

Abridged English version
The role of the landscape in the distribution of the animal populations has still been relatively little studied and largely depends on the biology of the species, particularly in relation to their diet (specialist or opportunist) and their moving capacities as well as the landscape variables taken into account.
The grey heron (Ardea cinerea) occurred throughout France at the beginning of 19th century, but was brought almost to extinction at the beginning of the following century. Only two significant breeding colonies remained in eastern and western France. From these two refuges, the species showed a very strong numerical and geographical expansion, involving a progressive recovering of its initial distribution. However this varied according to the area, and both the direct action of man and the structure of the hydrographical network probably played a role in this recovering.
The aim of this study is to analyse the role played by the hydrographical network structure in the process of grey heron recovery in France, and to determine whether there were differences in local strategies of spatial use, both between and within the two historical refuges where populations seem nowadays largely saturated. The results distinguished four groups of breeding colonies according to the hydrographical structure at local and regional scales.
Eastern France was homogeneous for the elements of the hydrographical network that influenced the distribution of the colonies. For these colonies, clustered in group I, the presence of zones of permanent open water was predominant, particularly when large portions of rivers and ponds were present.
In contrast to the East, the situations of group II and III of western France were very different in spite of the fact that the population of group III was derived from that of group II.
For group II, the importance of large marshes with their ponds and their significant network of small ditches played the major role in the distribution of colonies, followed by the broad portions of rivers, while for group III, the length of the coastline seemed the most significant factor in the choice of a breeding site, with the intertidal area and more particularly of mudflats.
This work shows the importance of the type of major aquatic environments (small channels, marshes, large rivers) and especially the differences existing between areas, even when they are close as in western France.
In addition to the specific structures of landscape of each area, the regional history of the populations of herons could influence the strategies of breeding colonies distribution, insofar as the young birds tend to favour the type of environment in which they were born, which leads to two very different situations between eastern and western France. In the West, the ‘mother’ colony at Grand-Lieu for a long time led to the installation of ‘daughter’ colonies in willow marshes, before the new colony of Guérande in the Loire-Atlantique (1966) generated in a second time a line of coastal colonies settling in conifer trees near the coast as far as Finistère. However, with group IV and the colony in the Loire-Atlantique present in group I, started a progressive colonisation of the inland environments, a strategy only observed in eastern France. In this area, the species remained faithful to the choice of small colonies in large trees in woods and in the grounds of chateaux.
The relation between the size of the colonies and the structure of the landscape needs more investigations, particularly by taking into account the fitting of the various elements of the hydrographical network within the landscape, and by discriminating the water depth, this species exploiting only shallow water.
1 Introduction
Si le héron cendré (Ardea cinerea) était présent sur l'ensemble du territoire français au début du XIXe siècle, des décennies de persécution à partir de 1870 conduisirent l'espèce à une quasi extinction au début du siècle suivant. Seules deux colonies importantes subsistaient, la première protégée depuis des siècles dans le parc du château d'Écury-le-Grand, dans l'Est de la France, la seconde apparue dans l'Ouest, dans les forêts flottantes de saules du lac de Grand-Lieu, difficilement accessibles pour l'homme, apparition favorisée par l'arrêt de la chasse pendant la première guerre mondiale [1]. À partir de ces deux zones refuges, l'expansion a été très lente jusque dans les années 1960, l'espèce n'étant plus considérée comme nuisible qu'à partir de 1968. Mais ce n'est qu'à partir de 1974, date de sa protection intégrale, que l'espèce a connu une très forte expansion numérique et géographique [2], entraînant une recolonisation progressive de son aire de répartition initiale (Fig. 1). Cependant, celle-ci s'est faite de manière variable selon les régions et, parallèlement à l'action directe de l'homme, la structure du réseau hydrographique a probablement joué un rôle dans cette reconquête.
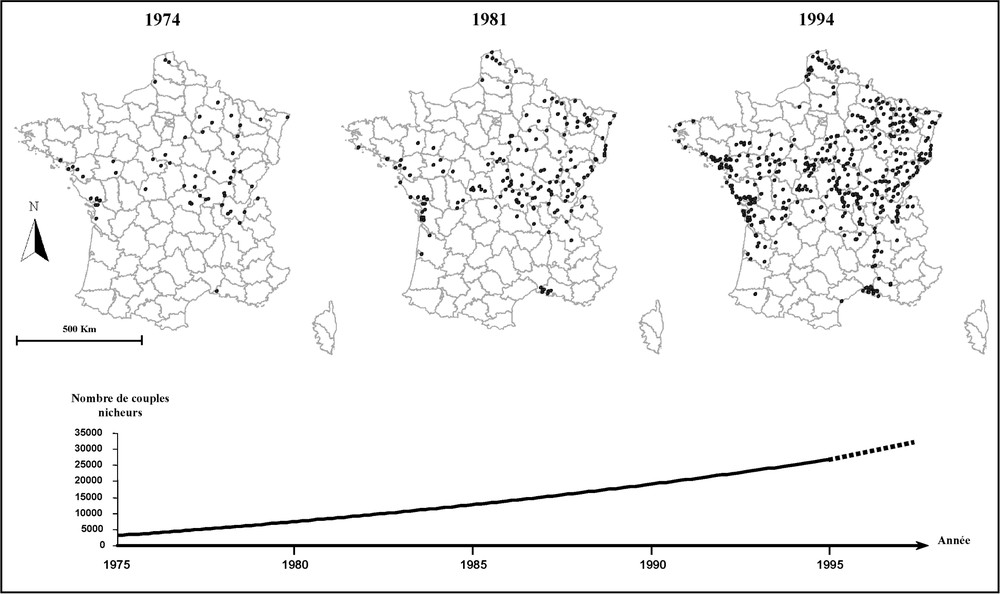
Évolution du nombre de colonies (haut) et de couples nicheurs (bas) de hérons cendrés depuis la protection intégrale de l'espèce en 1975 (d'après [1,6]).
Le rôle du paysage dans la distribution des populations animales est encore relativement peu étudié et varie fortement selon la biologie de l'espèce considérée [3], notamment selon son caractère spécialiste ou opportuniste et ses capacités de déplacement, ainsi qu'en fonction des paramètres du paysage pris en compte [4]. Par ailleurs, la dynamique des populations joue un rôle potentiellement important dans la mesure où les ressources naturelles, et par conséquent la répartition et la structure des habitats, interviennent dans la vitesse de limitation des populations et donc leur capacité à fournir des émigrants permettant l'expansion géographique [5–8]. Cependant, ces éléments locaux sont rarement pris en compte dans la littérature, ne serait-ce que parce qu'on considère généralement qu'une espèce répond de manière univoque aux critères du paysages sensés participer à sa niche écologique.
L'objet de la présente étude est de tester cette dernière hypothèse, en analysant, dans le cadre conceptuel de l'écologie du paysage [9], le rôle joué par la structure du réseau hydrographique dans le processus de reconquête de la France par le héron cendré, et de voir s'il existe réellement des différences de stratégies locales d'occupation de l'espace, notamment en fonction de la localisation des colonies refuges, et du degré de saturation des populations régionales.
2 Matériels et méthodes
2.1 Données sur le héron cendré
L'inventaire national des héronnières de France de 1994, réalisé par L. Marion (université Rennes-1, MNHN, ministère chargé de l'Environnement), avec l'aide du réseau d'observateurs locaux [6], a servi de données de base pour la localisation des colonies (Fig. 2) et la quantification de leurs effectifs.
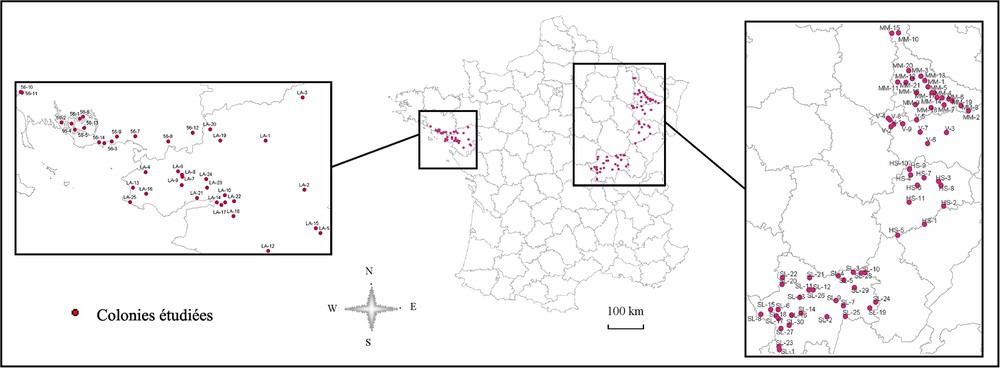
Répartition des colonies en 1994 dans les zones tests témoins.
La méthode du recensement de 1994 a été identique à celle utilisée lors des précédents recensements nationaux de 1974, 1981, 1985 et 1989 [6]. En principe, tous les nids occupés pendant la reproduction de 1994 ont été recensés entre mai et juillet. On peut considérer que ce recensement a été quasi exhaustif dans les régions retenues pour le présent travail, et que la distribution des colonies reproduit fidèlement la réalité de l'occupation du milieu par les oiseaux.
2.2 Aires d'étude
Lors de cette étude méthodologique, nous avons volontairement privilégié des grandes régions témoins à l'Ouest et à l'Est de la France, pionnières dans la colonisation du territoire français par cette espèce à la suite de sa quasi-disparition au début du XXe siècle. À l'ouest, nous nous sommes focalisés sur les départements de Loire-Atlantique et du Morbihan, et à l'est sur les départements de Meurthe-et-Moselle, Saône-et-Loire, Haute-Saône et des Vosges (Fig. 2), régions qui tiennent une place prépondérante dans les effectifs nationaux en France [6], pays qui abrite par ailleurs la plus grande population de hérons cendrés en Europe [2].
2.3 Données environnementales
Nous avons pris en compte différents descripteurs du réseau hydrographique, notamment en distinguant le réseau linéaire et le réseau surfacique. Le réseau linéaire correspond à tous les écoulements d'eaux permanents et intermittents, alors que le surfacique correspond aux étangs, marais et zones intertidales. Pour l'étude, 97 194 km de réseau linéaire et 280 633 ha d'éléments surfaciques ont été analysés. Ces données ont été extraites de BD-Carthage version 1.1, qui est un fichier numérique conçu conjointement par le ministère de l'Environnement et l'Institut géographique national (IGN), qui représente tout le réseau hydrographique, à la fois linéaire et surfacique. Elle décrit la nature (naturelle, canal, douve, estuaire), l'état (permanent ou temporaire) et la largeur des rivières. Elle décrit également les plans d'eau par leur nature (permanent d'eau douce, temporaire d'eau douce, permanent d'eau de mer et temporaire d'eau de mer) et leur type (marais, estran...). Cette base de donnée permet d'obtenir les mesures de longueur et de surface du réseau hydrographique selon les différents types que nous venons de définir. Cependant, du fait de certaines lacunes de cette base de données dans quelques-unes des parties de la zone d'étude, il a été nécessaire de modifier ce fichier en utilisant des jeux de données supplémentaires pour mieux correspondre aux besoins supposés des hérons. En effet, au niveau des marais salants et des grands estuaires, le réseau très fin de douves n'a pas été pris en compte lors de la réalisation de BD-Carthage, alors qu'il semble avoir une grande importance pour les hérons qui y sont souvent observés. Nous les avons donc numérisé en se basant sur les cartes au 1/25 000 de l'IGN. De même, nous avons dû faire deux classes de largeur de cours d'eau (0–5 et 6–15 m) au lieu de la classe unique de 0–15 m de BD-Carthage, insuffisamment discriminante. Ceci a été également effectué en se basant sur les cartes de l'IGN.
2.4 Traitement des données
Les données caractérisant la position des colonies et le réseau hydrographique ont été intégrées dans Arcview 3.1, un système d'information géographique (SIG). Le SIG permet l'analyse simultanée et croisée des différentes couches d'information (c'est-à-dire réseau hydrographique et position des colonies).
Nous avons déterminé des zones circulaires (buffers) de différents diamètres [10] autour de chaque colonie, afin de tester la zone d'influence optimale de chaque colonie et de mesurer le rôle de la structure du paysage dans le positionnement et la taille des colonies. Une étude préalable [10] a montré que les buffers tenant compte à la fois de la présence de colonies voisines et de l'effectif de chacune sont ceux qui représentent au mieux les surfaces réellement exploitées par les colonies. Nous avons retenu deux rayons prenant en compte les colonies voisines et les effectifs de chacune d'entre elles pour la suite de l'étude. Le réseau hydrographique linéaire et zonal en a ensuite été extrait, en utilisant la capacité du logiciel de calculer pour chaque colonie, et pour chaque rayon, la longueur de trait de côte, la longueur totale de rives des différents types de réseau hydrographique que nous avons défini ainsi que leur surface en eau, qui se trouve à proximité des colonies.
Une analyse en composantes principales (ACP) a été réalisée sur ces variables (Tableau 1) afin de regrouper les colonies qui se ressemblent le plus du point de vue de la structure du réseau hydrographique qui les entoure. La méthode de l'UPGMA a été utilisée, car elle regroupe les colonies en maximisant les ressemblances. Les analyses et les graphiques ont été effectués avec le logiciel ADE-4 [11].
Descriptif de la base de donnée BD-Carthage décrivant le réseau hydrographique linéaire et surfacique
| Type | Différents champs de description | Commentaires |
| Linéaire (Hya dans BD-Carthage) | État (données d'origine de BD-Carthage) | 1 – écoulements permanents |
| 2 – écoulements intermittents | ||
| Largeur (données modifiées) | 1 – de 0 à 5 mètres | |
| 2 – entre 16 et 50 mètres | ||
| 3 – plus de 50 mètres | ||
| 4 – entre 6 et 15 mètres | ||
| Nature (données d'origine de BD-Carthage) | 1 – cours d'eau à berges non maçonnées | |
| 2 – cours d'eau à berges maçonnées | ||
| 3 – canal : voie navigable artificielle ou autre voie d'eau portant la désignation « canal » et d'une largeur supérieure à 15 m | ||
| 4 – aqueduc, conduite forcée : tuyau ou chenal artificiel conçu pour le transport de l'eau | ||
| 5 – fossé : chenal pour l'irrigation, le drainage, douves des marais salants | ||
| 6 – autre | ||
| 7 – estuaire principal : écoulement d'un cours d'eau dans l'estran | ||
| 8 – estuaire secondaire : écoulement (autre que celui d'un cours d'eau) dans l'estran | ||
| Surfacique (Hyz dans BD-Carthage) | Nature (données modifiées) | 10 – eau douce permanente |
| 11 – eau douce non permanente | ||
| 12 – eau salée permanente | ||
| 13 – eau salée non permanente | ||
| Type (données modifiées) | 101 – cours d'eau | |
| 102 – plan d'eau, bassin, réservoir | ||
| 103 – ensemble de petits plans d'eau | ||
| 104 – traitement des eaux, station de pompage | ||
| 105 – bassin portuaire fluvial | ||
| 111 – zone temporairement recouverte d'eau | ||
| 112 – sables et graviers dans le lit d'un cours d'eau | ||
| 121 – pleine mer | ||
| 122 – écoulement d'eau | ||
| 123 – nappe d'eau | ||
| 124 – bassin portuaire | ||
| 131 – marais salants | ||
| 132 – zone rocheuse | ||
| 133 – zone mixte rocher et sable | ||
| 134 – zone de sable humide | ||
| 135 – zone de vase | ||
| 136 – zone de graviers et galets |
Nous avons ensuite réalisé des corrélations entre les effectifs des colonies et ces différents paramètres de l'hydrographie pour identifier quels sont ceux qui sont importants pour le choix de l'emplacement de la colonie. La significativité de ces corrélations a été déterminée par la relation entre la loi de r et la loi du t de Student, qui permet d'utiliser la table du t :
3 Résultats
3.1 Comparaison des colonies
L'ACP (Fig. 3) a permis de distinguer quatre groupes de colonies. L'axe F1 traduit un gradient de continentalité. Le groupe I rassemble toutes les colonies de l'Est de la France, plus une de Loire-Atlantique. C'est un groupe que l'on peut qualifier de continental. À l'opposé de l'axe F1, on trouve les groupes II et III, qui rassemblent les colonies de Loire-Atlantique et du Morbihan qui se trouvent dans une situation littorale. Ces deux groupes s'individualisent selon l'axe F2, qui représente un gradient sud–nord, avec le groupe II dans une situation plutôt sud, composé essentiellement de colonies de Loire-Atlantique, et le groupe III dans une situation plutôt nord, composé surtout de colonies du Morbihan. Quant au groupe IV, comprenant uniquement quelques colonies de Loire-Atlantique, il se trouve dans une situation intermédiaire suivant l'axe F1, et tend vers une situation continentale. Le rôle des différents descripteurs de la structure du paysage hydrographique ne pourra pas être caractérisé pour ce groupe, car il comprend un nombre trop faible de colonies.
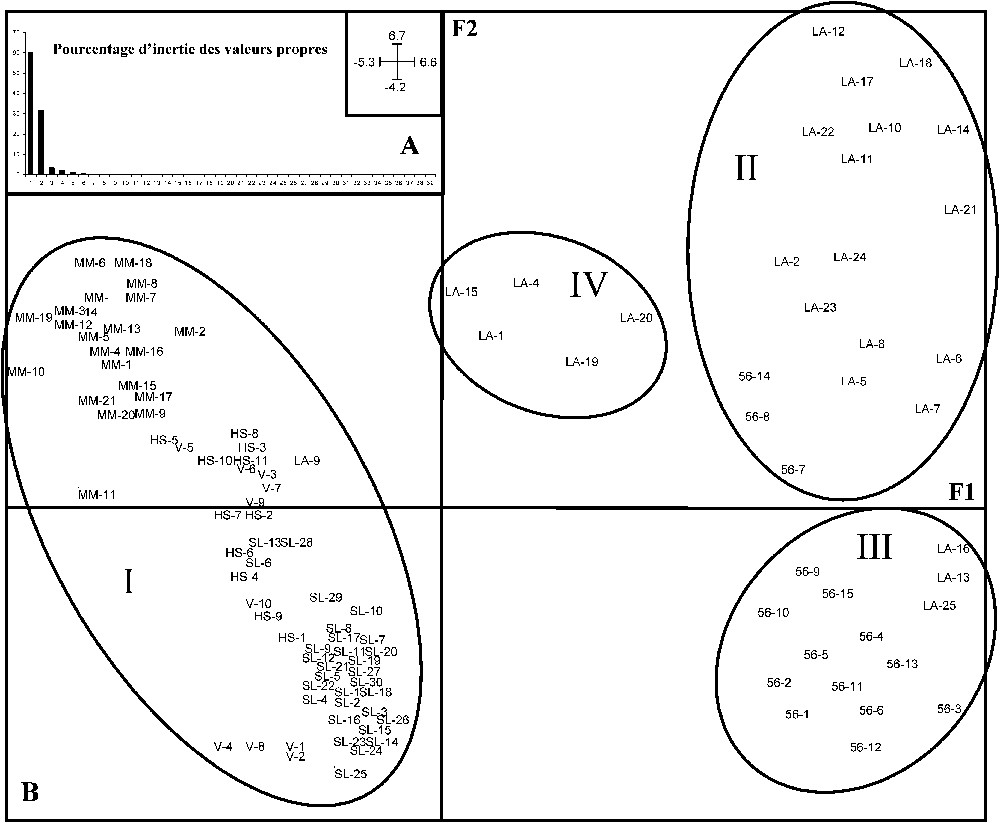
Représentation des axes F1 et F2 de l'ACP effectuée sur les variables caractérisant la structure du réseau hydrographique présent dans les alentours des colonies. A : Histogramme des valeurs propres. B : Projection des sites (LA-x pour la Loire-Atlantique ; 56-x pour le Morbihan ; MM-x pour la Meurthe-et-Moselle ; HS-x pour la Haute-Saône ; SL-x pour la Saône-et-Loire ; V-x pour les Vosges) et des ellipses (I, II, III, IV) distinguant les colonies selon l'UPGMA. Les coordonnées des axes sont dans le coin haut droit de la partie A de la figure.
3.2 Rôle des différents descripteurs de la structure du paysage hydrographique
Les situations entre le groupe II et le III de l'Ouest de la France sont très différentes, malgré le fait que la population de hérons du groupe III soit historiquement issue de celle du groupe II [12]. Contrairement à l'Ouest, la région Est est homogène quant aux éléments du réseau hydrographique qui influencent le positionnement des colonies, et a été analysée globalement.
3.2.1 Groupe I
Dans ce groupe uniquement représenté dans l'Est de la France (sauf une colonie), ce sont les zones en eau permanente (nature 10) et plus précisément les très larges portions de rivières (type 101) et des plans d'eau (type 102) qui sont prépondérants (Fig. 4-I).
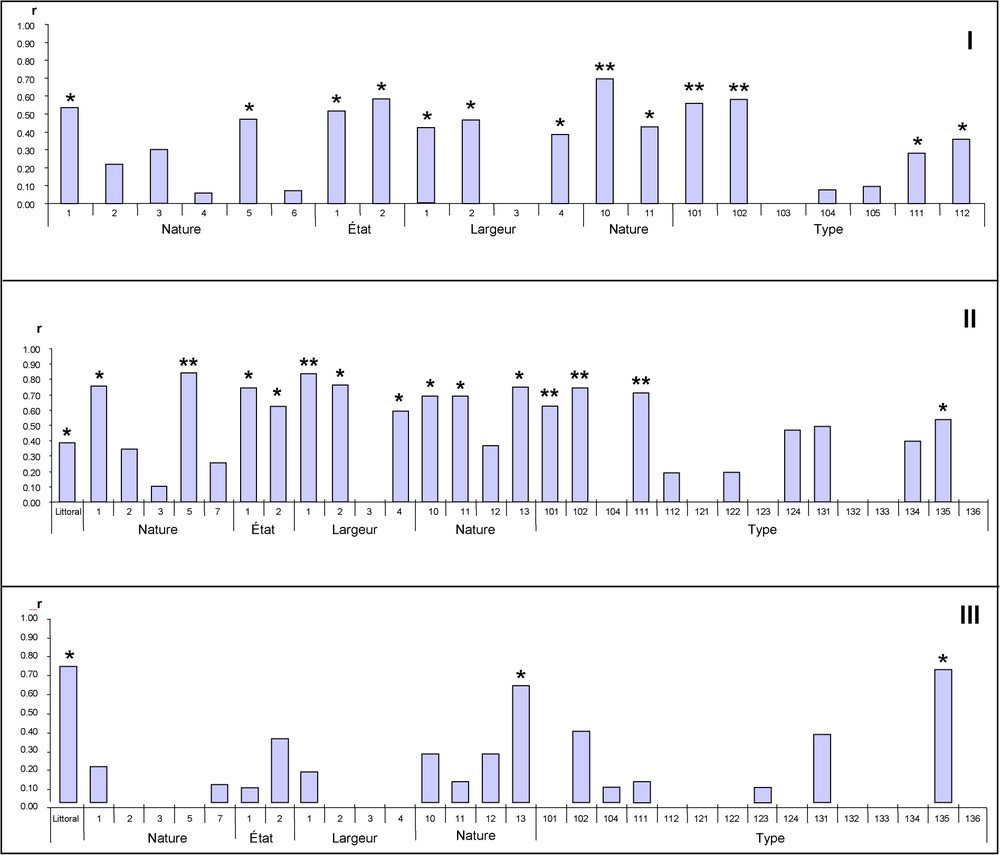
Niveau de corrélation (∗=p<0,05 ; ∗∗=p<0,01) pour chaque descripteur du réseau hydrographique entre la longueur de rives de ces descripteurs et l'effectif des colonies. (I) région Est, (II) Loire-Atlantique, (III) Morbihan. La classification des variables (Tableau 1) est celle de BD-Carthage (IGN).
3.2.2 Groupe II
Même si la majorité des descripteurs du réseau hydrographique semblent avoir un effet sur la localisation des colonies de ce groupe centré sur la Loire Atlantique (Fig. 4-II), l'importance des zones de marais (type 111) ressort le plus, avec leurs plans d'eau (type 102) et leur important réseau de petits canaux (nature 5, largeur 1). On observe également un rôle de première importance des très larges portions de rivières (type 101).
3.2.3 Groupe III
L'analyse en détail des colonies de ce groupe composé principalement de colonies du Morbihan montre que tous les descripteurs du réseau hydrographique qui répondent le mieux à la distribution des colonies diffèrent de ceux du groupe II (Fig. 4-III). La longueur de trait de côte (littoral) semble être le facteur le plus important dans le choix d'un site de nidification avec la surface des zones d'estran (nature 13), et plus particulièrement des parties vaseuses de celle-ci (type 135). On observe donc une réponse différente entre les deux populations de l'Ouest, l'une sélectionnant les eaux douces en Loire Atlantique et l'autre les eaux marines ou saumâtres dans le Morbihan, en dépit du fait que les deux départements disposent d'une façade maritime d'importance comparable, avec d'importants estrans, avec le Traict du Croisic et les vasières de la baie de Bourgneuf, qui ne sont pas ou que peu utilisées en Loire-Atlantique.
4 Discussion
Le héron cendré est une espèce coloniale qui se nourrit essentiellement sur des territoires alimentaires individuels [5,13] de proies essentiellement aquatiques pendant la période de reproduction [1,14,15]. L'emplacement de la colonie n'est donc pas choisi au hasard, les Ardéidés préférant les régions où les surfaces de zones humides ou le linéaire de rives sont élevées [1,16–19]. Les hérons cendrés localisent leur colonie généralement au centre de gravité de ces zones, excepté lorsque les perturbations humaines les contraignent à privilégier les sites inaccessibles à l'homme [1,7]. Ce choix n'est généralement pas lié à une meilleure connectivité des éléments favorables du paysage, contrairement à de nombreuses espèces [20,21], en raison des capacités volières du héron qui lui permettent de s'affranchir des obstacles, mais au coût énergétique du vol qui favorise l'installation des colonies au centre des zones alimentaires potentielles afin de minimiser les déplacements [1,5,7,13]. Au-delà des surfaces elles-mêmes, c'est surtout le linéaire de rives inclues dans l'aire alimentaire effective de la colonie qui paraissait jusqu'à présent déterminer la taille des colonies [1].
Le présent travail complète ces relations en montrant l'importance des milieux aquatiques prépondérants (petits canaux, marais, grands cours d'eau) et surtout les différences étonnantes existant entre régions et même entre départements voisins pour ce qui concerne l'Ouest de la France. La population de hérons de Loire-Atlantique (les colonies du groupe II) est très influencée dans son choix d'un site de nidification par la présence d'importantes zones de marais caractérisées par un maillage très dense de douves de petite taille et de plans d'eau. Ces éléments du paysage fournissent aux oiseaux des eaux peu profondes dans lesquelles ils peuvent facilement se nourrir. La partie estuarienne de la Loire joue secondairement un rôle très important pour une partie des colonies, qui y trouvent à la fois des milieux favorables pour s'alimenter, mais également des endroits isolés à l'abri de toute perturbation humaine pour établir les colonies.
Bien qu'issue de la population de Loire-Atlantique, celle du Morbihan (l'essentiel des colonies du groupe III) n'est pas influencée par les mêmes éléments du paysage. Les grandes zones humides dulçaquicoles largement représentées en Loire-Atlantique étant peu présentes dans le Morbihan, les nouveaux arrivants dans ce département ont utilisé des habitats qu'ils délaissent en Loire-Atlantique comme les estrans, d'où l'importance jouée dans le Morbihan par la longueur du trait de côte ainsi que la surface de vasières littorales devenus essentiels pour le développement des colonies dans ce secteur. Le plus étonnant est que les milieux intertidaux ou saumâtres soient devenus le milieu optimal chez cette population, qui délaisse les milieux dulçaquicoles. En fait, la conquête du Morbihan s'est probablement faite à partir de la colonie de Guérande, en Loire-Atlantique [1,12,22] qui avait été contrainte d'utiliser les conifères comme support de nids les plus sécurisants face à l'homme, en l'absence de marais inaccessibles sur la presqu'île Guérandaise, et qui a donné naissance à la lignée des colonies bretonnes littorales, cette colonie de Guérande exploitant essentiellement les marais salants et les vasières littorales du Traict du Croisic, milieu privilégié par la suite dans le Morbihan pour ces raisons historiques de colonisation.
Dans l'Est de la France, on ne retrouve pas cette hétérogénéité de réponse, malgré l'étendue de la zone étudiée, les rivières très larges, les plans d'eau et les zones inondables étant les éléments du paysage qui déterminent la taille et la position des colonies, sans exception. Ces habitats, voisins de ceux présents en Allemagne, expliquent sans doute en partie les différences de régime alimentaire observées dans les études de Schlegel [23] et de Lechner et Utschick [24] par rapport à l'Ouest de la France [1,15].
Au-delà des structures de paysage propres à chaque région, l'histoire régionale des populations de hérons influence d'une façon importante les stratégies de choix d'installation des colonies, dans la mesure où les jeunes oiseaux ont tendance à privilégier le type de milieu dans lequel ils sont nés [1], ce qui conduit à deux situations très différentes entre l'Ouest et l'Est de la France. Dans l'Ouest, la colonie mère de Grand-Lieu a longtemps fait privilégier chez ses colonies filles l'installation dans des saulaies ou aulnaies continentales inondables, puis est née une lignée de colonies côtières s'installant dans des résineux à proximité d'estrans, apparue à Guérande (cf. supra) et qui a généré une série de colonies côtières jusque dans le Finistère [1,12]. Cependant, avec le groupe IV et la colonie de Loire-Atlantique présente dans le groupe I, on observe une conquête récente des milieux continentaux par cette population qui se rapproche de ce fait des milieux qui sont utilisés dans l'Est de la France, où l'espèce est, en revanche, restée fidèle au choix de petites colonies dans les parcs et bois de feuillus. Il est évidemment encore trop tôt pour envisager une séparation écologique durable entre populations continentales et littorales chez le héron cendré, telle qu'elle existe par exemple chez la gorge bleue avec la sous-espèce Luscinia svecica namnetum propre aux marais saumâtres de la façade atlantique française [25,26], ou des deux sous-espèces de grand cormoran Phalacrocorax carbo carbo (marine) et Ph. c. sinensis (continentale), qui se reproduisent aussi dans cette région et qui montrent une différenciation génétique nette [27], contrairement à la gorge bleue [28].
La structure génétique des populations de hérons cendrés reste à étudier, et la relation entre la taille des colonies et la structure du paysage doit être approfondie, notamment en tenant compte de l'agencement des différents éléments du réseau hydrographique au sein du paysage, et en discriminant davantage la profondeur d'eau, car cette espèce n'exploite que des eaux de faibles profondeurs [1,16].
Remerciements
Cette étude a bénéficié des financements du ministère français chargé de l'Environnement (recensements nationaux et BD-Carthage) et de la SESLG. Nous tenons à remercier P. Marion, la LPO de Loire-Atlantique, Bretagne Vivante, le GOB, l'AOMSL, le GNFC, la LPO Lorraine et les services départementaux de l'ONCFS et de l'ONF en Lorraine, qui ont participé au recensement des héronnières coordonné au niveau national par L. Marion.