

1 Introduction
Deep-sea vent and seep animals communities have been thoroughly studied since they were discovered about 30 years ago. Initially, it was suggested that these animals were ‘living fossils’ [1], but combinations of fossil and molecular data indicate that most modern vent animal groups arose relatively recently [2]. Recent studies [3–6] underlined the ecological and zoological affinities of organisms associated with sunken woods with organisms associated with cold seeps, hydrothermal vents and whale falls. Indeed, the organic substrata of vegetal origin (wood, leaves, seeds, nuts) that accumulate in sedimentation basins or in estuaries, at depths beyond the penetration limit of sunlight, host an original but poorly studied fauna. Although insufficiently documented, it has been suggested that these communities, like hydrothermal or cold seep ones, would depend on chemoautotrophic bacteria [7].
The purpose of the present study is to discuss the ‘wooden steps to deep-sea vent’ hypothesis proposed by [3,4] for mytilids associated with deep-sea reducing environments, using datasets that include more species associated with sunken woods. These authors showed that all bivalve molluscs of the subfamily Bathymodiolinae [8] associated with seeps and vents, together with other mytilids associated with sunken woods and bones, form a monophyletic group and that within this clade one species, associated with sunken wood, had a basal position. From these results, they hypothesized that “decomposing wood and bone may have served as ‘steps’ for the introduction of mytilid taxa to vent and seeps.” However, although the results of [4] suggested that Bathymodiolinae species associated with organic substrata must be included into the dataset in order to better understand the evolutionary origins of vents' fauna, more recent phylogenetic studies are always biased toward vent and seep species [9,10]. To reduce this taxonomic bias, we present here an analysis of the molecular diversity of mytilids associated with sunken woods sampled in the Solomon Islands.
As the taxonomy of sunken wood mussels is poorly known, we first delimited the terminal taxa using the diversity of the fragment of COI gene as suggested in the ‘Barcoding of Life’ project [11]. Second, we explored the 18S rDNA dataset to confirm that the sunken woods mytilids of the Solomon Islands belonged to the monophyletic group identified by [4]. Third, we needed to determine which was the best outgroup to use in order to analyze the relationships within the monophyletic group that includes the Bathymodiolinae. Indeed, the relationships between mytilid subfamilies were not completely resolved with the 18S rDNA [12] and are still under discussion. Indeed, Chichvarkhin [13], using several morphological analyses [14,15] and fossil records [16], have proposed alternative hypotheses to that proposed by [4,12]. Thus, to determine which outgroup to use to study the relationships within Bathymodiolinae, we added the phylogenetic analysis of mytilids using the COI gene. Finally, we analyzed the relationships within the monophyletic group that included the Bathymodiolinae, first by using the diversity of COI, then by adding, for each terminal taxon – identified with the COI gene –, a portion of the mitochondrial gene ND4 and a portion of the nuclear genes 18S and 28S rDNA, two genes used by [5,9].
2 Materials and methods
2.1 Sampling
Mytilids associated with sunken plant materials (wood, seeds, nuts, etc.) were collected at 100–1200-m depth during the Salomon 2 cruise with R/V Alis in October 2004. A list of stations and localities is available at www.tropicaldeepseabenthos.org. Based on the morphology of the shell, Rudo von Cosel grouped the specimens into four morphospecies. For each morphospecies, we used three to eight individuals in the molecular analyses (Table 1). Overall, 22 individuals covering the morphological diversity of the sampling, as well as the explored depth and geographical ranges, were analyzed.
List of specimens sampled during the Salomon2 cruise in the Solomon Islands and sequenced for the gene COI, ND4, 18S, and 28S genes
| Species | Morphotype/Phylotype | Specimen label | GenBank N° | Habitat | |||
| 18S | 28S | COI | ND4 | ||||
| Idas sp. | SAL-1 | Rva | DQ340795 | – | DQ340775 | – | sunken wood |
| RVe | – | – | DQ340778 | – | sunken wood | ||
| RVh | – | – | DQ340780 | – | sunken wood | ||
| SIb | – | – | DQ340782 | – | sunken wood | ||
| TLa | DQ340794 | – | DQ340785 | – | sunken wood | ||
| TRa | – | – | DQ340787 | – | sunken wood | ||
| VGc | – | DQ863944 | DQ340790 | DQ863951 | sunken wood | ||
| Adipicola longissima | SAL-2 | CHb | – | – | DQ340773 | – | sunken wood |
| RVd | DQ340799 | – | DQ340777 | – | sunken wood | ||
| SId | – | – | DQ340784 | – | sunken wood | ||
| SIc | – | – | DQ340783 | – | sunken wood | ||
| VGb | – | DQ863945 | DQ340789 | DQ863950 | sunken wood | ||
| VLb | – | – | DQ340791 | – | sunken wood | ||
| VLc | – | – | DQ340792 | – | sunken wood | ||
| VGa | DQ340798 | – | DQ340788 | – | sunken wood | ||
| Idas sp. | SAL-3 | CHa | DQ340801 | – | DQ340772 | – | sunken wood |
| CHc | – | DQ863946 | DQ340774 | DQ863949 | sunken wood | ||
| SIa | DQ340800 | – | DQ340781 | – | sunken wood | ||
| Idas sp. | SAL-4 | RVb | DQ340796 | – | DQ340776 | – | sunken wood |
| RVc | DQ340797 | – | DQ340793 | – | sunken wood | ||
| RVg | – | DQ863947 | DQ340779 | DQ863948 | sunken wood | ||
| TLb | – | – | DQ340786 | – | sunken wood |
2.2 Molecular methods
DNA was extracted from mussel tissues, avoiding the gills, which may contain many associated organisms, such as symbiotic bacteria. We used the ABI PRISM 6100 (Applied Biosystem) extraction and purification station. The Cytochrome Oxidase I (COI) mitochondrial gene was amplified for all the selected specimens using universal primers LCO 1490 and HCO 2198 developed by [17]. Then, one specimen for each identified terminal taxon was amplified for NADH dehydrogenase subunit 4 (ND4) mitochondrial gene using primers ArgBl and NAP2H [18,19], as well as a fragment of 18S rDNA nuclear gene using universal primers 18S1F, 18SBi, 18S5F, and 18S9R [12], and domains D1, D2 et D3 [20] of 28S rDNA nuclear gene using primers C1′(5′ACCCGCTGAATTTAAGCAT3′) and C4(5′TCGGAGGGAACCAGCTACTA3′).
PCR reactions were performed in a 25-μL final volume, containing approximately 3 ng template DNA, 1.5 mM MgCl2, 0.26 mM of each nucleotide, 0.3 μM of each primer, 5% DMSO and 0.75 unit of Taq Polymerase (Qbiogene). Amplification products were generated by an initial denaturing step of 4 min at 94 °C followed by 35 cycles (for COI, 28S and ND4)/37 cycles (for 18S) at 94 °C for 1 min, 50 °C for 1 min and 1 min at 72 °C, and by a final extension at 72 °C for 7 min. PCR products were purified using TM PCR Centrifugal Filter Devices (Millipore) and sequenced [21] on a Ceq2000TM automated sequencer (Beckman) for COI and at the Genoscope (Évry, France) for 18S, 28S and ND4, in both directions to confirm the accuracy of each sequence.
2.3 Molecular divergence among morphospecies
We used the same part of the COI gene as proposed by [11] for the Barcoding approach. Nucleotide-sequence divergences were calculated using the Kimura-two-parameter (K2P) model, which is suggested to be the best metric when distances are low [22]. In order to evaluate the species delineations, we compared genetic distances within morphospecies versus between morphospecies. We also used the neighbour-joining (NJ) analysis, implemented in MEGA 3 [23], to determine if haplotypes of each morphospecies were more closely related to each other than with haplotypes from other morphospecies.
2.4 Phylogenetic analyses
Phylogenetic relationships were estimated using three methods. First, we conducted an equally weighted maximum-parsimony (MP) research with a heuristic search option with 1000 random taxon-addition (RA) replicates and tree bisection and reconnection (TBR) branch-swapping using PAUP* v4.0b10 [24]. Second, the best fitting model of the sequence evolution for the maximum-likelihood (ML) analyses was determined by hierarchical likelihood ratio tests (hLTR) implemented in Modeltest version 3.06 [25]. The parameters estimated for the best-fit sequence evolution model were used in the ML heuristic searches with 100 RA replicates with TBR branch swapping using PHYML 2.4.4 [26]. For both MP and ML analyses, robustness of the nodes was assessed with nonparametric bootstrapping [27] with 1000 bootstrap replicates, TBR branch-swapping, and 10 RA replicates. Third, Bayesian analyses (BA) were performed with MrBayes v3.0 [28]. Six Markov chains were run in two parallel analyses using the parameters of the model used in the ML searches. Each Markov chain was run for 6 000 000 generations with a sampling frequency of one tree every hundred generations and a burning period of 15 000 trees. Convergence between the two analyses was assessed using likelihood curves, standard deviation of split frequencies, and potential scale-reduction factor (PSRF), as indicated by some authors [28,29]. All Bayesian analyses were performed on the cluster developed at the MNHN (17 nodes, 2-Go RAM per node, 30 AMDs 64 bits CPU's for the slave nodes and 4 Xeon 32 bits CPUs for the two master nodes).
The phylogenetic analyses were first performed on the RNAr 18S matrix constituted of a subset of eight specimens representative of the four identified morphospecies (Table 1) and of the sequences used in [4,9,12,30] (Tables 2 and 3).
List of Bathymodiolinae species used in this study and available from GenBank
| Species | GenBank N° | Habitat | |||
| 18S | 28S | COI | ND4 | ||
| Bathymodiolus heckerae BR | AY649830 | AY781139 | AY649793 | AY130245 | seep |
| Bathymodiolus heckerae WFE | AF221639 | AY781138 | AY649794 | AY130246 | seep |
| Bathymodiolus azoricus | AY649822 | AY781148 | AY649795 | AF128534 | vent |
| Bathymodiolus puteoserpentis | AF221640 | AY781151 | AY649796 | AF128533 | vent |
| Bathymodiolus brooksi AC | AY649826 | AY781136 | AY649797 | AY130247 | seep |
| Bathymodiolus brooksi WFE | AY649825 | AY781135 | AY649798 | AY649805 | seep |
| Bathymodiolus brevior MT | AY649824 | AY781150 | AY649799 | AY649806 | vent |
| Bathymodiolus brevior LBA | AY649827 | AY781143 | AY275544 | AY046277 | vent |
| Bathymodiolus marisindicus | AY649818 | AY781147 | AY275543 | AY046279 | vent |
| Bathymodiolus thermophilus A | AF221638 | AY781141 | AF456285 | AY649807 | vent |
| Bathymodiolus thermophilus B | AY649829 | AY781142 | AF456303 | AY649808 | vent |
| Bathymodiolus aff. thermophilus | AY649823 | AY781140 | AF456317 | AY649809 | vent |
| Bathymodiolus childressi | AF221641 | AY781137 | AY649800 | AY130248 | seep |
| Bathymodiolus mauritanicus | AY649828 | AY781144 | AY649801 | AY649810 | seep |
| Gigantidas gladius | AY649821 | AY781149 | AY649802 | AY649813 | vent |
| Bathymodiolus tangaroa | AY649820 | AY781134 | AY608439 | AY649811 | seep |
| Tamu fisheri | AF221642 | AY781132 | AY649803 | AY649814 | seep |
| Idas washingtonia | AF221645 | AY781146 | AY275546 | AY649815 | whale bones, wood |
| Idas macdonaldi | AF221647 | AY781145 | AY649804 | AY649816 | seep |
| NZ3 | AY649819 | AY781133 | AY608440 | AY649812 | vent |
| Myrina pacifica | AF221646 | – | – | – | whale bones |
| Idas arcuatilis | AF221643 | – | – | – | whale bones |
| Adipicola arcuatalis | AF221644 | – | – | – | whale bones |
List of Mytilidae (except Bathymodiolinae) and Bivalvia outgroups used in this study
| Subclass | Family | Sub-family | Species | GenBank N° | |||
| 18S | 28S | COI | ND4 | ||||
| Pteriomorpha | Mytilidae | Crenellinae | Musculista senhousia | AF124207 | – | AB076942 | – |
| Crenellinae | Musculus discors | AF124206 | – | – | – | ||
| Lithophaginae | Lithophaga nigra | AF124209 | – | – | – | ||
| Lithophaginae | Lithophaga lithophaga | AF124208 | – | AF120644 | – | ||
| Modiolinae | Benthomodiolus lignicola | AF221648 | AY781131 | AY275545 | AY649817 | ||
| Modiolinae | Geukensia demissa | L33450 | – | AY621926 | – | ||
| Modiolinae | Modiolus auriculatus | AF117735 | – | – | – | ||
| Modiolinae | Modiolus modiolus | EF526454 | EF526455 | U56848 | EF526453 | ||
| Modiolinae | Myrina pacifica | AF221646 | – | – | – | ||
| Mytilinae | Perna viridis | – | – | AF298852 | |||
| Mytilinae | Brachidontes modiolus | – | – | AY621918 | – | ||
| Mytilinae | Brachidontes exustus | AF229623 | – | – | – | ||
| Mytilinae | Hormomya exustus | – | – | AY621945 | – | ||
| Mytilinae | Hormomya domingensis | AF117736 | – | – | – | ||
| Mytilinae | Ischadium recurvum | – | – | AY621929 | – | ||
| Mytilinae | Mytilus galloprovincialis | L33451 | – | – | – | ||
| Mytilinae | Mytilus trossulus | L33453 | – | – | – | ||
| Mytilinae | Mytilus californianus | L33449 | – | – | – | ||
| Mytilinae | Mytilus edulis | L24489 | – | AY377727 | – | ||
| Mytilinae | Trichomya hirsuta | – | – | AY296816 | – | ||
| Dacrydiinae | Dacrydium zebra | – | – | AB076945 | – | ||
| Ostreidae | Crassostrea virginica | X60315 | – | – | – | ||
| Ostreidae | Ostrea edulis | U88709 | – | AF120651 | – | ||
| Pinnidae | Atrina pectinata | X90961 | – | AB076914 | – | ||
| Arcidae | Arca noae | X90960 | – | – | – | ||
| Arcidae | Glycymeris sp | X91978 | – | – | – | ||
| Arcidae | Barbatia virescens | X91974 | – | – | – | ||
| Pectinidae | Chlamys islandica | L11232 | – | – | – | ||
| Pectinidae | Placopecten magellanicus | X53899 | – | – | – | ||
| Paleoheterodonta | Unionidae | Elliptio complanata | AF117738 | – | – | – | |
| Heterodonta | Myidae | Mya arenaria | AF117739 | – | – | – | |
| Protobranchia | Solemyidae | Solemya reidi | AF117737 | ||||
| Solemyidae | Solemya velum | AF120524 | – | U56852 | – |
Then, as there was no consensus on the relationships between Mytilids subfamilies (see [13]), we analyzed the relationships of the subfamily Bathymodiolinae with other subfamilies of Mytilidae (Rafinesque, 1815). For that purpose, we used COI data from GenBank to determine which taxa were the more closely related to the bathymodiolin group in order to use them as outgroups in the study of the relationships within this group. We included in this analysis one sequence from each one of our morphospecies and one to five sequences for each Mytilid subfamily. In this analysis, we only used the first and second codon positions of the COI gene, because the third position of this gene was saturated.
The outgroups identified by this analysis were subsequently used in the analysis of the COI matrix, including the GenBank sequences from [9,31], attributed to Bathymodiolinae (Table 2), and our own sequences of sunken wood mytilids from the Solomon Islands.
Finally, we sequenced two other genes (ND4 and rDNA 28S genes) used by [5,9] to improve our phylogenetic analyses. For that purpose, one sequence of each one of these two genes was used for each morphospecies validated by the analysis of COI diversity (Table 1). We first explored separately each one of the four single gene datasets by the maximum-likelihood approach and performed an incongruence length difference (ILD) test [32] in order to validate congruence between all genes. Mitochondrial dataset analyses were first performed using the whole dataset (i.e. the three positions of each codon). As Jones et al. [9] suggested that these genes are saturated at the third codon position among Bathymodiolinae, each mitochondrial dataset was also analyzed without this position. Then, taking into account the results of the effect of the saturation of the third position, we performed a combined Bayesian analysis for which the number of substitution types of each gene-specific model, as defined using Modeltest 3.06, was implemented.
3 Results
3.1 The diversity of sunken wood mussels from Solomon Islands
Analysis of the COI gene yielded four distinct haplotype clusters in the NJ tree (i.e. phylotypes, figure not shown). These phylotypes were separated by large genetic distances (ranging from 15.1% to 19.2%), whereas the genetic distances within phylotypes were not higher than 1.5% (Table 4). There was no overlap between the ranges of intra-phylotypic and inter-phylotypic genetic distances. Moreover, the NJ tree indicated that haplotypes obtained for each morphospecies were in the same phylotype and thus that within a morphospecies, haplotypes were more closely related to each other than to haplotypes obtained for other morphospecies. These four phylotypes were consistent with the morphospecies delimitation that was a priori defined looking at the global morphology of the shells (named SAL-1 to SAL-4).
Matrix of genetic distance (K2P) within and between phylotypes. Standard errors are in brackets
| SAL-1 | SAL-2 | SAL-3 | SAL-4 | |
| SAL-1 | 0.013 (0.003) | |||
| SAL-2 | 0.157 (0.018) | 0.003 (0.002) | ||
| SAL-3 | 0.180 (0.018) | 0.157 (0.004) | 0.001 (0.001) | |
| SAL-4 | 0.186 (0.018) | 0.151 (0.016) | 0.192 (0.018) | 0.015 (0.004) |
3.2 Phylogenetic analyses
Whatever the phylogenetic reconstruction method used, all specimens sequenced for the rRNA 18S gene belong to a monophyletic group that includes all other mussels associated with reducing environments (Fig. 1). However, as shown by [4,12], the low variability of this gene at this level did not allow one to further elucidate the relationships.

Phylogenetic relationships among mytilids (18S gene) determined using the Bayesian approach. Maximum likelihood was calculated using TrNef+[Γ]+I with I=0.5978, [Γ]=0.7230, and equal frequencies of nucleotides (−lnL=5324.5322; K=4). Bootstrap proportions and Bayesian posterior probabilities were presented at nodes. Nodes for which posterior probabilities were below 0.95 and/or bootstrap value below 80% were collapsed.
The variability of the first and second positions of the COI gene allowed us to resolve the phylogenetic relationships within mytilids. The same relationships were obtained with the three reconstructions methods (Fig. 2). These analyses suggested that mytilids are divided into two major and well-supported lineages. (i) One of these lineages included Lithophaga lithophaga and Dacrydium zebra at the base of two resolved clades. One of these clades included the species Trichomya hirsuta, Musculista senhousia, Mytilus edulis, and Perna viridis, and the other clade regrouped the species Ischadium recurvum, Geukensia demissa, Brachidontes exustus and Hormomya domingensis. These two clades both include species attributed to Mytilinae and thus make this sub-family polyphyletic. (ii) The second lineage regrouped all the Bathymodiolinae, including our Solomon Islands' sunken woods morphospecies. This result confirmed that our sunken wood mussels belong to Bathymodiolinae. This lineage displayed Modiolus modiolus (Linnaeus, 1758) in the most basal position immediately followed by Benthomodiolus lignicola (Dell, 1987), indicating that these two shallow water mussels are the closest relatives to Bathymodiolinae. These two species were subsequently used as outgroups in our analysis of the relationships within Bathymodiolinae.
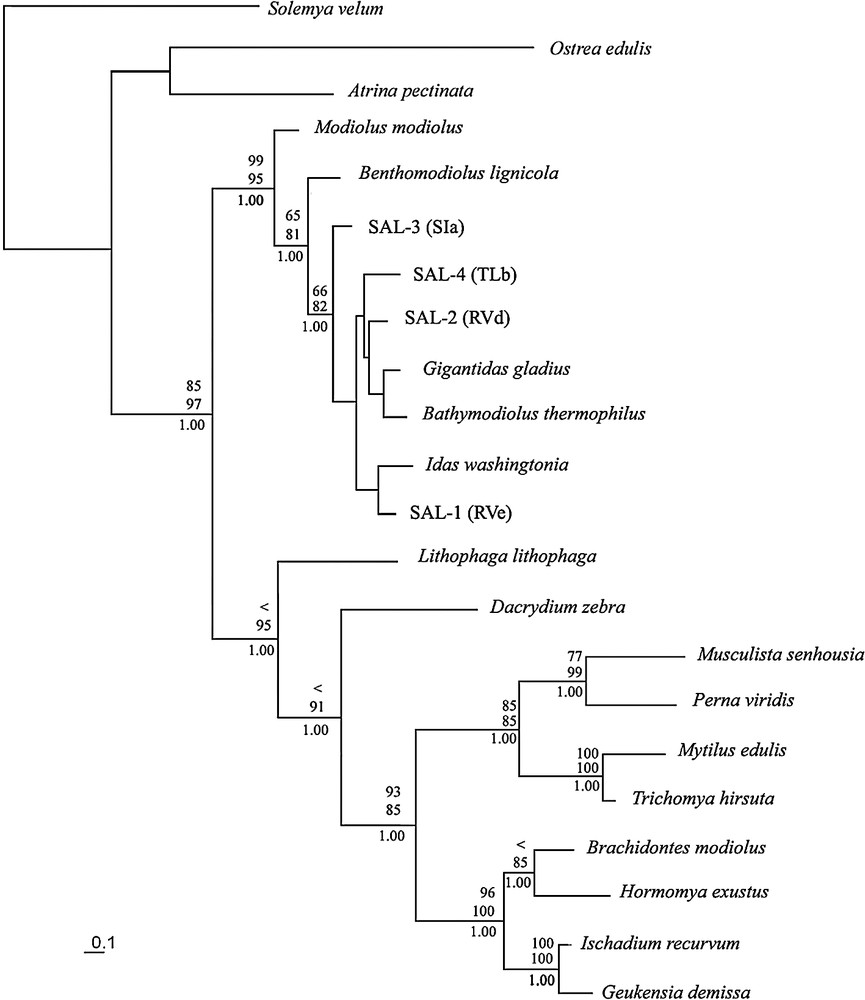
Phylogenetic relationships among mytilids (COI gene) determined using the maximum-likelihood method. Likelihood substitution model: HKY-85+[Γ]+I with [α]=0.449 and base frequencies and ti:tv (4.675) estimated from the data. Bootstrap proportions for parsimony (upper) and ML (middle) analysis are presented (percentage of 1000 replicates). Dashes are values <50%. Bayesian posterior clade probabilities (bottom) are presented (consensus of 50 000 trees).
The latter analysis was conducted on COI data of all our specimens together with available Bathymodiolinae sequences from GenBank (Table 2), using M. modiolus and Be. lignicola as outgroups. The Bayesian analysis of this matrix, which included data on the mytilids from hydrothermal vents, cold seeps, whale falls, and sunken woods, revealed that all vent and seep mussels involved in our analysis are derived taxa within Bathymodiolinae (Fig. 3), but this result was poorly sustained on maximum-likelihood and maximum-parsimony trees by bootstraps values. Additionally, this analysis suggested that SAL-1 is a sister species for Idas washingtonia.
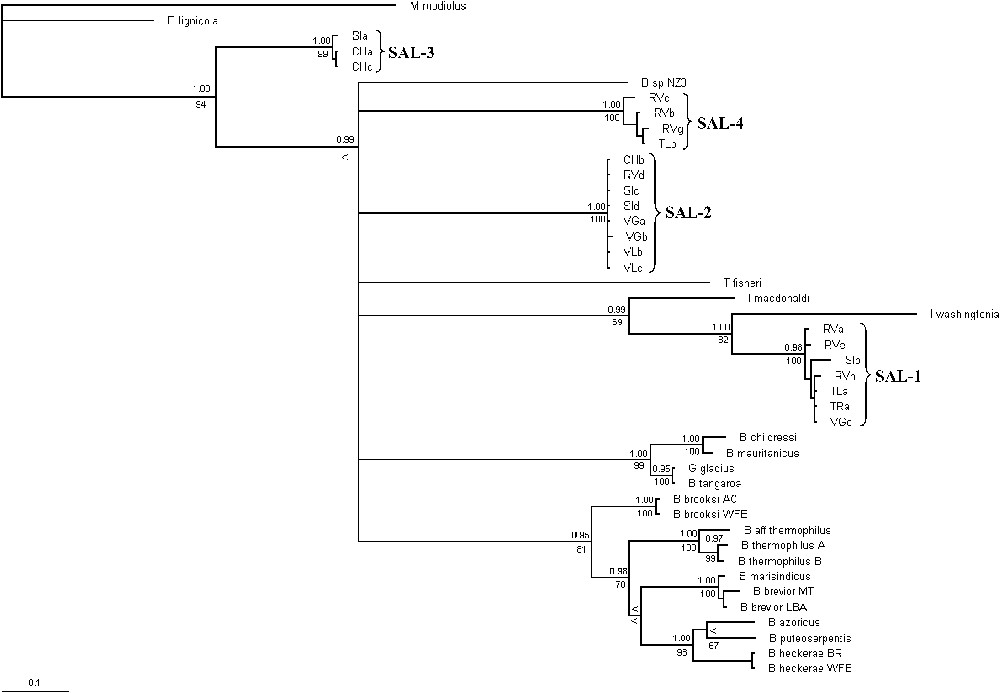
Phylogenetic relationships among Bathymodiolinae (COI gene) using the maximum-likelihood method. Likelihood substitution model: HKY-85+[Γ]+I, with [α]=0.704 (−lnL=3973.9949) and base frequencies and ti:tv (13.34) estimated from the data. Bayesian posterior probabilities (top) and ML (bottom) analysis are presented (percentage of 1000 replicates). Dashes are values of <50%. Nodes for which posterior probabilities were below 0.95 and bootstrap value below 80% were collapsed.
When removing the third position of the codon in the COI dataset, all the resolution within the in-group was lost. Conversely, when the third position of the codon was removed in the ND4 dataset, the resolution of the tree was improved. Thus, as already suggested by [9], the COI dataset within Bathymodiolinae appeared only slightly saturated, contrary to the ND4 dataset. Thus in the combined four gene analysis, we removed the third codon position only for ND4. The combined four-gene analysis exhibited congruent topologies with the single-genes analyses. This combined Bayesian analysis first confirmed the basal position of individuals identified as SAL-3 relatively to all other species, NZ3 excluded (Fig. 4). Several lineages were identified in this most derived node but the relationships among them were not resolved by our analysis. Indeed, this analysis did not clarify relationships between T. fisheri, SAL-2, SAL-4 and three well-supported clades: (i) a ‘childressi’ clade that included B. childressi, B. mauritanicus, G. gladius and B. tangaroa; (ii) an ‘Idas’ clade that included SAL-1, I. washingtonia and I. macdonaldi; (iii) a ‘Bathymodiolus’ clade with well-supported internal nodes.
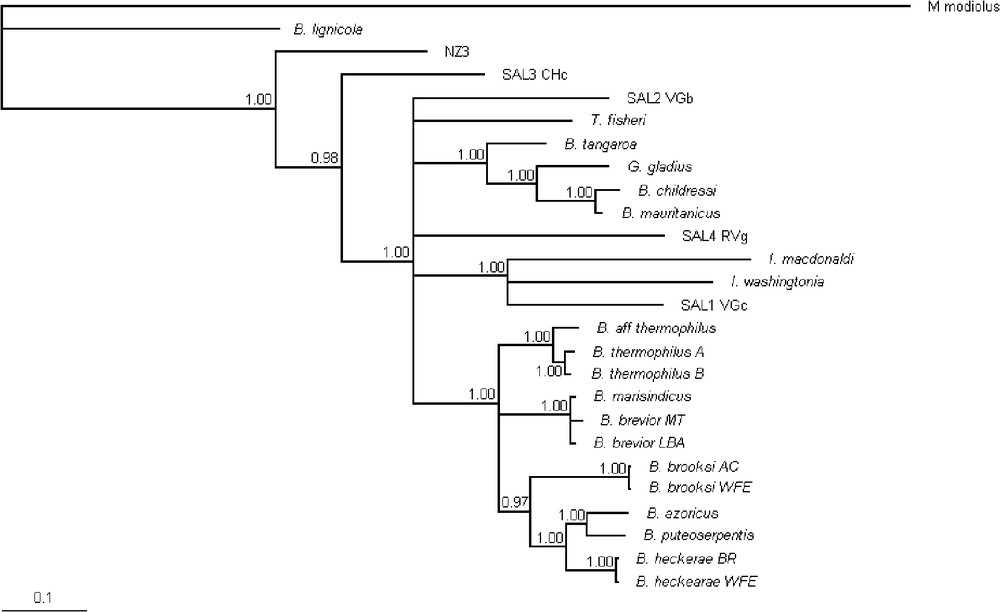
Phylogenetic relationships among Bathymodiolinae based on four genes (COI, ND4, 28S, 18S) using the Bayesian analysis. For additional genes, Modeltest3.4 analyses allowed us to use HKY+[Γ] for ND4 (−lnL=2636.3689; K=5), TrN+[Γ]+I for 28S rRNA (−lnL=2370.9358; K=7), K80+[Γ]+I for 18S rRNA (−lnL=2820.5027; K=3). Bayesian posterior probabilities are presented. Nodes for which posterior probability was less than 0.95 were collapsed.
4 Discussion
Using a Barcoding-like approach, we were able to confirm our morphospecies delimitation. Indeed, four genetic clusters were detected, each one corresponding to a unique morphospecies. Moreover, there was no overlap between intra-phylotype and inter-phylotype genetic distances. The calculated intra-phylotypic distances within the four phylotypes were similar to intraspecific distances calculated within other bathymodilin species [5,6,31]. We then tried to attribute our morphospecies to described species. Rudo von Cosel identified the phylotype SAL-2 as Adipicola longissima (Thiele and Jaeckel, 1932). However, he could only give a genus name (Idas) to the three other morphospecies. We thus overall suggest that four distinct species from two genera were present in our sampling from the Solomon Islands. The three Idas species (SAL-1, -3 and -4) might correspond to new species.
Both analyses of nuclear 18S rRNA and mitochondrial COI genes revealed that the four sunken woods mussel species sampled from the Solomon Islands were included in the monophyletic group recognized by [4] as involving all species from hydrothermal vents, cold seeps and whale falls. Thus, our results confirmed, as suggested by [4], that the sunken woods mussels are closely related to mussels from reducing environments.
Moreover, contrary to the analysis of the 18S gene performed by [4], the analysis of the COI gene permitted to determine what outgroups to use in the analyses of the subfamily Bathymodiolinae. Indeed, this clade was robustly rooted on the species Modiolus modiolus. To determine how to root our ingroup, we needed to examine the relationships among Mytilids subfamilies. As a result, we confirmed the polyphyletic nature of Modiolinae and Mytilinae, already revealed by [12] with the analysis of 18S variability. Our results also permitted to clarify the points raised by [13] concerning the classification of Mytilidae. For example, from the data of [4,12,13], one can consider that the subfamily Arcuatulinae, defined by [15], was supported by the clustering of Hormomya domingensis and Geukensia demissa. Our analysis of COI data also supported this proposition. Moreover, [13] using several morphological analyses [14,15] and fossil records [16], as well as results of [4,12] suggested to cluster in the family Lithophagidae (Adams, 1857) five sub-families, among which Lithophaginae (Adams, 1857), Dacrydiinae (Ockelmann, 1983), Modiolinae s.s (Keen, 1958), and Bathymodiolinae. Our analysis revealed that the subfamily Bathymodiolinae is robustly rooted within a monophyletic group that included the species Modiolus modiolus that could be considered as a Modiolinae s.s. However, contrary to the proposition of [13], Lithophaginae and Dacrydiinae are not rooted within this clade.
Recent studies on Bathymodiolinae largely covered the diversity of vent and seep lineages, letting the sunken wood species apart. The study of [10], covering species from both Atlantic and Pacific ridge and from seeps and vent, revealed three distinct lineages. The study of [9] revealed that the taxa stemming from basal nodes occur in shallow sites, whereas the more derived taxa tend to occur at deeper sites. Thus, although these authors noted some exceptions to this trend, their dataset roughly support the general pattern recognized by Craddock et al. [3].
Our study recovered the hydrothermal lineages revealed by [9]. The slight differences observed with the topology of [9] concerned the position of B. brooksi within the ‘thermophilus’ clade and the unresolved position of B. tangaroa and G. gladius included within the ‘childressi’ clade in the analysis of Jones et al. [9]. These differences may either be due to the addition of more species in the dataset or to the slight differences between the models used for DNA evolution. Most of these lineages appear as strictly linked to hydrothermal environment. However, compared to previous studies, even if we increased the number of sunken wood species included in the phylogenetic analyses, we are far to cover the specific diversity of the sunken wood mytilids. Therefore, we cannot exclude that some sunken wood species belong to these apparently strictly hydrothermal lineages.
Our study, which added to the available datasets four sunken wood species, revealed that one of them – the SAL-3 morphospecies – has a basal position within the Bathymodiolinae monophyletic group. Thus, the results of [4], based only on the position of Benthomodiolus lignicola – which were moreover obtained with a poorly informative gene – are here supported by an enlarged dataset that includes more sunken wood species and more genes. In our analyses, vent and seep Bathymodiolinae appear as derived species, as well as the sunken wood morphospecies SAL-1, SAL-2 (A. longissima), and SAL-4.
Last, the four-gene analysis suggested that the lineage conducing to the undescribed NZ3 mussel sampled from a shallow hydrothermal seamount emerged after the lineage of Be. lignicola, but before that of the SAL-3 morphospecies, which emerged before the lineage that included all other Bathymodiolinae. Thus, the relative positions of NZ3 mussel and Be. lignicola suggest the existence of another older colonization event from wood to vents and seeps. This hypothesis may also explain why, based on morphological characters, it is difficult to place the NZ3 mussel within the genus Bathymodiolus and, more generally, within the genera sampled in modern hydrothermal vents and seeps. Modioliform mussels are known from hydrothermal vents since at least the Mesozoic [2]. However, it has recently been proposed that the first modioliform mussels presenting the morphological characteristics of modern Bathymodiolinae appeared during the Eocene both on cold seep carbonate and on sunken wood assemblages [33,34]. Thus, Be. lignicola and NZ3 mussels could belong to older lineages.
Overall, our results stress that, to understand the origin of hydrothermal vents and seep species, we need to have a better taxonomic coverage within the Bathymodiolinae monophyletic group. For that purpose, sunken woods and whalebones species from a larger geographical range must be included into the phylogenetic datasets.
Acknowledgements
We are grateful to the crew of the R/V Alis and the technical support of the IRD at Nouméa, to the staff of the ‘Service de systématique moléculaire’ at the ‘Muséum national d'histoire naturelle, Paris’, for technical facilities, and to the ‘Consortium national de recherche en génomique’, Genoscope. We thank P. Bouchet and B. Richer de Forges, the co-principal investigators of the Salomon2 cruise, and T. Haga for his great help in sampling during the cruise. We are grateful to Cyrille D'Haese for the development and access to the cluster at the MNHN. Special thanks to F. Pleijel for constructive comments and improvements of the English manuscript, and to F. Rousseau and an anonymous referee for helpful advices. This work belongs to a project included in the European research group DiWood, supported by the CNRS (France).