

1 Introduction
Un certain nombre de caractéristiques permettent de décrire l'état hydrique du végétal. Elles peuvent être groupées en deux grandes catégories : l'une fait appel à la mesure de la quantité d'eau contenue dans les organes végétaux (TRE), et l'autre à la mesure de l'état énergétique de cette eau (
En conditions de sécheresse, la baisse du potentiel hydrique des plantes induit une importante perte de turgescence au niveau des feuilles [2,3]. Le maintien d'un potentiel hydrique élevé est lié à l'aptitude à extraire l'eau du sol et à la capacité à limiter les pertes d'eau par transpiration [4]. Il caractérise une stratégie d'esquive à la déshydratation [5]. En effet, chez le mil, les mécanismes physiologiques (ajustement osmotique et régulation stomatique) et morphologiques (par exemple la croissance racinaire et la sénescence foliaire) favorisent l'extraction de l'eau du sol ou réduisent les pertes par transpiration [6,7].
Malgré ces réponses physiologiques des plantes à la sécheresse, une diminution continue du potentiel hydrique du substrat finit par se répercuter sur le potentiel hydrique de la plante et sur son rendement [8–10]. Les espèces ou les variétés les plus sensibles montrent alors des valeurs de potentiel hydrique plus négatives que les plantes résistant à la sécheresse.
À l'inverse, les faibles potentiels hydriques observés chez le blé dur soumis à un stress hydrique sont associés à une tolérance à la déshydratation liée non seulement à la précocité de son cycle, mais également à sa capacité d'ajustement osmotique [11]. La capacité de maintenir une TRE élevée est donc considérée, dans cette approche, comme la meilleure représentation de l'ajustement osmotique. La teneur relative en eau au niveau de la plante peut varier entre 68% et 84% dans les bonnes conditions hydriques, et de 45% à 63% en présence d'un déficit hydrique [12].
Pour le cas du mil, il a été constaté [13] que cette céréale perd sa turgescence pour des valeurs importantes de la TRE (TRE > 80%) mais garde un volume d'eau liée élevé (40% de la TRE). De même, le potentiel osmotique du mil reste très bas (environ −2,5 MPa) pour des TRE importantes, indiquant que cette espèce retient une importante quantité d'eau sous l'effet de la déshydratation. Il a aussi montré que le potentiel de turgescence varie considérablement pour de faibles variations de la TRE. Ainsi, nous pouvons résumer les objectifs de ce travail comme suit :
- – voir l'influence du manque d'eau sur l'évolution du potentiel hydrique foliaire et la teneur relative en eau ;
- – étudier l'influence des caractéristiques hydriques sur le rendement des divers écotypes ;
- – chercher une variabilité génétique au sein des écotypes de mil pour la résistance à la sécheresse.
2 Matériel et méthodes
2.1 Matériel végétal
Le matériel végétal est composé de six écotypes autochtones de mil, appartenant à des étages bioclimatiques différents (Tableau 1).
Caractéristiques climatiques des zones de collecte des différents écotypes. Données de l'Institut national de météorologie (INM), 2006, Tunisie
| Écotype | Origine | Étage bioclimatique | Précipitation moyenne annuelle (mm) | Température moyenne annuelle (°C) | Température moyenne du mois de janvier (°C) | Température moyenne du mois de juillet (°C) |
| HG | Hamam Laghzaz ; Nabeul | Semi-aride | 400–600 | 18,3 | 12,1 | 26,0 |
| AM | El Amra ; Sfax | Aride moyen | 200–400 | 21,8 | 11,2 | 25,9 |
| EC | Echabba ; Mahdia | Aride supérieur | 200–400 | 19,5 | 11,9 | 26,3 |
| KS | Kairouan Sud ; Kairouan | Aride moyen | 200–400 | 19,9 | 11,5 | 28,7 |
| D | Île de Djerba ; Médnine | Aride inférieur | 100–200 | 21,3 | 12,4 | 26,9 |
| ZZ | Zarzis ; Médnine | Aride inférieur | 100–200 | 22,7 | 12,7 | 27,3 |
2.2 Protocole expérimental
Les semis ont été réalisés le 23 mai 2002 à la station expérimental de l'Inra de Tunis (altitude : 10 m ; latitude : 36°51′ ; longitude : 10°11′). Le climat est de type semi-aride, à hiver doux.
Le sol est d'apport alluvial ; il est profond, homogène et de texture argileuse, avec une teneur en éléments fins >70%. Les propriétés physiques et chimiques du sol de l'InraT sont résumées dans le Tableau 2.
Propriétés physiques, chimiques et hydriques du sol de l'InraT
| Propriétés physiques (% de matière sèche) | Composition chimique (%) | Caractéristiques hydriques (%) |
| Argile = 16 | Calcaire actif = 13 | C.C. = 22 |
| Limon fin = 19,5 | Azote total = 0,08 | P.F.P. = 18 |
| Limon grossier = 14 | P2O5 total = 4 | R.E.U. = 4 |
| Sable fin = 27,5 | P2O5 assimilable = 0,01 | |
| Sable grossier = 20 | K2O total = 3,87 | |
| K2O assimilable = 0,94 | ||
| pH = 8,2 |
Chaque répétition est subdivisée en six parcelles élémentaires de 4 m2 de superficie. Chaque carré (correspondant à un écotype) est formé de quatre lignes de 2 m de longueur. Les lignes sont espacées de 33 cm et, sur la ligne, deux plants sont distants de 30 cm.
Les traitements sont espacés de 3 m et sont entourés dans toutes les directions de lignes de bordure.
L'eau d'irrigation est délivrée aux parcelles par submersion.
En adoptant les valeurs de l'évaporation de référence mesurées à la station de Tunis au niveau des cases lysimétriques [14] et celles des coefficients culturaux déterminés selon la méthode de la FAO [15], nous avons estimé les besoins théoriques en eau (ETM) du mil à 580 mm (100%), la fréquence d'irrigation étant la même (10 j au début de la culture et une fois par semaine dès le début de l'épiaison).
Les traitements appliqués sont :
- • T100, témoin irrigué à 100% ETM.
- • T70, traitement recevant 70% ETM.
- • T35, traitement irrigué à 35% ETM.
L'eau d'irrigation utilisée est celle courante, titrant 1 g/l en NaCl.
Le dispositif statistique expérimental est en Split Plot à trois répétitions, avec comme facteur principal la quantité d'irrigation et, comme facteur secondaire, les écotypes de mil.
3 Mesures réalisées
Le potentiel hydrique foliaire (
La teneur relative en eau (TRE) a été calculée selon la formule suivante :
Le temps de saturation pour le mil est de 4 h au minimum [17]. L'hydratation des feuilles drapeaux a lieu à l'obscurité et à température ambiante, durant 12 h. Le poids sec est déterminé après passage à l'étuve à 80 °C pendant 48 h. Les mesures de la TRE ainsi que du potentiel hydrique foliaire ont été réalisées à midi sur les feuilles drapeaux et à raison de quatre répétitions par écotype et par traitement.
À maturité, les chandelles sont battues et le rendement est estimé par qx/ha.
4 Étude statistique
L'analyse statistique a été réalisée à l'aide du logiciel STATIT-ITCF. Les moyennes sont comparées par la plus petite différence significative (ppds) à l'aide du test de Bonferonni au seuil de 5%.
5 Résultats et discussion
5.1 Variation du potentiel hydrique foliaire sous stress hydrique
Les résultats de la variation du potentiel hydrique foliaire en fonction des écotypes et des traitements sont présentés dans la Fig. 1. Cette figure montre que, chez tous les écotypes étudiés, l'augmentation de l'intensité du stress s'accompagne d'un abaissement du potentiel hydrique foliaire par rapport au témoin. En revanche, chez les plantes témoins, aucune différence significative entre les potentiels hydriques foliaires n'a été constatée.
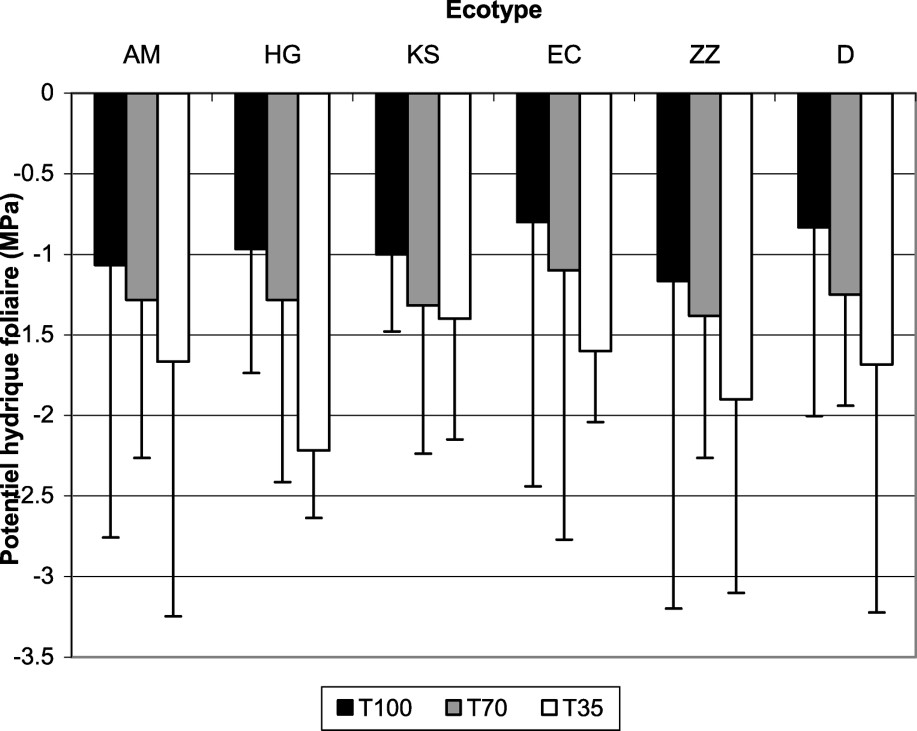
Variation du potentiel hydrique foliaire en fonction des six écotypes de mil et des trois traitements hydriques.
La baisse du potentiel hydrique est d'autant plus accentuée que l'intensité de la contrainte hydrique appliquée est plus sévère. En effet, le potentiel hydrique chute de 30% lorsqu'on passe du témoin au traitement modéré (T70) et de 80% lors de l'application du stress sévère (T35). Face à ce dernier traitement, ce sont les écotypes HG, D et EC qui ont été les plus touchés, avec des augmentations de potentiel (en valeur absolue) de plus de 100%. Ce sont en fait les populations qui possèdent des
La chute du potentiel hydrique foliaire du mil, consécutive à l'application du stress hydrique, a été attribuée par certains auteurs [18] aux diminutions des potentiels osmotiques et de turgescence. Nous avons montré également qu'en absence de stress hydrique, le potentiel hydrique foliaire est sensiblement identique pour les accessions étudiées. Ces faits ont été déjà trouvés sur le mil [19].
En présence de déficit prononcé, les potentiels hydriques foliaires de HG, D et EC subissent des abaissements de plus de 100% (par rapport à leur témoin respectif), ce qui leur permettrait de garder un gradient de potentiel hydrique important entre les tissus et le milieu de culture, leur assurant une meilleure alimentation en eau.
En situation de contrainte hydrique, le potentiel hydrique foliaire est d'autant plus bas que l'écotype de mil est sensible [18].
Lorsque la diminution du potentiel hydrique foliaire est élevée (>100%), les variétés sont caractérisées par un ajustement osmotique de type adaptatif [20], comme c'est le cas des écotypes HG, D et EC. Ils sont capables de moduler leur potentiel osmotique et, par conséquent, leur potentiel hydrique en fonction de la disponibilité de l'eau dans le sol. Une baisse pareille de
Le maintien d'un
Une variabilité génétique importante a été mise en évidence en ce qui concerne l'évitement de la déshydratation. Chez le sorgho, par exemple, on a noté des potentiels hydriques foliaires, dans les conditions du champ, compris entre −1,34 et −1,83 MPa [23].
5.2 Variation de la teneur relative en eau sous stress hydrique
L'état hydrique des feuilles paniculaires chez les écotypes de mil soumis à un déficit hydrique est illustré sur la Fig. 2. L'analyse statistique montre que les effets de ces différents facteurs sont significatifs. Toutefois, aucune interaction significative n'a été trouvée.

Variation de la teneur relative en eau en fonction des six écotypes de mil et des trois traitements hydriques.
L'allure générale de la figure montre, chez tous les écotypes, que la TRE diminue proportionnellement à l'intensité du stress hydrique appliqué. Elle passe de 84% chez le témoin à 80% pour le traitement modéré et à 76% pour la contrainte sévère, soit des réductions respectives de 4,76% et de 9,52%.
En comparant les accessions indépendamment des traitements hydriques, nous constatons que c'est la population de Kairouan (KS) qui présente la teneur relative en eau la plus élevée. Elle est suivie par AM et ZZ, puis du groupe formé par les écotypes D, EC et HG. Il est à signaler que ce dernier écotype (HG) est caractérisé par la TRE la plus faible.
L'abaissement régulier de la TRE en présence de contraintes hydriques chez les six écotypes de mil a été signalé par de nombreux auteurs sur plusieurs plantes, telles Vigna unguiculata et Phaseolus vulgaris [24] et le blé dur [11]. Ce dernier auteur a attribué ce phénomène à la diminution du potentiel osmotique.
La comparaison de l'état hydrique des six écotypes de mil a montré des différences significatives entre les diverses accessions et a révélé la supériorité de l'écotype KS par rapport à HG. La présence d'une variabilité génétique pour la TRE a été signalée par ailleurs, pour des variétés de blé tendre [25].
Les écotypes de mil ont gardé en moyenne une TRE de 80% en présence d'un stress hydrique. Cette teneur en eau relativement élevée, indique que le mil est une plante de type « non sénescent » ou stay green, autrement dit qui conserve des feuilles vertes, succulentes et photosynthétiquement actives, même en présence de contraintes abiotiques [26]. En effet, une TRE élevée et une turgescence positive peuvent être nécessaires pour soutenir l'activité cellulaire près du niveau normal. À cet égard, on considère qu'une plante dont la TRE est comprise entre 68% et 84%, est une plante bien hydratée [12].
Le maintien d'une teneur élevée en eau dans les feuilles en croissance et dans les feuilles en expansion, sous stress hydrique, indique l'efficacité de l'ajustement osmotique, qui contrebalance la diminution du potentiel hydrique, sans diminution significative de la TRE [27]. Ce manque d'eau, se traduisant au niveau de la plante par une diminution de la TRE ne dépassant guère les 10% pour le stress hydrique le plus sévère (Fig. 3), pourrait indiquer une grande capacité d'ajustement osmotique [28]. Également, la stabilité du rendement de l'orge et la tolérance de cette espèce sont associées au maintien de la TRE à des
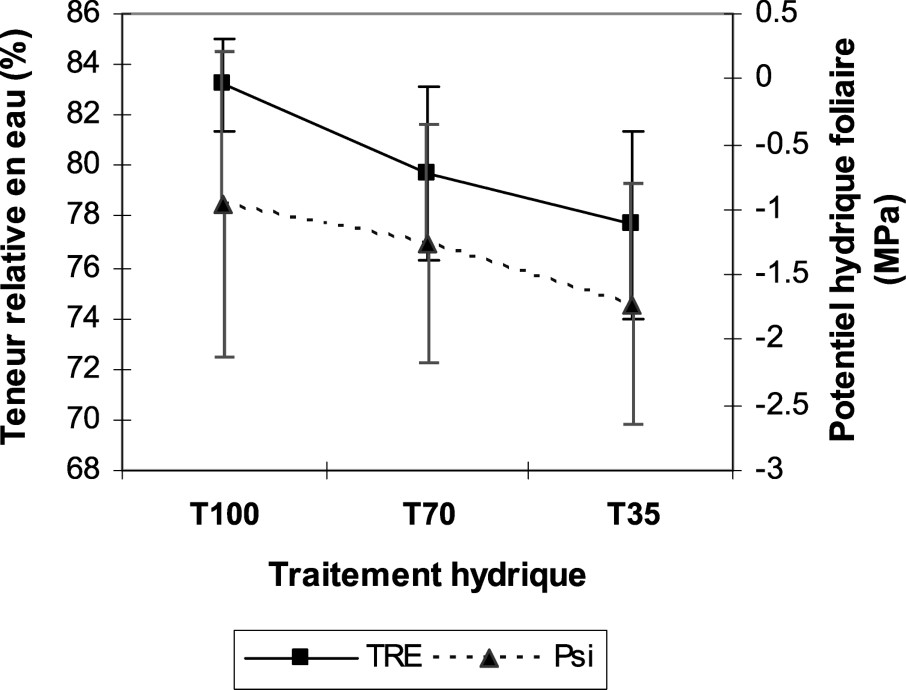
Variation du potentiel hydrique foliaire et de la teneur relative en eau en fonction des trois traitements hydriques.
Par ailleurs, la comparaison de l'état hydrique des six écotypes de mil étudiés montre que ces populations ont gardé pratiquement le même classement quant à leur TRE sous les différents stress appliqués. Ce classement met en valeur la supériorité de l'écotype KS, qui affiche la TRE la plus importante (environ 85%). Il est suivi de près par les accessions AM et ZZ (TRE > 80%). Les écotypes D, EC et HG ont des TRE < 80% (variant entre 74% et 80%).
5.3 Variation du rendement en grains sous stress hydrique
La Fig. 4 représentant l'estimation des rendements en qx/ha, calculés à partir des rendements par mètres carrés montre que seule la dose très restrictive a réduit de façon significative le rendement. La baisse de rendement dans ce cas est de 29%.

Variation du rendement en grains en fonction des six écotypes de mil et des trois traitements hydriques.
Cette figure montre également que KS et ZZ sont les accessions les plus productives en absence de stress hydrique et enregistrent des rendements supérieurs à 80 qx/ha (aux alentours de 100 qx/ha pour KS), alors que HG et D arrivent en seconde place, avec des rendements moyens de 70 qx/ha. Au dernier rang, nous retrouvons AM et EC produisant en moyenne entre 65 et 70 qx/ha. De plus, les résultats de l'étude montrent que les écotypes HG, EC et AM ont enregistré une légère amélioration de leur rendement sous stress hydrique modéré (amélioration de 4% pour le premier, 14% pour le second et 12% pour le troisième). Lorsque le stress est plus intense, KS et ZZ sont les plus affectés et se montrent les plus sensibles au manque d'eau, puisque la perte de rendement dans ce cas est voisine des 50% (47%). La chute de rendement est de 34% pour l'écotype D et elle tourne aux alentours de 27 à 30% pour les populations AM, EC et HG.
La baisse du rendement chez le mil, obtenue en présence du déficit hydrique sévère, semble consécutive [29] à la diminution de toutes les composantes du rendement, particulièrement les caractéristiques de la chandelle principale (poids, longueur, rendement en grains) et aussi à la réduction des dimensions relatives aux descripteurs de la structure de la plante (surface foliaire paniculaire, hauteur de la tige principale, nombre de talles).
De nombreux auteurs ont signalé la réduction du rendement de mil sous stress hydrique sévère [30–34]. D'après Balaños et Eadmeades [35], Eadmeades et al. [36], Andrade et al. [37] et Setter et al. [33], cette chute de rendement est causée à la fois par la diminution du nombre de chandelles récoltables à maturité et par une diminution de la taille des chandelles restantes. En revanche, Setter et al. [33] l'expliquent par une augmentation de la teneur en ABA. Ils attribuent à cet acide un rôle important dans la perte de rendement, puisque son accumulation inhibe la photosynthèse par fermeture stomatique [38]. De récentes études ont montré que l'invertase est l'enzyme responsable de l'inhibition du remplissage des grains durant un stress hydrique [39].
D'autres auteurs, Bidinger et al. [8,9] attribuent la baisse de rendement en situation contraignante à la perte de matière sèche des tiges, qui correspond vraisemblablement à la mobilisation d'une partie de la matière sèche des tiges pour le remplissage des épis. À l'instar de nos propres résultats, Mahalakshmi et al. [31] ont obtenu des réductions de rendement de l'ordre de 30 à 40%. De même, Do et Winkel [10] ont trouvé que la baisse de rendement variait entre 14 et 40% pour deux écotypes de mil soumis à un stress hydrique.
Au contraire, lorsque la contrainte hydrique est modérée, le rendement n'est pas compromis. Ce résultat est corroboré par ceux de Yazar et al. [40] et de Crépin [41]. Au contraire, un effet bénéfique de ce traitement a été observé au niveau du rendement en grains de certains écotypes.
Ces différences comportementales des divers écotypes de mil en situation de sécheresse ont été signalées par Fussel et al. [42]. En effet, les six écotypes de mil se sont départagés en groupes à comportements différents. Ainsi, les populations tardives KS et ZZ, les plus vigoureuses et les plus productives aussi bien en contexte irrigué qu'en situation de stress, ont été les plus pénalisées par le manque d'eau. Dijak et al. [43] ont justement signalé que le rendement était supérieur chez les variétés à tige longue plutôt que courte. Budak et al. [44] ont également trouvé que les réductions en termes de rendement touchaient davantage les variétés tardives que celles précoces.
Le groupe réunissant les écotypes EC et AM est caractérisé par un comportement moyen sur tous les plans. En effet, ces écotypes sont les moins productifs en l'absence de stress, mais les moins compromis en conditions contraignantes. D'ailleurs, Blum [45] a suggéré l'existence d'une relation positive entre la longueur du cycle et le rendement potentiel. Le recours à une stratégie d'évitement conduit ainsi à « sacrifier » une part de la productivité.
Ce genre de comportement a été signalé par Budak et al. [44], qui ont montré que les variétés semi-naines de blé présentaient une meilleure stabilité du rendement comparativement aux variétés à haute taille.
5.4 Corrélations
Des corrélations positives entre le rendement et le potentiel hydrique foliaire, d'une part, et le rendement et la teneur relative en eau, d'autre part, ont été trouvées (Tableau 3). En effet, les écotypes KS, ZZ et AM, caractérisés par des potentiels hydriques et des TRE élevés, sont les plus productifs, même en situation de sécheresse.
En étudiant l'effet de la contrainte hydrique sur deux variétés de blé dur, Slama [46] a constaté que la variété ayant le potentiel le plus bas est la plus sensible à la sécheresse et que c'est elle qui a donné le rendement en grains le plus faible. Ce résultat est corroboré par Clarke et Mc Caig [47] et Jardat et Konzaz [48]. Tous ces résultats plaident en faveur de l'importance des caractéristiques hydriques de la feuille drapeau dans la détermination du rendement. Ils sont plus importants que les caractéristiques morphologiques de la plante [49]. D'ailleurs, la feuille étendard est l'organe le moins sensible au manque d'eau, d'ou l'importance de cet organe dans la détermination du rendement en cas de déficit hydrique [49]. Ainsi, un potentiel hydrique et une teneur en eau élevés agissent positivement sur le rendement. En effet, les écotypes KS et ZZ, caractérisés par des caractéristiques hydriques très importantes au niveau de la feuille drapeau, ont donné les meilleurs rendements.
6 Conclusion relative à l'état hydrique du mil
Pour caractériser le statut hydrique d'une plante, on fait couramment appel à un paramètre qui découle du potentiel chimique de l'eau : le potentiel hydrique.
La TRE est elle aussi généralement utilisée pour estimer le statut hydrique d'une plante soumise à un stress hydrique. L'évolution de ce paramètre rend compte non seulement des variations de la quantité d'eau présente dans les tissus, mais aussi des modifications de leur capacité à incorporer l'eau en condition de déficit hydrique.
Les mesures simultanées de ces deux paramètres permettent, en plus, grâce aux courbes de variation de la TRE en fonction du potentiel hydrique, d'évaluer l'aptitude de la plante à réaliser un ajustement osmotique actif.
Les études effectuées sur les six écotypes de mil soumis à trois régimes hydriques (témoin, modéré et sévère) ont révélé deux comportements types, qui sont :
- • un effet variétal du stress hydrique, se traduisant par une réduction importante du potentiel hydrique foliaire (>100%) en parallèle avec la réduction la plus élevée de la TRE, traduisant ainsi une chute de la turgescence cellulaire, une absence d'ajustement osmotique actif et la présence d'un ajustement osmotique adaptatif. C'est le cas des populations HG, D et EC ;
- • un effet variétal du stress hydrique, se traduisant par une faible réduction du potentiel hydrique foliaire, tout en maintenant une TRE sensiblement plus élevée que celle des autres écotypes : la réduction du potentiel hydrique pourrait, dans ce cas, être le résultat d'une réduction importante du potentiel osmotique due à une accumulation active de solutés. C'est le cas des populations KS, AM et ZZ.
Nos résultats ont également montré que le potentiel hydrique foliaire et la TRE évoluent de la même manière, dans le sens d'une diminution suite à l'application d'une contrainte hydrique. Ces caractéristiques hydriques sont également très importantes, puisqu'elles conditionnent le rendement. De plus, la variabilité génétique du matériel végétal vis-à-vis du stress hydrique ne semble pas liée à l'origine géographique des écotypes de mil, mais plutôt à leur phénologie et à leur morphologie.