

1 Introduction
Animals live in a complex environment, which varies in time and space, and they have to behave accordingly to these variations. Until recently, most studies on animal behaviour tested the influence of environmental factors separately; but in natural conditions, animals interact with several factors and they have to take them into account simultaneously in order to survive and to maximize their fitness. Several authors suggest that decisions made by animals, i.e. where, when, how to forage, to reproduce... could result of trade-off between costs and benefits associated with different environmental situations (reviewed in [1–3]). Here, we tested the simultaneous influence of prey and conspecifics on web-building behaviour in spiders.
Like in many other animal species, in spiders, an increase of food intake has a direct positive effect on growth rate and reproductive success [4–6], thus foraging decisions would affect spider fitness. Web building associates costs and benefits: a web requires investments in time (web building duration) and energy (silk production and physiological effort), but provides energy for survival and reproduction through prey capture. The ratio cost/benefit could vary with environmental conditions. Web construction and web characteristics materialize the foraging investment and thus reflect the foraging decisions of spiders, and could be seen as a response of spiders to their energetic needs and to environmental characteristics. Therefore, orb-weaving spiders are interesting models to study the influence of environment on behaviour.
Variations in web characteristics may be influenced by the spider state – e.g. growth, leg regeneration, feeding, silk supplies, egg production, experience – [7–13], and environmental conditions – e.g. light, temperature, humidity, wind, gravity, web supports, presence of prey or conspecifics – [14–21]. These variations underline the behavioural plasticity that spiders exhibit to fit their web to the situation.
It has been shown that in natural habitat, Zygiella x-notata (Clerck), an orb-weaving spider (Araneae, Araneidae) often lives in aggregations in which web building is influenced by conspecifics [18], and that under laboratory conditions prey influenced web construction and web characteristics [19].
Under natural conditions, prey and conspecifics are present simultaneously, so web building may be affected by them. We can hypothesize that spiders should take into account these two factors together to adjust their web to the situation. As web building decision and web parameters (web building duration and web characteristics) could reflect the spider foraging decision, we tested the influence of prey and conspecifics on these parameters.
2 Materials and methods
2.1 Spiders
Z. x-notata is commonly found on human constructions (fences or window frames). Most of the time, they renew their web daily at the end of the night [22]. First, the spider builds a frame, then radii and an auxiliary non-sticky spiral before the final sticky capture spiral. The web usually has a free sector. The retreat is connected to the hub by a signal thread. During the day, the spider stays in its retreat.
We collected adult females in September 1996 and 1997 in several localities (Center-East part of France). We kept them individually into plastic boxes (10×7×2.5 cm) in laboratory, under constant abiotic conditions (temperature: 22–24 °C; 12 h of light per day: 20:00–08:00). They could not build an orb web in the boxes, but they laid silk and captured prey; they received two cricket larvae, Gryllus bimaculatus (Orthoptera, Gryllidae), twice a week and were provided with water three times a week. Flies, Calliphora vomitaria (Diptera, Calliphoridae), were used as prey during experiments. All the prey had approximatively the half of the spider size.
Before experiments, spiders were weighed with scales (Sartorius Basic BA110S, ±0.1 mg), and we measured their total body length with an electronic digital calliper rule (±0.05 mm). Each spider was marked individually with a dot of paint (enamel paint, Revell, Germany) on the postero-dorsal part of abdomen.
2.2 Experiments
Experiments were carried out in 1996, 1997 and 1998. Spiders were introduced into clean wooden frames (50×50×10 cm) closed by two window panes at dark-light transition (t0). The frame size was wide enough to allow two spiders to build a web [17,18]. Spiders were assigned randomly to four groups: one group in which there was only one spider per frame (‘control’ group, N=94), one group in which we put one spider and four flies per frame (‘prey’ group, N=84), one group with two spiders per frame (‘spider’ group, N=81), and one group in which we put simultaneously two spiders and four flies per frame (‘spider + prey’ group, N=61). Spiders and prey were introduced at the same time (t0) into the frames, which were distributed randomly in the experimental room to avoid room heterogeneity effects.
Initially the same number of spiders was assigned to each group, but we excluded spiders that laid eggs during experiments or ate the prey or the conspecific before building their web, because it entailed modifications of the spider internal state, which could lead to changes in web-building behaviour [11,12,23]. Moreover, for the ‘spider’ and ‘spider + prey’ groups, we only took into account the pairs of spiders for which the two individuals could be kept for the analysis (cf. above). The spiders used for experiments were heavier and larger in 1997 than in 1996 and 1998, but there was no difference between the four groups (ANOVA, weight: F3,460=0.18; length: F3,460=0.52 NS).
2.3 Parameters
Observations started at t0 and were done half-hourly during the first 24 h, but continuously when a spider was building a web. After the first 24 h, two observations were done at 48 h and 72 h. We noted the first construction of each spider, and recorded different parameters: latency to start building (from t0 to the beginning of the construction), web building duration (from the first radii construction to the hub remodelling), and web characteristics (the diameter measured as the distance between the two most distant spirals of the orb, the number of radii, the number of spirals in the lowest sector of the web, and the distance between the innermost and outermost spirals in this sector). We calculated the mesh size as the ratio between the innermost–outermost spiral distance and the number of spirals in the lowest sector.
In the ‘control’ group, most of the parameters measured were correlated. As diameter and mesh size were considered as good indicators for respectively web size and web structure, only these were kept for the analysis.
The distribution of the number of web building spiders was studied over time. We called ‘anticipating spiders’, the spiders of the experimental groups (‘prey’, ‘spider’ and ‘spider + prey’) that built a web before 18 h (after t0), because most of the spiders of the ‘control’ group (65/66) built after this period.
In the ‘spider’ and ‘spider + prey’ groups, there were two spiders per frame which could decide to build a web, and when both spiders spun the timing of web construction differed; then we divided these groups into sub-groups: the ‘first spiders’ sub-group for spiders building first in a pair, and the ‘second spiders’ sub-group for spiders building in second. But when only one spider of a pair built a web, it was allocated to the ‘first spiders’ sub-group and the non-building spider to the ‘second spiders’ sub-group. When the two spiders of a pair did not build, we allocated randomly one of these spiders to each one of the two sub-groups ‘first spiders’ and ‘second spiders’.
For data analysis, we combined the data of the three years (1996, 1997, 1998). We chose this method because our experiments were nicely balanced: we tested each year the same number of spiders for each one of the four groups, and the spiders of these groups did not differ. It means that the effect of year acted in the same way on each group; so the analysis by combined data was justified.
In the ‘spider’ and ‘spider + prey’ groups, the two spiders decided to build or not and when building. They constituted a ‘first spiders’ and a ‘second spiders’ sub-group. The spiders of these sub-groups were not randomised, since they were separated by the timing of web building, which depended on the spider itself; therefore, we could not compare the data of these sub-groups with those of the randomised single spider groups (‘control’ and ‘prey’). To make this comparison, we split up the single spider groups into two sub-groups: ‘first control’ and ‘second control’, ‘first prey’ and ‘second prey’; among the spiders of the ‘control’ and ‘prey’ groups, we drew pairs of spiders at random, and for each pair we assigned the spider building first (time after t0) to the sub-group ‘first control’ or ‘first prey’, and the spider building in second to the sub-group ‘second control’ or ‘second prey’. For each group, to avoid pairing bias, we repeated this procedure 20 times; thus we got 20 times a number of web building spiders. After that, we used the median of this number for comparisons.
We used pairwise Fisher's exact tests for testing comparisons between groups, and, to reduce the probability of incorrectly rejecting null hypothesis as a result of multiple tests, we applied the sequential Bonferroni test [24] to our experiments.
For web building parameters (web building duration and web characteristics), we used a two-way ANOVA design to examine simultaneously the effects of the presence of prey and conspecific. A significant interaction between prey and conspecific would indicate that the two factors interact in the ‘spider + prey’ group.
3 Results
3.1 Percentage of web building
At the end of experiments (t=72 h), the percentage of spiders building a web differed significantly between the sub-groups (Chi-square test: χ27=131.81, P<0.001) (Table 1).
Web building percentage in the different groups. The sub-groups randomly chosen (see Methods) within the ‘control’ and ‘prey’ groups are in italics. P is the statistical result of the Fisher's exact test from a comparison between each sub-group and the corresponding ‘control’ sub-group. All comparisons significant at P<0.005 remained significant after the sequential Bonferroni test
| Group | Number of spiders | Percentage of web building | P | |
| Sub-group | ||||
| Control | 94 | 68.7 | ||
| First | 47 | 91.5 | ||
| Second | 47 | 44.7 | ||
| Prey | 84 | 90.5 | 0.0004 | |
| First | 42 | 100.0 | 0.119 | |
| Second | 42 | 80.9 | 0.0005 | |
| Spider | 162 | 63.6 | 0.419 | |
| First spiders | 81 | 87.6 | 0.571 | |
| Second spiders | 81 | 39.5 | 0.582 | |
| Spider + prey | 122 | 91.8 | <0.0001 | |
| First spiders | 61 | 96.7 | 0.400 | |
| Second spiders | 61 | 86.9 | < 0.0001 |
The presence of prey stimulated web building. In the ‘prey’ group, a higher percentage of spiders built a web than in the ‘control’ group (90.5% vs 68.7%) (Table 1).
The presence of a conspecific did not affect web building. In the ‘spider’ group, the two sub-groups did not differ from those of the ‘control’ group (87.6% vs 91.5% and 39.5% vs 44.7%; Table 1).
In the ‘spider + prey’ group, the ‘first spiders’ sub-group did not differ from the ‘first control’ sub-group (Table 1); however, despite this absence of a significant difference, building percentages (87.6% for the ‘spider’ group, 91.5% for the ‘control’ group, 96.7% for the ‘spider + prey’ group, and 100% for the ‘prey’ group) seemed to indicate an effect of prey.
For the ‘second spiders’ sub-group, a higher percentage of spiders built a web than in the ‘second control’ sub-group (Table 1). The web building percentage of the ‘second spiders’ sub-group of the ‘spider + prey’ group did not differ from that of the ‘second prey’ sub-group.
3.2 Distribution of web construction over time
To study the distribution of the number of web building spiders over time, we divided the observation period into three equal periods: 0–24 h, 24–48 h, and 48–72 h. The percentage of web building differed significantly between the sub-groups for each of the two first periods (Fig. 1) (Chi-square test, 0–24 h: χ27=141.24, P<0.001; 24–48 h: χ27=37.67, P<0.001). For the third period, statistical tests were not valid, because too many expected values were equal to zero, very few spiders built after 48 h.
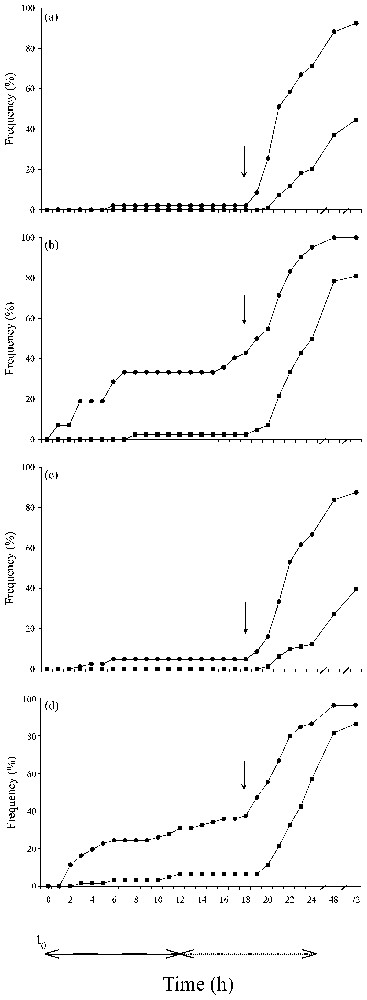
Cumulative percentage of spiders building a web over the 72-h-period for the different groups: (a) ‘control’ group; (b) ‘prey’ group; (c) ‘spider’ group; (d) ‘spider + prey’ group. In (a) and (b), • represents the ‘first control’ or the ‘first prey’ sub-group, and ■ the ‘second control’ or the ‘second prey’ sub-group. In (c) and (d), • and ■ represent respectively the ‘first spiders’ and ‘second spiders’ of the couples of spiders. Inside each graph, the arrow indicates the percentage of spiders that built a web before 18 h, so that anticipated web construction. At the bottom of the figure, continuous and dotted lines indicate respectively when the light was on or off, and t0 corresponds to the introduction of spiders into the frame.
In the ‘prey’ group, 22.6% of the spiders anticipated web construction and built before the spiders of the ‘control’ group (Fig. 1) (Fisher's exact test: P<0.001, Bonferroni corrected), and a higher percentage of spiders of this group than in the ‘control’ group built during the first period (Fisher's exact test, 0–24 h: P<0.001; 24–48 h: NS; 48–72 h: NS, Bonferroni corrected). So the presence of prey stimulated web construction during the first 24 h, and some spiders anticipated web construction.
In the ‘spider’ group, only 4.9% of the spiders of the ‘first spiders’ sub-group and no spider of the ‘second spiders’ sub-group anticipated web construction and built before the spiders of their respective ‘control’ sub-group (Fig. 1) (Fisher's exact test: NS). Moreover, the percentage of web building spiders in the two sub-groups of the ‘spider’ group did not differ from the control for each period (Fig. 1) (Fisher's exact test: NS). So the presence of a conspecific did not affect web construction over time.
In the ‘spider + prey’ group, 37.7% of the spiders of the ‘first spiders’ sub-group anticipated web construction and built before the spiders of the ‘first control’ sub-group (Fig. 1) (Fisher's exact test: P<0.005, Bonferroni corrected). The percentage of web building spiders between the ‘first spiders’ sub-group and the ‘first control’ sub-group did not differ for each period (Fig. 1) (Fisher's exact test: NS); however, despite this absence of a significant difference, building percentages for the first period (66.7% for the ‘spider’ group, 72.3% for the ‘control’ group, 86.9% for the ‘spider + prey’ group, and 92.9% for the ‘prey’ group) seemed to indicate an effect of prey.
For the ‘second spiders’ sub-group of the ‘spider + prey’ group, 6.6% of the spiders anticipated web construction and built before the spiders of the ‘second control’ sub-group (Fig. 1) (Fisher's exact test: NS), and a higher percentage of spiders built than in the ‘second control’ sub-group during the first period (Fig. 1) (Fisher's exact test, 0–24 h: P<0.005; 24–48 h: NS; 48–72 h: NS, Bonferroni corrected). The behaviour of the ‘second spiders’ of the ‘spider + prey’ group did not differ from that of the spiders of the ‘second prey’ sub-group.
3.3 Web building duration
The presence of prey led to a significant reduction of web building duration, whereas the presence of a conspecific had no effect on this parameter (Table 2; Fig. 2a). The interaction between these two factors was not significant (Table 2) indicating that there was no crossed effect, and that only prey affected web-building duration.
Two-way ANOVA on the duration of web building and the characteristics of the webs
| Treatment | Web building duration (min) | Diameter (cm) | Mesh size (mm) | ||||
| N | N | N | |||||
| Presence of prey | 134 | 49.6±1.7 | 175 | 11.5±0.3 | 172 | 1.9±0.0 | |
| Absence of prey | 104 | 54.4±2.0 | 163 | 16.3±0.3 | 163 | 2.6±0.0 | |
| Presence of conspecific | 143 | 50.6±1.8 | 196 | 13.7±0.4 | 194 | 2.2±0.0 | |
| Absence of conspecific | 95 | 53.4±1.8 | 142 | 14.0±0.4 | 141 | 2.2±0.1 | |
| Treatment factor | Web building duration | Diameter | Mesh size | ||||
| df | F | P | F | P | F | P | |
| Presence of prey | 1 | 3.92 | 0.049 | 99.16 | <0.0001 | 115.95 | <0.0001 |
| Presence of conspecific | 1 | 1.37 | 0.243 | 0.67 | 0.413 | 0.57 | 0.452 |
| Presence of prey × presence of conspecific | 1 | 1.29 | 0.257 | 0.09 | 0.764 | 0.02 | 0.879 |

Means (and Standard Error) of (a) web building duration, (b) web diameter, and (c) web mesh size, in the different groups. ∗ When groups differed significantly from the ‘control’ group.
3.4 Web characteristics
The presence of prey led to a significant decrease of web diameter and mesh size, whereas the presence of a conspecific had no effect on these parameters (Table 2; Fig. 2b and c). The interaction between these two factors was not significant (Table 2). So only prey seemed to affect web size and web structure.
4 Discussion
Through this study, in spiders, it was the first time that one searched for a simultaneous effect of two environmental factors on web building behaviour.
In response to different environmental situations, Z. x-notata adapted its web building behaviour. The presence of prey stimulated web building, and spiders modified their web characteristics, building a smaller web with a smaller mesh size, in a shorter time; moreover, some of them anticipated web construction. Once spiders perceived potential prey in their environment, they set up quickly a trap. On the other hand, in the presence of a conspecific, Z. x-notata did not modify its web building behaviour (web building decision, web building duration or web characteristics); this latter result differed from what was found in a previous study [17] where second spiders built a smaller web. This difference might be due to differences of the experiment design.
Our results showed that the spiders took into account the presence of prey in web building. However, even if we did not show any influence of the presence of a conspecific on its building behaviour, previous data on the influence of the conspecific [17,18] might lead us to think that Z. x-notata was able to take into account prey and conspecifics simultaneously and to adapt its behaviour.
The modifications of web building behaviour in the presence of prey suggest that spiders get information on their presence. This ability of spiders to detect prey in their environment before capture might be advantageous because they could assess prey abundance in an unknown patch [25] without a previous investment, and could decide to invest or not energy in web building. We showed that when spiders perceive prey in their environment they build rapidly a small web, which confirmed previous results [19]. This behaviour could have two consequences: first, building a smaller web might be less costly, but secondly, a smaller web has theoretically a lower probability to intercept prey [26]. This latter effect could be balanced by the mesh size: the number of captured prey increasing as the mesh size decreases [27–29]. In our study, spiders built a smaller web with a smaller mesh size, so they could balance the negative effect of the web size reduction on the prey capture probability. But, for a given web size, a smaller mesh size is more costly because it needs a longer capture thread; thus, spiders in the presence of prey did not reduce their web investment, but they reduced the web building duration. As they also reduced the latency before building, in the presence of prey, Z. x-notata favoured the fact to be ready to capture prey.
In this work, we suggested that the presence of a conspecific had no effect on web building behaviour. In the frames, we observed interactions, sometimes aggressive, between the two spiders. This supports the idea that spiders might get information on the presence of the conspecific, but here the presence of a conspecific did not influence web building. In fact, the influence of conspecifics was demonstrated in natural conditions: Z. x-notata may live in aggregations of several individuals where each spider has its own web. In such aggregations, small and large webs alternate and contacts between webs exist through common threads [18]; this shows that, as for colonial spider species [30], constraints on space occupation exist, and web size may be limited by the presence of a neighbour. These data suggest that the influence of conspecifics on web building seems more complex than that of prey and depends probably more on local conditions.
Prey influenced the whole decisions of the web building behaviour of Z. x-notata (building or not, when, how long, which web) even in the presence of a conspecific. The short-term response displayed by spiders in the presence of prey (a quick trap construction) could be a functional explanation that a direct link exists between prey capture and food intake, and spiders could maximize the probability of prey capture when prey are present by setting up rapidly a trap.
In the natural populations of Z. x-notata, spiders usually have a web from which they can get information on their environment. But prey are not continuously present near web; so there might be quantitative or qualitative differences of information perceived by spiders between laboratory and field situations. Observations carried out under natural conditions confirmed this: web renewal usually takes place at the end of the night (4–7 h) and no spider anticipated, and, the characteristics of rebuilt webs depend on the nature of interactions taking place during the day between spiders, their webs and potential prey (prey flight, impact or retention into webs, or capture by spiders) (personal observations) more than the direct presence of prey.
In the laboratory situation, Z. x-notata behaves in the presence of both factors simultaneously as if it takes into account only prey, ignoring the presence of conspecifics. Two interpretations might be put forward: first, in the presence of prey, Z. x-notata would not perceive conspecifics, which is unlikely since some contacts were observed between the spiders into frames; second, Z. x-notata would be aware of the presence of prey and conspecifics, but it would decide its response according to a hierarchical level, responding preferentially to prey. Thus, in the presence of prey, prey capture would be more essential than space management. In the field, the response to prey would not be as strong as in the laboratory because of a dilution of information with other environmental factors. The response to conspecifics would be a middle- or long-term response having an influence on the structuring of spider populations rather than web building. This could be parallel with what we know about colonial orb-weaving spiders where space competition and interactions between spiders determine the structure of the colony [30], and prey have a direct influence on web building.
Acknowledgements
We thank Prof. Fritz Vollrath and Dr Marie-Claude Bel for their helpful comments on an earlier draft of this manuscript. This work received financial support from the CNRS.