

1 Introduction
Angiosperm lignins are composed of syringyl (S) and guaiacyl (G) units derived from sinapyl alcohol and coniferyl alcohol, respectively. These units are interconnected by various ether and carbon–carbon bonds. The ether β-O-4 linkages are both frequent and labile, which makes them the target of the delignification process. In contrast, the carbon–carbon linkages are resistant, especially the biphenyl 5–5 bonds involving the aromatic C-5 position, which is available for inter-unit linkages only in G units [1,2]. This is the reason why conifer wood lignins essentially made of G units are less susceptible to kraft delignification than deciduous wood lignins comprising both G and S units. Tailoring plants more amenable to industrial delignification may be achieved by lowering lignin concentration and/or altering lignin structure to an extent that does not affect plant development and defence (reviewed in [3–5]). Transgenic or mutant plants with modified expression of the genes involved in monolignol biosynthesis have been studied in detail. Taken together, these studies have demonstrated that plants can tolerate the down-regulation of various genes involved in monolignol formation by incorporating uncommon monomers into lignins (as reviewed in [4] and [6]). For instance, down-regulating cinnamyl alcohol dehydrogenase (CAD) in tobacco [7], poplar [8,9] and Arabidopsis [10] mainly results in the occurrence of β-O-4 linked sinapyl aldehyde lignin units, as recently evidenced by the identification of specific indene compounds [11] derived from lignins by the thioacidolysis degradative method [2,12]. This method also provided the first conclusive evidence that silencing caffeic acid O-methyl transferase (COMT) in maize [13,14], alfalfa [15], tobacco [16] or poplar [17,18] plants affects the frequency of S lignin units and induces the incorporation of 5-hydroxyconiferyl alcohol into lignins as 5-hydroxyguaiacyl (5-OH G) units [13] involved in the unusual benzodioxane bonding pattern [18].
Although the pathway leading to lignins has been studied for decades [1], the monolignol biosynthesis scheme, once outlined as a metabolic grid, has been substantially reassessed over the past few years [4,6,19]. It has become apparent that the hydroxylation/methylation steps allowing the conversion of G to S moieties did not occur at the level of free hydroxycinnamic acids, but involved their corresponding aldehydes [19–21] and alcohols [22–25]. These results were obtained by in vitro studies of enzyme substrate specificities and by in vivo lignin labelling studies.
To further clarify the pathway leading to sinapyl alcohol in angiosperms, we have studied the lignification of wild type (WT) poplar plantlets and of transgenic plantlets with a reduced COMT activity. This study was carried out by analysis of stem or root samples from a few week-old plantlets by thioacidolysis and then determining the lignin-derived G and S monomers by GC-MS analysis of their trimethylsilylated derivatives (Fig. 1). At the onset of the in vitro culture on solid medium, a series of microcuttings were fed with coniferin or syringin, deuterated at their methoxyl groups and incorporated into the culture medium. These glucosides, which are the storage and the more putatively transport forms of the monolignols, were synthesized according to a previously reported procedure [26]. The incorporation of aglycones, namely coniferyl and sinapyl alcohols, into lignins was evaluated by tracing out the deuterium labelling in the G and S thioacidolysis monomers recovered specifically from G and S lignin units involved in β-O-4 bonds.
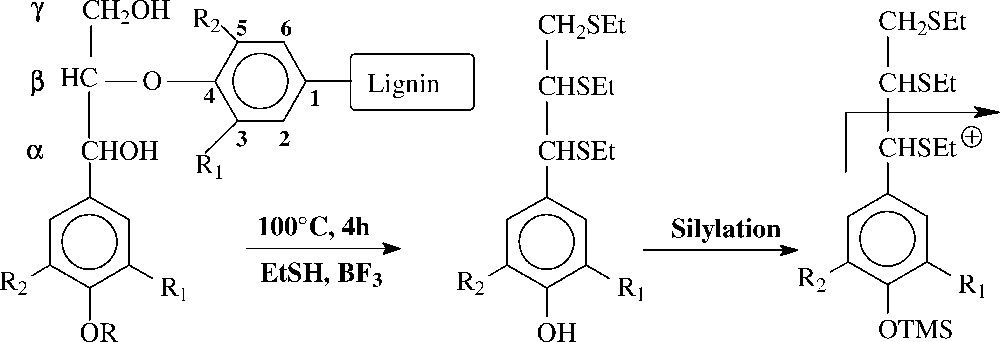
Principle of the thioacidolysis analytical degradation applied to lignins from unlabeled poplar plantlets and from poplar plantlets fed with deuterium-labelled lignin precursors. Thioethylated guaiacyl G (R1=OMe, R2=H) and syringyl S (R1=R2=OMe) monomers are specifically released from β-O-4 lignin structures (R=H or side chain of lignin unit) and determined by GC-MS of their trimethylsilylated (TMS) derivatives, using ion chromatograms reconstructed at their specific base peaks ions (benzylic ions resulting from the Ca–Cb cleavage). These base peak ions are observed at m/z 269 and 299 for unlabelled G and S TMS monomers, at m/z 272 and 302 for G and S TMS monomers with one OCD3 methoxyl groups and at m/z 305 for S TMS monomers with two OCD3 methoxyl groups.
2 Material and methods
2.1 Chemicals
Trideuterated vanillin and hexadeuterated syringaldehyde were prepared respectively by monomethylation of 3,4-dihydroxybenzaldehyde [27] and copper assisted methoxy substitution of 3,5-dibromo-4-hydroxy-benzaldehyde [28].
Trideuterated coniferin and hexadeuterated syringin were prepared as previously reported [26].
Trideuterated coniferin spectrometric data: 1H NMR (CD3OD): (d, 1 H, Harom2, ), 7.01 (d, 1, Harom5, ), 6.87 (dd, 1 H, Harom6, ; 2 Hz), 6.54 (d, 1 H, ArCHCH, ), 6.25 (dt, 1 H, ArCHCH, ; 5.6 Hz), 4.86 (d, 1 H, H1, ), 4.08 (d, 2 H, ArCHCHCH2, ), 3.75 (m, 1 H, H6b), 3.58 (m, 1 H, H6a), 3.27–3.43 (m, 4 H, H2,3,4,5). MS (EI, 70 eV) (%): 183 (100), 140 (79), 131 (23), 127 (78), 119 (30), 109 (30), 107 (26), 105 (24), 103 (34), 92 (26), 91 (75), 89 (27), 85 (22), 81 (32), 79 (36), 78 (32), 77 (66), 73 (51), 71 (28), 69 (31), 63 (23), 60 (47), 57 (53), 56 (26), 55 (87). C16H19D3O8.(H2O)2: C: 50.24 (calcd 50.39), H: 6.68 (calcd 6.87).
Hexadeuterated syringin spectrometric data: 1H NMR (DMSO, d6): (s, 2 H, Harom2,6), 6.45 (d, 1 H, ArCHCH, ), 6.30 (dt, 1 H, ArCHCH, ; 4.9 Hz), 4.90 (d, 1 H, H1, ), 4.10 (d, 2 H, ArCHCHCH2, ), 3.58 (m, 1 H, H6b), 3.40 (m, 1 H, H6a), 3.00–3.25 (m, 4 H, H2,3,4,5). MS (EI) : 217 (33), 216 (100), 188 (22), 187 (10), 173 (27), 160 (17), 152 (12), 73 (11), 60 (11); HRMS: calcd: 379.1875, found 379.1900.
2.2 Plant material and administration of deuterium-labelled lignin precursors
2.2.1 Plant labelling and culture
About 200 microcuttings from a wild-type hybrid poplar (Populus alba × Populus tremula, clone 717-1-B4) and from a COMT-deficient line ASOMT2B (COMT residual activity = about 5% of the control level on stem extracts of greenhouse-grown poplars) [17] were cultured aseptically on solid medium, according to a previously described procedure [17,29]. Half of the samples were deuterium-labelled as follows: after the sterile deposition of a shoot tip into culture tubes, 2.5 mg of coniferin trideuterated on its methoxyl group were solubilized in 30 μl dimethylsulfoxide and the solution was then deposited on the surface of the agar medium without direct contact with the shoot tip. The unlabelled and deuterium-fed microcuttings were then cultured in the growth chamber during 2 to 16 weeks (, 75% humidity, 16 h daytime, 10 μmol m−2 s−1 luminosity at leaf level from strip lighting). Another series of deuterium-labelling experiments was run with WT plantlets fed either with trideuterated coniferin or hexadeuterated syringin. The deuterated precursors were administered either one time at the onset of the culture, or three times (at the onset, and after 1 and 4 weeks of in vitro culture).
2.2.2 Sampling procedure
Five plantlets were collected every two weeks for the unlabelled and labelled WT or ASOMT2B lines. Within each series, the five new stems formed from the axillary bud of the microcutting were gathered for lignin analysis. Such a sampling procedure was also done for the roots. All the poplar samples were freeze-dried (total amount varying between 0.5 and 50 mg) and directly subjected to thioacidolysis for lignin evaluation.
2.3 GC-MS analysis of lignin-derived thioacidolysis monomers
Root or stem samples were washed with methanol, dried and then subjected to thioacidolysis as previously described [12]. The lignin-derived thioacidolysis monomers were trimethylsilylated and injected for GC-MS analysis on a Varian Saturn IV (Varian, Palo Alto, USA) ion trap mass spectrometer using the conditions previously described [9].
2.4 Histochemistry
Hand sections of 16 week-old plantlets, displaying about a dozen of internodes and fully developed leaves, were made using a razor blade. To detect native lignins from their cinnamaldehyde end-groups, these sections were treated with phloroglucinol, 2% (w/v) in ethanol, mixed 2:1 with concentrated hydrochloric, which revealed lignified areas from their purple colour [30]. To detect syringyl units in lignins, the Mäule staining was carried out by immersing the sections in a 1% (w/v) water solution of potassium permanganate for 5 min. The sections were then thoroughly rinsed in distilled water and treated with 3% hydrochloric acid until the dark colour faded, after which concentrated ammonium hydroxide was added to the sections to develop a magenta colour in syringyl lignified tissues [30]. These sections were done at the top (penultimate internode from the top), at the middle (fifth internode from the top) and at the base (tenth internode from the top) of the stem plantlets.
3 Results and discussion
3.1 Histochemical observation of lignins in plantlets
To visualise the lignified tissues in 16 week-old plantlet stems, cross sections of top, middle and bottom internodes were treated with the Wiesner (phloroglucinol–HCl) and the Maüle reagents. Phloroglucinol staining has long been utilized for localizing lignins in plant tissue sections [30]. It stains lignins a purple coloration deriving from reactions of hydroxycinnamyl aldehyde end-groups that are systematically present in native lignins, albeit in low amounts. The Maüle reagent specifically stains bright red the S lignin units. In agreement with literature data [31], the stems regenerated from the bud displayed a fully developed vascular cambium at the basal and middle parts of the 16 week-old plantlets whereas the top internode only displayed primary vascular bundles.
When the middle stem cross-sections were subjected to the phloroglucinol–HCl staining, the xylem of the control plants stained more importantly than that of the ASOMT2B ones (Fig. 2). In addition, in the top internodes provided only with primary vascular bundles, the WT sample stained red purple, which is diagnostic for lignified cell walls. In contrast, the vascular bundles of the ASOMT2B top sections did not stain, which suggests that they are not lignified or not lignified enough for a positive Wiesner coloration. Not unexpectedly, lignin in the secondary xylem of the basal and middle stem cross-sections of the WT line exhibited the positive magenta coloration diagnostic of S units while the analogous ASOMT2B sections showed a reduced staining, indicating a reduction in S lignin units (Fig. 2), which is consistent with the genetically-reduced COMT activity [17]. Taken together, these observations suggest that the COMT reduced activity not only decrease the frequency of S lignin units, but also induces some delay in lignification.
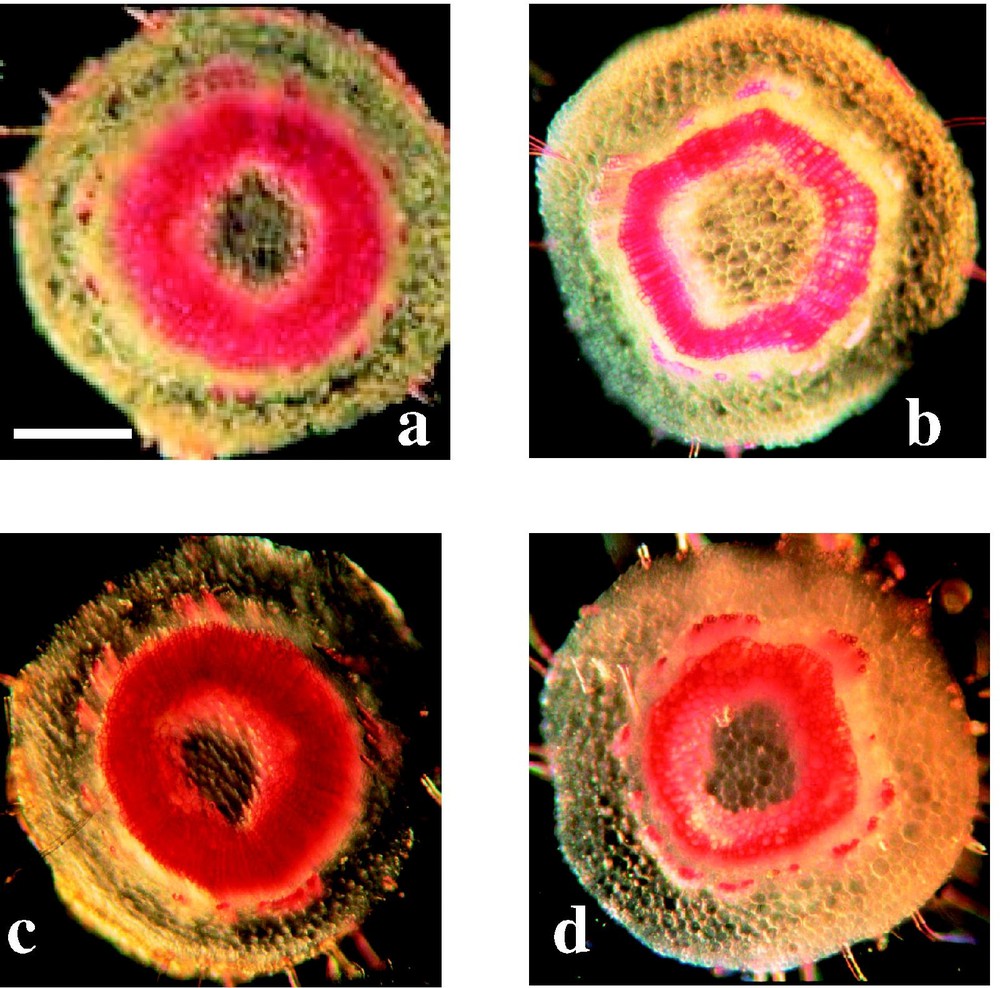
Histochemical staining of stem cross-sections from WT (a and c) and ASOMT2B (b and d) 16-week old plantlets (fifth internode from the top) with Wiener (a and b) and Maüle (c and d) reagents (white bar: 0.5 mm): relative to the control left sections, the ASOMT2B right sections showed reduced Wiener and Maüle xylem staining, diagnostic for a reduction in the lignin level and in the frequency of S lignin units, respectively.
3.2 Effective partial incorporation of deuterated precursors into stem and root lignins
The deuterated coniferin and syringin used for the set of experiments described herein were prepared as previously described in details [26]. The employed method is based on the coupling of a glucosidation and a Wittig reaction, leading to -unsaturated aldehydes, then easily reduced to the alcohols [32].
The coniferin and syringin deuterated at their methoxyl groups carried three or six deuterons, respectively. These deuterium atoms on methoxyl groups are not prone to exchange during lignin biosynthesis. Their incorporation into G or S lignin units was monitored by GC-MS examination of the base peaks of the mass spectra of G and S thioacidolysis monomers. In the unlabelled G and S monomers recovered from WT and ASOMT2B samples not fed with deuterated precursors, the base peaks were respectively observed at 269 and 299, which corresponds to the benzylic ions G-CHSEt+ and S-CHSEt+ (with G and S = trimethylsilylated guaiacyl and syringyl rings). The natural isotopic patterns for these unlabeled G and S ions were repeatedly found to give to M ratio lower than 2% while the to M ratio of the S compound was found close to zero. Accordingly, the incorporation of tri- or hexa-deuterated G and S rings into the lignins of deuterium fed plantlets could be traced out from abnormally high or signals.
When added to the solid culture medium, deuterated coniferin was massively used for the biosynthesis of root lignins, as revealed by the recovery of high deuterium level in thioacidolysis monomers released from the roots of WT and ASOMT2B plantlets (Table 1). In contrast, stem lignins were found to be deuterium-labelled to a much lower extent. This result establishes that coniferin diffuses into the gelose medium, is absorbed by the roots and then migrates essentially to the lignifying root cell walls where its aglycone is used for lignin biosynthesis. The administration way proposed in this study seems therefore more adapted to the study of root lignification. The deuterium labelling of root lignins could be evidenced not only in G units but also, more markedly, in S units. Deuterium incorporation into root lignins increases between the fourth and the tenth week after the beginning of the plantlet culture. In the following weeks, this deuterium labelling of root lignins was found to decrease. In other words, the effective use of the deuterated coniferin added to the medium lasted at least over a 10-week-long period. COMT deficiency in the ASOMT2B line did not seem to impede the use of deuterated coniferin. The only visible difference between the WT and transgenic samples was the higher deuterium level in the roots of the 4 and 16 week-old plantlets.
Thioacidolysis evaluation of deuterium incorporation into G and S lignin units of poplar WT (717 1B4) and ASOMT2B plantlets fed with [3-OCD3] coniferin (2.5 mg dissolved in 30 μl dimethylsulfoxide and then deposited on the surface of the culture medium at the beginning of the in vitro culture)
| G labellinga | ||||
| Age (weeks) | Root WT | Root ASOMT2B | Stem WT | Stem ASOMT2B |
| 4 | 29 | 45 | 7 | 4 |
| 6 | 42 | 52 | 3 | 5 |
| 10 | 53 | 56 | 3 | 3 |
| 12 | 44 | 40 | 3 | 3 |
| 16 | 23 | 32 | 3 | 3 |
| S labellingb | ||||
| Age (weeks) | Root WT | Root ASOMT2B | Stem WT | Stem ASOMT2B |
| 4 | 61 | 75 | 13 | 6 |
| 6 | 71 | 72 | 3 | 6 |
| 10 | 75 | 79 | 3 | 3 |
| 12 | 65 | 67 | 3 | 3 |
| 16 | 34 | 53 | 3 | 3 |
a Area percentage of signals on reconstructed ion chromatograms.
b Area percentage of signals on reconstructed ion chromatograms.
The syringin incorporation was performed in parallel to another coniferin feeding, using a different administration procedure (addition of the precursors at 0, 1 and 4 weeks after the beginning of the in vitro culture). No major difference, but a slightly higher deuterium concentration was observed when trideuterated coniferin was fed in three portions, delayed by few weeks. The highest deuterium incorporation in stem lignins was obtained with three administrations of [3,5-OCD3]-syringin, with which hexadeuterated S units reach 23% (Table 2). Different explanations can account for this result. Syringin may enter or migrate more easily than coniferin. In addition, since the deuterium is found only in S lignin units (Table 2), it means that is not divided between G and S units, contrarily to the deuterium of coniferin.
Thioacidolysis evaluation of deuterium incorporation into G and S lignin units of poplar WT (717 1B4) fed with [3-OCD3] coniferin or with [3,5-OCD3] syringin (2.5 mg dissolved in 30 μl dimethylsulfoxide and then deposited on the surface of the culture medium). The administration of the deuterated precursors was done one time at the beginning of the in vitro culture or repeated 3 times at the onset and after 1 and 4 weeks of in vitro culture
| [3-OCD3] coniferin | ||||
| Age in weeks (feeding number) | Root G labellinga | Root S labellingb | Stem G labellingd | Stem S labellingb |
| 7 (1) | 48 | 84 | 4 | 5 |
| 7 (3) | 62 | 87 | 4 | 5 |
| 9 (3) | 76 | 91 | 4 | 5 |
| [3,5-OCD3] syringinc | ||||
| Age in weeks (feeding number) | Root G labellinga | Root S labellingd | Stem G labellingd | Stem S labellingd |
| 7 (1) | 2e | 56 | 2e | 4 |
| 7 (3) | 2e | 60 | 2e | 23 |
| 9 (3) | 2e | 76 | 2e | 11 |
a Area percentage of signals on reconstructed ion chromatograms.
b Area percentage of signals on reconstructed ion chromatograms.
c The occurrence of S lignin units with only one deuterated methoxyl group was negligible as revealed by the percentage similar to that of unlabeled samples.
d Area percentage of signals on reconstructed ion chromatograms.
e Values similar to the percentage obtained for unlabeled samples.
Taken together, the data of Tables 1 and 2 reveal that the deuterium of syringin was exclusively directed to syringyl lignin units, whereas the deuterium of coniferin could be evidenced not only in G lignin units, but also and more markedly in S lignin units. In agreement with recent studies [22–24], this result further establishes that the hydroxylation/methylation steps leading to the formation of the S aromatic ring may occur at the alcohol stage and not only at the aldehyde stage [21].
3.3 Lignification of WT and COMT deficient poplar plantlets
The total yield and relative ratio of the G and S thioacidolysis monomers reflects the total lignin amount and the relative frequency of the G and S lignin units (Table 3), without interference from non-lignin components. For all plantlet roots, the (S + G) yield stabilized around 100 μmol per dry weight gram. As the recovery yield of thioacidolysis monomers released from 1 g of native poplar lignins varies between 1000 and 2000 μmol per dry weight gram [2,12], we may roughly estimate the root lignin content of the poplar plantlets varying between 5 and 10% (weight percentage). This approximation reveals that plantlet roots are substantially lignified. In addition, it should be kept in mind that the S and G thioacidolysis monomers may originate not only from lignin, but also from the aromatic domain of suberin, as both lignin and suberin have been reported to be important components of the root apoplastic barriers [33]. A fairly stable S/G thioacidolysis ratio, close to 0.4, was observed for all the root plantlet samples, whatever the line. That WT and ASOMT2B root samples yield similar (S + G) and S/G thioacidolysis values indicates that COMT deficiency has no impact on root lignification of in vitro grown poplar plantlets. This observation could be related to the lower fact that COMT is less repressed in the root than in the stem of the ASOMT2B line (Jouanin, unpublished results). The most important result revealed by the kinetic monitoring of lignification in the unlabelled plantlets and in the plantlets fed with deuterated coniferin or syringin is the following one. While the effective and massive incorporation of deuterium in G and S root lignin units could be assessed, this incorporation had no obvious impact on the lignin content and structure that were found to be very similar in the unlabelled and in the labelled samples. In other words, plantlets used exogenous material to synthesize root lignins, without any modification of lignin content or S/G ratio. When deuterated coniferin was fed to the plantlet, the S units of root lignins were substantially more deuterated than the G units (Table 1). The conversion of G rings into S rings at the stage of p-OH cinnamyl alcohols therefore allowed the plantlet to redirect the G-type lignin precursor towards the synthesis of S lignin units so that the S/G ratio is maintained at a genetically-programmed value in the unlabelled or labelled samples. The lignin content and structure seem therefore to be under the control of a tight regulation mechanism.
Thioacidolysis monitoring of lignification in root and stem of poplar WT (717 1B4) and ASOMT2B plantlets, without and with administration of [3-OCD3] coniferin (2.5 mg dissolved in 30 μl dimethylsulfoxide and then deposited on the surface of the culture medium at the beginning of the in vitro culture). The total yield S+G (expressed in μmol per dry weight gram) and molar ratio S/G (values between brackets) of the thioacidolysis S and G monomers give an estimate of lignin content and structure
| Roots | ||||
| Age (weeks) | WT | WT fed with coniferin | ASOMT2B | ASOMT2B fed with coniferin |
| 6 | 68 (0.47) | 79 (0.37) | 80 (0.33) | 108 (0.49) |
| 10 | 106 (0.39) | 108 (0.43) | 110 (0.37) | 87 (0.47) |
| 12 | 102 (0.45) | 107 (0.41) | – | 123 (0.43) |
| Stems | ||||
| 6 | 68 (0.47) | 57 (0.41) | 52 (0.23) | 48 (0.23) |
| 10 | 162 (0.62) | 159 (0.66) | 140 (0.59) | 100 (0.52) |
| 12 | 149 (0.79) | 180 (0.79) | 133 (0.52) | 120 (0.52) |
In contrast to root lignification and in agreement with a wealth of literature data [17,18,34], stem lignification was found to be affected by COMT deficiency. The total thioacidolysis yields were systematically found to be lower in the ASOMT2B stems than in the WT analogues (Table 3). The most likely explanation for these lower thioacidolysis yields and in agreement with the aforementioned reduced Wiesner staining is that the ASOMT2B stems are less lignified than their WT counterparts. In addition, and in agreement with the reduced Maüle staining, the ASOMT2B stem lignins displayed a reduced frequency of S lignin units when compared to the control. This result confirms that the biosynthesis of S lignin units is affected by COMT deficiency at the very early stages of lignification in growth-chamber-cultivated plantlets regenerated from microcuttings. The 5-OH guaiacyl (5-OH G) units which are the diagnostic signature of COMT deficiency were found in trace amounts before the 12th week of in vitro culture. It was only in the 16 week-old plantlets that the frequency of these units slightly increased relative to the control value (data not shown).
4 Conclusion
The labelling protocol used for this study seems to be suitable for tracing the metabolism of lignin precursors at least in root tissues, without handling radioactive materials or wounding the plant. The employed deuterium labelling does not affect the growth parameters of the plantlets. Due to the high sensitivity of the thioacidolysis method, it is possible to monitor lignification in minute amount of plantlet roots and stems. GC-MS analysis proved that trideuterated coniferin and hexadeuterated syringin were incorporated mainly in root lignins and, to a lower extent, in stem lignins without major effect on plantlet growth and lignin profile. Deuterium from coniferin was recovered in G and S lignin units, whereas deuterium from syringin was only found in S units, which further establishes that the conversion of G to S lignin precursors may occur at the level of p-OH cinnamyl alcohols.
Acknowledgments
We wish to thank Laurence Piumi and Charlette Franscesch for their contribution to the synthesis and administration of the deuterated precursors as well as Valérie Ferret and Véronique de Nadaï for the in vitro cultures. We would also like to sincerely thank Dr K. Fukushima for his kind help in histochemistry and for valuable discussions. This work was supported by a grant from the French ‘Ministère de l’Éducation nationale, de l'Enseignement supérieur et de la Recherche' (ACCSV-14).