion de structure temporelle de croissance. Notre propre analyse montre ce que peut apporter la conjonction de ces deux aspects. Elle souligne que l'explication d'une cinétique de croissance requiert de satisfaire les deux conditions suivantes : (i) quantifier correctement l'activité instantanée de croissance (ce qui implique de structurer la population) ; (ii) recourir à un formalisme, précis et de portée générale, pour l'autocatalyse en tant que processus de base de toute croissance.
6 Conclusion
1. La structure temporelle de la croissance logistique (au sens de la décomposition du cursus temporel en phases selon les valeurs du couple (V,Γ)) ne correspond pas à des caractéristiques particulières de la structure de la population (sa distribution en classes d'état). Son interprétation repose sur les variations du potentiel de division qui exprime les ressources mitotiques instantanées de la population. Celles-ci évoluent au cours de la croissance par le fait (i) du caractère limitant de la source conditionnant l'état limite en fin de processus, (ii) de la transition entre classes d'états qui détermine, à tout instant, la quantité de cellules aptes à se diviser.
2. Ceci nous apparaı̂t comme une base intéressante fournissant une interprétation, en termes biologiques, de la fonction logistique. Celle-ci ne doit plus être considérée comme un simple outil empirique d'ajustement, mais comme un modèle formel de population structurée sur un critère d'état et soumise au jeu conjoint autocatalyse + compétition. Par ailleurs, l'intérêt de la notion de structure temporelle d'un processus s'en trouve renforcé, puisque les caractéristiques de la suite de phases de croissance sont dépendantes des coefficients de transition entre classes d'état. D'où l'on peut envisager son application à d'autres types de croissance, monotone ou non.