

Version française abrégée
Le genre Microbuthus a été décrit par Kraepelin [1], fondé sur l'espèce Microbuthus pusillus Kraepelin, 1898 collectée dans la région de Tadjura Bay (Tadscharabay), dans le golfe d'Aden (Yémen). Par la suite, Birula [2] transfère Butheolus litoralis Pavesi 1885 décrite d'Assab, côte de la mer Rouge, en Érythrée, dans le genre Microbuthus. Le genre demeure composé de ces deux seules espèces jusqu'à la description par Vachon [3] de Microbuthus fagei, collecté le long du rivage de l'océan Atlantique, à Nouakchott, dans le Sud de la Mauritanie. Plus récemment, Lourenço [4] suggère l'existence d'une sous-espèce de M. fagei dans le Sud du Maroc, et la décrit en tant que Microbuthus fagei maroccanus Lourenço, 2002.
Parmi les nombreux genres de la famille des Buthidae, Microbuthus demeure un groupe énigmatique. Les raisons pour cela tiennent non seulement aux caractéristiques morphologiques très particulières du genre, mais aussi à la rareté du matériel connu. De plus, certains des spécimens types semblent égarés, ou même perdus, et ceci complique davantage une définition précise de la composition du groupe. À titre d'exemple, le spécimen type de Butheolus litoralis décrit par Pavesi n'a été retrouvé dans aucun musée d'Italie, et même le matériel type de M. fagei, en principe déposé au Muséum à Paris, est désormais égaré.
La collecte récente d'un exemplaire unique d'un Microbuthus en Égypte amène à la description d'une nouvelle espèce, Microbuthus flavorufus sp. n. Cette découverte est par ailleurs la première mention du genre en Égypte. Avec la description de M. flavorufus sp. n., le nombre total des espèces connues s'élève à cinq. Le modèle de distribution périsaharienne et disjointe présenté par les espèces du genre Microbuthus est à nouveau confirmé.
Biogéographie du genre Microbuthus
Au cours d'un colloque organisé par la Société de biogéographie sur les origines du peuplement de l'Afrique septentrionale, Furon [5] soulignait déjà que les faunes et les flores autochtones du Sahara pouvaient être très anciennes. Leur répartition actuelle ne serait pas uniquement la conséquence de facteurs paléogéographiques proprement dits, mais également de facteurs paléoclimatiques. Ces phénomènes paléoclimatiques prennent toute leur importance au Quaternaire. Pendant les périodes glaciaires d'Europe (et d'Amérique du Nord), l'Afrique connaissait des périodes pluviales relativement intenses, avec une grande extension des glaciers de montagne (en Afrique orientale en particulier). Le Sahara a connu des périodes humides, dont la dernière est géologiquement très récente, se terminant vers 3000 ans BP.
Ainsi que cela a déjà été souligné par Qi et Lourenço [6], la composition actuelle de la faune saharienne est en réalité l'héritage de faunes plus anciennes, déjà présentes en Afrique du Nord depuis le début ou tout au moins la moitié de la période Cénozoïque [7]. L'Afrique du Nord a connu diverses vicissitudes paléoclimatologiques au cours des derniers millions d'années, certaines même au cours de la période quaternaire. Le Sahara a pu traverser une série de périodes humides, la plus récente ayant lieu entre 10 000 et 5000 ans BP, et ce n'est que depuis 3000 ans environ que le Sahara connaît son aridité actuelle [8–10]. Des études récentes suggèrent que le désert du Sahara pourrait être bien plus ancien que ne l'indiquent les datations préalablement proposées [11] ; cependant, il semble plausible de postuler que des régions arides auraient existé comme des zones enclavées, au moment même où le climat général de l'Afrique du Nord a pu connaître des conditions plus tempérés. Dans de telles régions désertiques, une faune scorpionique spécialisée a pu évoluer. À l'opposé, avec l'expansion du désert, d'autres lignées moins bien adaptées aux conditions d'aridité, et habitant des environnements plutôt tempérés, ont connu une régression importante dans leurs distributions. Par ailleurs, certaines de ces lignées ont pu connaître une sélection négative et disparaître totalement. Dans d'autres cas, leurs populations ont vu leurs distributions réduites à des enclaves écologiques, avec comme conséquence des distributions fortement disjointes.
Parmi les modèles de distribution observés pour les scorpions de l'Afrique du Nord [6], la répartition totalement disjointe présentée par le genre Microbuthus est particulièrement remarquable. Deux espèces sont présentes en Mauritanie et au Maroc, à l'ouest, et deux autres en Érythrée et à Djibouti, à l'est [4]. Vachon [7,12] signalait déjà une répartition particulièrement localisée des espèces de Microbuthus. Il définit ce modèle comme celui des genres habitant un territoire restreint, mais très dissocié. Une question déjà posée par Vachon [12] peut une nouvelle fois être soulevée : comment expliquer la présence de formes apparentées tant au nord qu'au sud, mais aussi à l'est et à ouest du Sahara ? D'après Braestrup [13], un mécanisme d'échanges à travers le Sahara a souvent été utilisé comme argument pour expliquer la répartition des faunes sahariennes. Il mentionne des traversées du domaine déserticole, soit par des éléments méridionaux (éthiopiens) qui auraient gagné les régions septentrionales, soit par des éléments septentrionaux (paléarctiques), dispersés jusque dans des régions du Sud du Sahara. Une telle hypothèse est plausible pour des organismes dotés d'une grande capacité de dispersion, capables de parcourir de longues distances et d'établir de nouvelles colonies loin de leur territoire. Or, la faune des scorpions est constituée, en général, de populations autochtones stables et prévisibles, c'est-à-dire globalement très anciennes et individuellement très localisées, peu enclines à de telles migrations. De ce fait, les répartitions géographiques actuelles des différents groupes de scorpions doivent être davantage associées à une situation plutôt relictuelle qu'à la conséquence d'une expansion dynamique.
Cette dernière hypothèse semble s'appliquer aux différentes espèces du genre Microbuthus, qui présentent actuellement des répartitions géographiques très disjointes. Les différentes espèces, morphologiquement très voisines, habitent également des milieux semblables. Selon Vachon [16], les espèces du genre Microbuthus peuvent être classées comme des scorpions « halophiles », puisqu'elles sont retrouvées exclusivement dans les zones côtières d'Afrique, aussi bien orientale qu'occidentale. La nouvelle espèce décrite à présent vient conforter le modèle de distribution défini pour le genre Microbuthus.
Taxonomie
Microbuthus flavorufus sp. n.
Diagnose. Scorpion de petite taille avec 19,7 mm de longueur totale. Coloration de jaunâtre à jaune rougeâtre. Peignes avec 8–8 dents. Anneau caudal I plus large que long ; anneaux II à IV aussi longs que larges ; anneau V plus long que large. Éperon tibial absent de la patte III et très réduit sur la patte IV. Modèle trichobothrial : néobothriotaxie minorante ; pince avec l'absence des trichobothries est, Esb, Eb3 ; tibia avec la trichobothrie d2 très réduite et sept trichobothries externes ; fémur avec l'absence de la trichobothrie d2 et une trichobothrie d5 très réduite.
1 Introduction
The genus Microbuthus was described by Kraepelin [1], with Microbuthus pusillus Kraepelin, 1898 from the region of Tadjura Bay (Tadscharabay) in the Gulf of Aden (Yemen) as type species. A second species, Butheolus litoralis Pavesi, 1885 from Assab, on the Red Sea coast of Eritrea, was subsequently transferred to the genus by Birula [2]. The genus Microbuthus was represented solely by these two species until the description of Microbuthus fagei, collected at Nouakchott in the coastal region of the Atlantic Ocean, in the South of Mauritania [3]. More recently, Lourenço [4] has shown that a subspecies of Microbuthus fagei exists in the South of Morocco. He described this as Microbuthus fagei maroccanus Lourenço, 2002.
Among the numerous known genera of Buthidae, Microbuthus remains enigmatic. The reasons for this are associated not only with several unique morphological features, but are also due to the scarcity of known material. Furthermore, some of the type specimens have been mislaid, which creates further difficulties in presenting a well-defined and precise composition of the group. For example, the type specimen of Butheolus litoralis cannot be found in any of the Italian Museums, and even the type material of M. fagei in the collections of the ‘Muséum’ in Paris has been mislaid.
The recent collection of a single specimen of Microbuthus in Egypt has led to the description of a new species, Microbuthus flavorufus sp. n. This is moreover the first record of the genus Microbuthus from Egypt. With the description of M. flavorufus sp. n., the total number of known species in the genus is raised to five. The disrupted Perisaharan pattern of distribution previously observed among the species of the genus is once again confirmed.
2 Biogeography of the genus Microbuthus
In a symposium of the Society of Biogeography, dealing with the origins and settlement of the fauna of North Africa, Furon [5] suggested that the flora and fauna now present in the Sahara may well be very old. Their present pattern of distribution reflects not only the consequences of palaeogeographic factors, but is also largely the result of various palaeoclimates. These palaeoclimatic events had an important impact during the Quaternary when Europe (and North America) faced periods of glaciation. During these, Africa experienced periods of intense rain and, additionally, an increase in the amount of ice on the mountains (particularly in Oriental Africa). The last wet period in the Sahara was very recent, only some 3000 years BP.
These facts were discussed once again by Qi and Lourenço [6], who agreed that the present composition of the Saharan fauna is, in fact, the heritage of ancient faunas that have been present in North Africa since the beginning of the Cenozoic, or at least since Middle Cenozoic times [7]. North Africa has experienced numerous other palaeoclimatological vicissitudes in the last few million years, some of which were in more or less recent Quaternary times. The Sahara has undergone a long series of wet periods, the most recent occurring 10 000–5000 years BP. It was not until about 3000 years BP that the Sahara assumed its present arid state [8–10]. Recent studies suggest that the Sahara desert may be much older than was previously thought [11]. It seems reasonable, therefore, to postulate that extremely arid areas may have existed as patchy desert enclaves for a very long time, even when the general climate of North Africa enjoyed more mesic conditions. In these desert regions, a specialized scorpion fauna would have evolved. In contrast, other lineages less well adapted to drought, and previously present only in mesic environments, have regressed markedly in their distribution. They have therefore experienced negative selection and are on the road to extinction. In other cases, populations have been reduced to very limited and patchy zones sometimes with remarkable disjunctions in their distribution patterns.
Among the patterns observed today in the distribution of North African scorpions [6], one of the most remarkable is the disjunction presented by the distribution of the genus Microbuthus, with two species known in Mauritania and Morocco in the West, and two others respectively in Eritrea and Djibouti in the East. Vachon [7,12] had already drawn attention to this extremely localized pattern of distribution of the species of Microbuthus, and defined it as a ‘disrupted and limited territory’. Vachon also attempted to explain the observed pattern, and made reference to Braestrup [13], who had suggested a mechanism for exchanges through the Sahara desert. According to this, southern elements (Ethiopian) were able to reach the northern regions, and northern elements (Palaearctic) were able to disperse to the southern regions of the Sahara. This hypothesis is valid for dynamic elements with a marked capacity for dispersion. On the other hand, scorpion populations are, in most cases, very predictable and stable. They are quite incapable of readapting to new environments. The present pattern of distribution of several groups of scorpions, and in particular the one presented by the genus Microbuthus, must reflect a large range of distribution in the past. The distinct palaeoclimatic vicissitudes experienced by the Sahara have constituted an important selective factor over its scorpion populations. The reaction of these to abiotic factors was certainly varied depending on their own ecological strategies [14,15]. In some cases, the populations showed a significant regression in their distribution, and some populations may well have totally vanished. These regressions led to marked disruptions in geographic distributions and resulted in their present patchy distribution. This hypothesis explains the disrupted pattern of distribution presented by the species of Microbuthus. All species are very similar morphologically and inhabit similar ecological environments. In fact, according to Vachon [16], Microtityus species can be classified as halophiles, since they are exclusively found in coastal zones from the Red Sea to the Atlantic Ocean. The new species described below provides additional support for this pattern of distribution (Fig. 14).
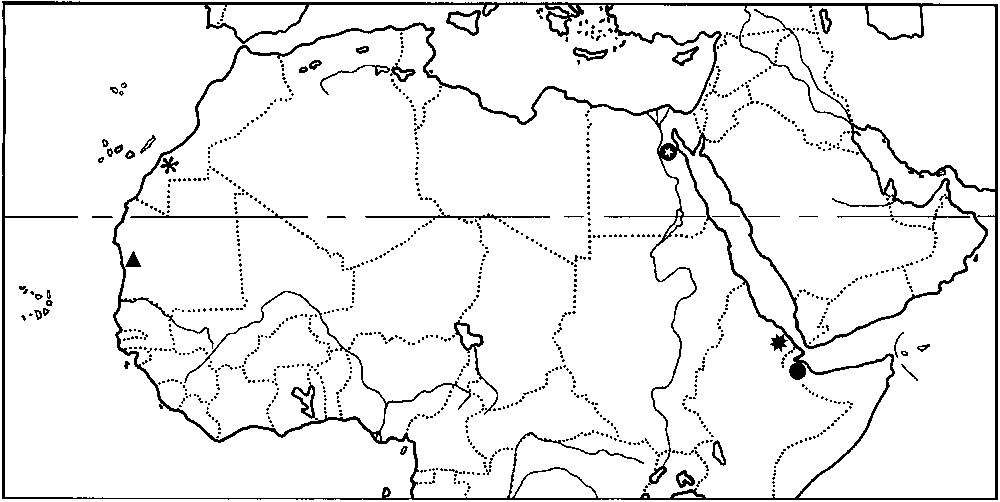
Map of North Africa, showing the type localities of the known species of Microbuthus. M. litoralis (black star), M. pusillus (black circle), M. fagei (black triangle), M. maroccanus (black flower), M. flavorufus sp. n. (black circle with white star).
3 Taxonomic treatment
Family Buthidae, C.L. Koch, 1837
Genus Microbuthus Kraepelin, 1898
Microbuthus flavorufus sp. n. (Figs. 1–13)
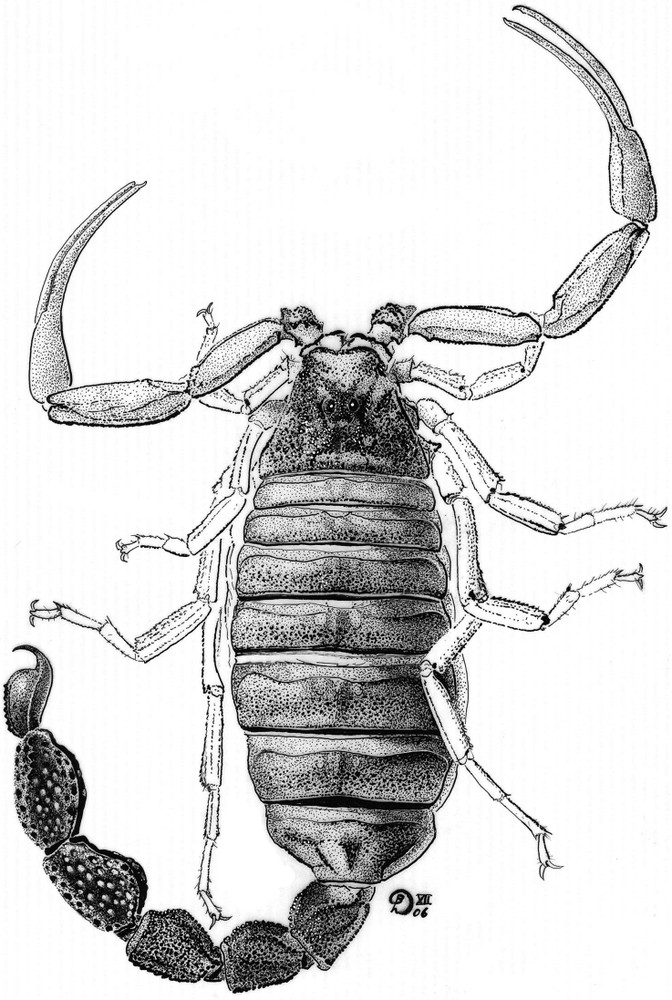
Microbuthus flavorufus sp. n. Female holotype. Habitus.
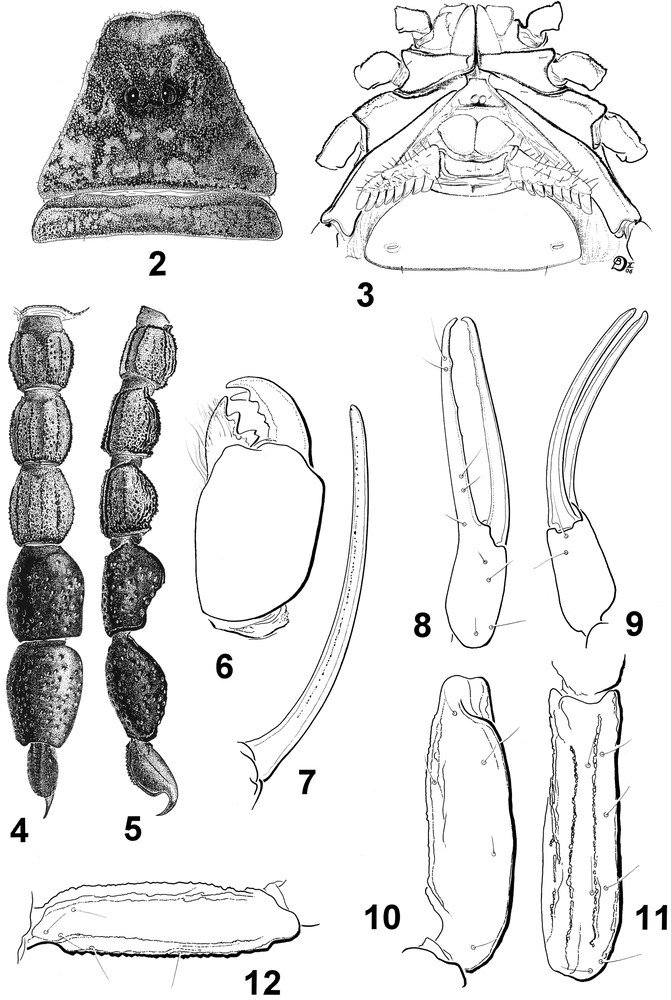
Microbuthus flavorufus sp. n. Female holotype. 2. Carapace and tergite I. 3. Ventral aspect showing coxapophysis, sternum, genital operculum and pectines. 4–5. Metasoma and telson, ventral and lateral aspects. 6. Chelicera. 7. Cutting edge of movable finger with rows of granules. 8–12. Trichobothrial pattern. 8–9. Chela dorso-external and ventral aspects. 10–11. Patella, dorsal and external aspects. 12. Femur, dorsal aspect.

Microbuthus flavorufus sp. n. Female holotype alive, in its natural habitat.
Type material. Female holotype, Egypt, 22 km south of Zafarana, Koraimat road (29.077°N, 32.451°E), 5/V/2006 (P. Geniez). Deposited within the collections of the ‘Muséum national d'histoire naturelle’, Paris.
Etymology. Specific name refers to the reddish-yellow coloration of the new species.
Diagnosis. A small scorpion only 19.7 mm in total length. Coloration not dark, as in the other species, but ranging from yellowish to reddish-yellow. Pectinal tooth count 8–8. Metasomal segment I wider than long; segments II to IV with similar values for length and width; segment V longer than wide. Tibial spurs on leg III absent; extremely reduced, almost obsolete on leg IV. Trichobothrial pattern: minorante neobothriotaxy; chela (hand + fixed finger) with the absence of trichobothria est, Esb, Eb3; patella with trichobothria d2 extremely reduced and seven external trichobothria; femur with absence of trichobothria d2, and an extremely reduced d5.
By its general morphology, the new species shows affinities in particular to Microbuthus litoralis from Djibouti. The known species of Microbuthus can be readily distinguished with the aid of the key presented here. In this, the subspecies Microbuthus fagei maroccanus is raised to the rank of species as Microbuthus maroccanus n. rank.
- 1. Patella with six external trichobothria; metasomal segments IV and V not enlarged...............2
- (1). Patella with seven external trichobothria; metasomal segments IV and V generally enlarged......3
- 2. Sternites yellowish; pedipalps with brownish spots ...............................M. maroccanus
- (2). Sternites blackish-brown; pedipalps yellowish without spots..........................M. fagei
- 3. General coloration blackish-brown; femur trichobothrium d5 absent.......................4
- (3). General coloration yellowish to reddish-yellow; femur trichobothrium d5, very reduced but present ............................M. flavorufus sp. n.
- 4. Metasomal segments and vesicle with intense setation .................................M. pusillus
- (4). Metasomal segments and vesicle without intense setation............................M. litoralis
Description based on female holotype (measurements following the description).
Coloration. Basically yellowish to reddish-yellow. Prosoma: carapace yellowish, spotted with reddish pigment, particularly in the central zone; eyes surrounded by black pigment. Mesosoma: yellowish with minute reddish spotted granulations; tergite VII slightly darker, almost reddish. Metasoma: segment I yellowish; segments II and III reddish-yellow; segments IV and V reddish. Vesicle reddish-yellow; aculeus yellowish at the base and reddish at the tip. Venter yellowish except in sternite VII where it is slightly reddish-yellow. Pectines and genital operculum pale yellow. Chelicerae pale yellow without any dark reticulated spots; fingers yellowish with reddish teeth. Pedipalps: yellowish; granulations on cutting edge of fingers slightly reddish. Legs pale yellow, without any spots.
Morphology. Carapace strongly narrowed anteriorly, almost triangular; anterior margin almost without a median concavity, straight. Carinae almost obsolete; granulations strongly marked by pearl-like granules. Furrows weak. Median ocular tubercle only slightly anterior to the centre of the carapace; median eyes separated by one and half ocular diameters. Three pairs of lateral eyes; the third pair reduced. Sternum triangular, wider than long. Mesosoma: tergites weakly granular. Median carina moderate and present in all tergites; the two lateral carinae vestigial. Tergite VII pentacarinate, but weakly crenulate. Venter: genital operculum of large size divided longitudinally and longer than the sternum. Pectines: pectinal tooth count 8–8; basal middle lamellae of the pectines not dilated; fulcra reduced. Sternites III–VI with thin granulation; granules on VII stronger with two carinae; two lateral furrows present on sternites III–VI. Short semi-slit-like spiracles. Metasoma: segments rounded with ten carinae moderately to strongly marked on segments I to III; carinae on segment III partially fused with the granulations; segments IV and V with only vestigial dorsal carinae and with numerous punctuations. Intercarinal spaces moderately to strongly granular. Telson slightly punctuated, with two small lateral furrows and one ventral carina with a serrula shape; aculeus very short and strongly curved; subaculear tooth absent. Cheliceral dentition characteristic of the family Buthidae: the basal teeth in the movable finger are almost fused [17]. Pedipalps: femur pentacarinate; patella with seven carinae and the internal face without any spinoid granule; chela with vestigial carinae; all faces with thin but intense granulation. Fixed and movable fingers with one linear row of granules divided by some stronger accessory granules; extremity of the fingers with one strong accessory granule giving the shape of forceps to the fingers. Trichobothriotaxy; minorante neobothriotaxy; A-β (beta) for the disposition of the dorsal trichobothria of the femur [18,19]; trichobothria d2 is absent from the femur, and d5 is extremely reduced; patella with seven external trichobothria, and a very reduced d2; chela (hand + fixed finger) with the trichobothria est, Esb, Eb3 absent. Legs: tarsus with a few median fine setae ventrally; pedal spurs moderate on legs III and IV; tibial spurs absent on leg III and reduced on leg IV.
Morphometric values (in mm) of the new species. Total length, 19.7*. Carapace: length, 2.5; anterior width, 1.7; posterior width, 3.6. Metasomal segments. I: length, 1.3; width, 1.6; II: length, 1.6; width, 1.6; III: length, 1.8; width, 1.7; IV: length, 2.0; width, 2.1; V: length, 2.5; width, 2.0; depth, 1.4. Vesicle: width, 0.7; depth, 0.8. Pedipalp: femur length, 2.7, width, 0.8; patella length, 2.9, width, 0.9; chela length, 5.2, width, 0.9, depth, 0.8; movable finger length, 3.4 (* without the vesicle).
Acknowledgements
We are very grateful to Prof. John L. Cloudsley-Thompson, London, for reviewing the manuscript.