

1 Introduction
L'extension de l'agriculture irriguée et l'utilisation intensive des ressources en eau, combinées à une forte évaporation dans les régions arides et semi-arides, conduit inévitablement à la salinisation des sols et des nappes d'eau. Dans de nombreuses régions semi-arides, et en particulier en Tunisie, cette situation est aggravée par la raréfaction des ressources en eau douce. En effet, les eaux considérées comme saumâtres titrant entre 2 et 6 g de sels par litre constituent 50% des eaux souterraines dans le Sud de la Tunisie. En agriculture irriguée, les eaux provenant de barrages présentent une charge en sel de 2 à 3 g/l, alors que celle des puits titrent de 4 à 7 g/l [1].
La valorisation de ces eaux en agriculture se heurte à plusieurs difficultés, et en particulier au degré de tolérance au sel des espèces cultivées, qui sont pour la plupart des glycophytes sensibles [2].
L'approche alternative consiste à faire appel à des plantes qui ont une certaine tolérance aux sels, c'est-à-dire dont les rendements restent satisfaisants face à des niveaux de sels élevés.
Le choix de ces espèces tolérantes aux sels constitue un des moyens de préserver ces sols et offre la possibilité de valoriser ces eaux saumâtres et les régions marginales affectées par la salinité. Dans ce contexte, la recherche d'une variabilité intraspécifique est aussi recherchée et ouvre des perspectives d'amélioration variétale des cultures.
Bien que, depuis fort longtemps, le comportement en milieu salé des plantes cultivées ait fait l'objet de nombreux travaux, le mil tunisien (Pennisetum glaucum (L.) R. Br.) n'a pas reçu une attention particulière. Or, cette céréale estivale conduite en irrigué, qui sert à la fois à l'alimentation humaine et animale grâce à sa valeur nutritionnelle [3] est vivement indiquée, en raison de son importante résistance aux sels [4,5].
En Tunisie, au contraire d'autres pays d'Afrique, le mil ne constitue pas l'alimentation de base des populations rurales [6]. Pourtant, il occupe une part très importante des surfaces mises en culture chaque année dans le Centre et le Sud du pays, ainsi que sur les régions côtières [7].
Le premier impératif d'une culture satisfaisante consiste en son implantation convenable, et donc la réussite de la phase de germination est déterminante dans tout le reste de la période végétative. De plus, la réponse des semences à la salinité pourrait être un indicateur de la tolérance des plantes au sel pour les stades ultérieurs du développement [8], d'où l'intérêt de notre étude, dont les objectifs principaux sont :
- • l'identification d'écotypes de mil pouvant s'accommoder le mieux des eaux saumâtres et qui pourraient constituer un matériel végétal alternatif pour les zones marginales ;
- • la recherche d'éventuelles corrélations pouvant lier le comportement au stade germination à celui de stades ultérieurs, et qui pourraient constituer un test rapide à cet égard.
2 Matériel et méthodes
2.1 Matériel végétal
Cet essai a porté sur six écotypes autochtones de mil collectés à travers le territoire tunisien. Ces écotypes appartiennent à des étages bioclimatiques différents, sont cultivés sur des sols de textures diverses et sont irrigués avec des eaux de qualité médiocre. Ces écotypes sont :
- • D : écotype originaire de l'île de Djerba (latitude , longitude ). Il est précoce, à tige courte et possède des chandelles petites, légères et peu productives ;
- • ZZ : écotype collecté à Zarzis (latitude , longitude ), présentant une longue tige et des chandelles fines à structure lâche. Son cycle est assez long, et il est productif ;
- • HG : écotype collecté au cap Bon (Hammam Laghzaz : latitude , longitude ) possédant une courte tige et des chandelles larges et compactes. C'est un mil à cycle court et moyennement productif ;
- • KS : mil de la plaine de Kairouan (latitude , longitude ), très vigoureux, à paille haute, à chandelles de fort calibre très productives. C'est un écotype à long cycle ;
- • AM : écotype provenant de la zone de El Amra (Sfax : latitude , longitude ). Il est du type mixte (précoce à semi précoce), avec des chandelles larges et assez productives ;
- • EC : mil collecté à Echaba (Mahdia : latitude , longitude ). C'est un écotype moyen, qui présente à la fois les caractéristiques des écotypes à cycle long (tige longue) et celles des écotypes à cycle court (chandelles peu productives et peu vigoureuses). Il est semi précoce et moyennement productif.
2.2 Protocole expérimental
Les essais ont été conduits au laboratoire et au champ.
Au laboratoire, il a été procédé à des essais concernant l'effet du chlorure de sodium (NaCl) sur le taux de germination et sur les longueurs de l'épicotyle et des racines des différents écotypes étudiés. Pour chaque écotype, les grains sont lavés à l'eau du robinet, désinfectés pendant quelques minutes dans de l'eau de Javel, puis rincés à l'eau distillée. Ils sont placés à raison de 50 grains dans des boites de Pétri sur du papier filtre imbibé avec, soit 20 ml d'eau du robinet (témoin, titrant environ 1 g/l), soit des solutions salines contenant respectivement 3 et 6 g de sel dilué dans l'eau du robinet. Ces traitements peuvent être résumés comme suit :
Ces concentrations sont choisies parce qu'elles sont proches des charges en sel des eaux utilisées pour l'irrigation du mil dans le sud tunisien. Chaque traitement a été répété cinq fois.
Après une semaine d'incubation à l'obscurité et à 30 °C, nous avons compté les graines germées et mesuré les longueurs respectives de leur épicotyle et de leur radicule.
Les essais en plein champ ont été conduits dans une zone semi aride à hiver doux. Les semis ont été réalisés le 23 mai 2005 à la station expérimentale de l'Inra de Tunis (altitude : 10 m ; latitude : ; longitude : ).
Le sol est d'apport alluvial ; il est profond, homogène et de texture argileuse, avec une teneur en éléments fins >70%.
L'eau d'irrigation utilisée est celle courante, titrant 1 g/l en NaCl. Elle est délivrée aux parcelles par submersion.
Pour réaliser le stress salin, nous avons adopté un dispositif en Split Plot permettant de donner à l'eau d'irrigation la concentration en sel souhaitée et qui est la même que celle adoptée pour les essais de germination au laboratoire ().
Les besoins théoriques en eau du mil sont estimés à 580 mm (ETM = 100%) [8,9].
La fréquence d'irrigation était la même : 10 jours au début de la culture et une fois par semaine dès le début de l'épiaison. Les traitements ont été démarrés au stade quatre feuilles (deux fois 27 mm au début, puis sept fois 75 mm d'eau).
Les différents écotypes ont été semés en cinq répétitions. Chaque répétition est représentée par un carré de 25 m2. La densité de semis est de 200 000 plants/ha. Après démariage, la densité devient de 10 plantes au mètre carré.
Les traitements sont espacés de 3 m et sont entourés, dans toutes les directions, de lignes de bordure.
3 Mesures de la croissance des plantes
- • La mesure de la hauteur des plantes (HAT), en centimètres, est déterminée à l'aide d'une règle graduée du collet de la plante à l'insertion de la chandelle, au stade maturité.
- • La mesure de la surface foliaire de la feuille drapeau ou feuille paniculaire (SFD), en centimètres carrés, a été déterminée à l'aide d'un planimètre CI-202 au stade pâteux.
4 Mesures des composantes de rendement
À la récolte, les chandelles principales (100 chandelles par placette) sont coupées, mesurées, pesées, puis battues.
Les caractéristiques propres à la chandelle principale sont :
- • la longueur de la chandelle (LOC), en centimètres ;
- • le rendement en grains de la chandelle (RGC), en grammes.
5 Analyse statistique
Les résultats sont soumis à l'analyse de la variance à un ou à deux facteurs, et les moyennes sont comparées selon la méthode de Bonferonni, basée sur la plus petite différence significative.
Chaque moyenne est affectée d'une lettre, les moyennes suivies d'une même lettre n'étant pas significativement différentes.
6 Résultats et discussion
6.1 Résultats des essais de germination (laboratoire)
6.1.1 Taux de germination
Le Tableau 1 montre que, quel que soit l'écotype, le taux de germination des semences stressées est supérieur à celui des graines non contraintes (quoique de façon non significative pour la dose ). Cette amélioration est d'environ 12% pour la salinité modérée, et ce quel que soit l'écotype considéré.
Taux de germination
| Traitement | Taux de germination en % |
| 89,3 b | |
| 100 a | |
| 89,8 b |
L'analyse de la variance n'a pas montré de différence significative pour le taux de germination des six écotypes de mil (Tableau 2).
Analyse de variance relative à l'effet du NaCl sur la germination des différents écotypes de mil
| Source de variation | Taux de germination | Longueur de l'épicotyle | Longueur de la radicule | |
| Écotype | ddl | 5 | 5 | 5 |
| Test F | 17,31ns | 1,07 ns | 4,53⁎⁎ | |
| ppds | ns | ns | 3,09 | |
| Traitement | ddl | 2 | 2 | 2 |
| Test F | 113,44⁎⁎ | 62,45⁎⁎ | 121,99⁎⁎ | |
| ppds | 2,05 | 0,37 | 1,39 | |
| Ecotype × traitement | ddl | 10 | 10 | 10 |
| Test F | 11,65⁎⁎ | 0,43 ns | 4,40⁎⁎ | |
| ppds | 5,01 | ns | 3,42 | |
| CV (%) | 2,6 | 7,3 | 11,4 |
⁎⁎ Seuil de signification 5%.
La stimulation de la germination par le sel a été mentionnée par de nombreux auteurs sur le mil [4,10]. D'ailleurs, le trempage préalable des semences de nombreuses cultures dans une solution saline augmente la germination, l'installation des plantules et, dans certains cas, stimule la croissance et le rendement [11–13].
6.1.2 Longueur de l'épicotyle
Une concentration en sel de 4 g/l () a amélioré de façon significative (l'épicotyle a gagné en moyenne 2 cm en longueur) la croissance de l'épicotyle chez tous les écotypes (Fig. 1). Des observations similaires ont été rapportées [14,15].
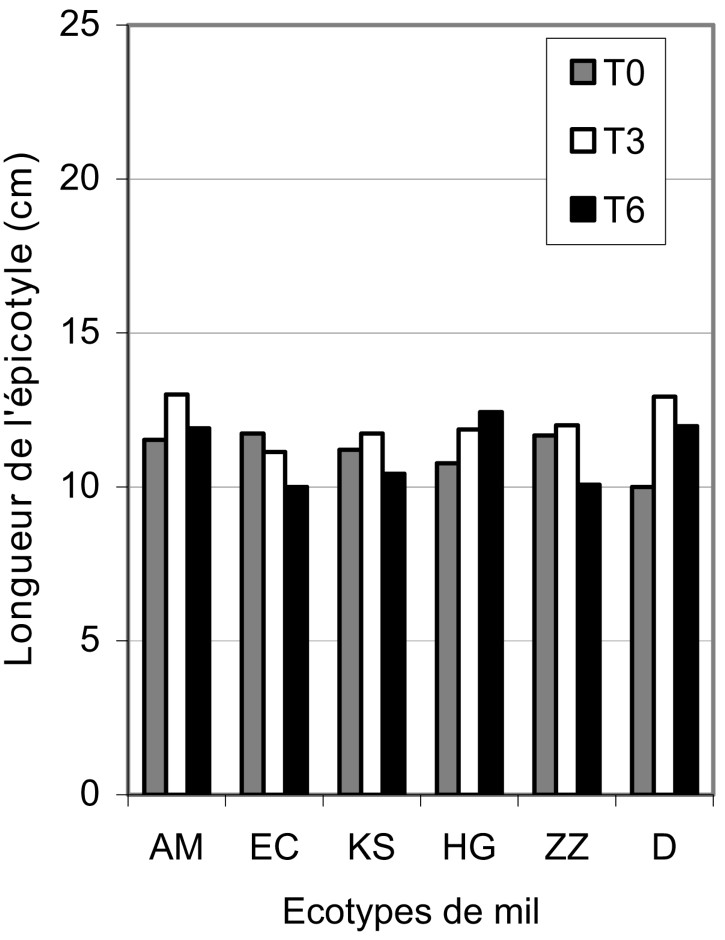
Variation de la longueur de l'épicotyle des différents écotypes de mil en fonction en fonction du stress salin.
Lorsque le traitement est sévère (), l'écotype HG montre une réponse d'autant plus positive que la concentration en sel est élevée (Fig. 1). Il est suivi des écotypes D, AM et KS, qui constituent une classe homogène gardant la même longueur du coléoptile, quelle que soit la concentration en sel. Au bas de l'échelle se situent les écotypes ZZ et EC, qui réagissent négativement dès que la concentration en sel dépasse 4 g/l. Ces résultats sont en accord avec ceux de Ben Naceur et al. [16], qui ont montré un effet variable de la salinité sur l'épicotyle de nombreuses variétés de blé.
6.1.3 Longueur de la radicule
Le résultat de l'étude de la croissance radiculaire sous stress salin est illustré par la Fig. 2. Notons que, plus la concentration en sel augmente, plus la radicule est longue. En effet, cette augmentation est de 21% dans le cas du stress modéré et de 51% pour le stress sévère. De plus, certains écotypes, comme AM, HG, D et EC, se distinguent par leur allongement radiculaire dans le même sens que la charge en sel. Dans le cas de ces écotypes, les allongements radiculaires varient en moyenne entre 29 et 76% pour le traitement et entre 65 et 153% pour le traitement .
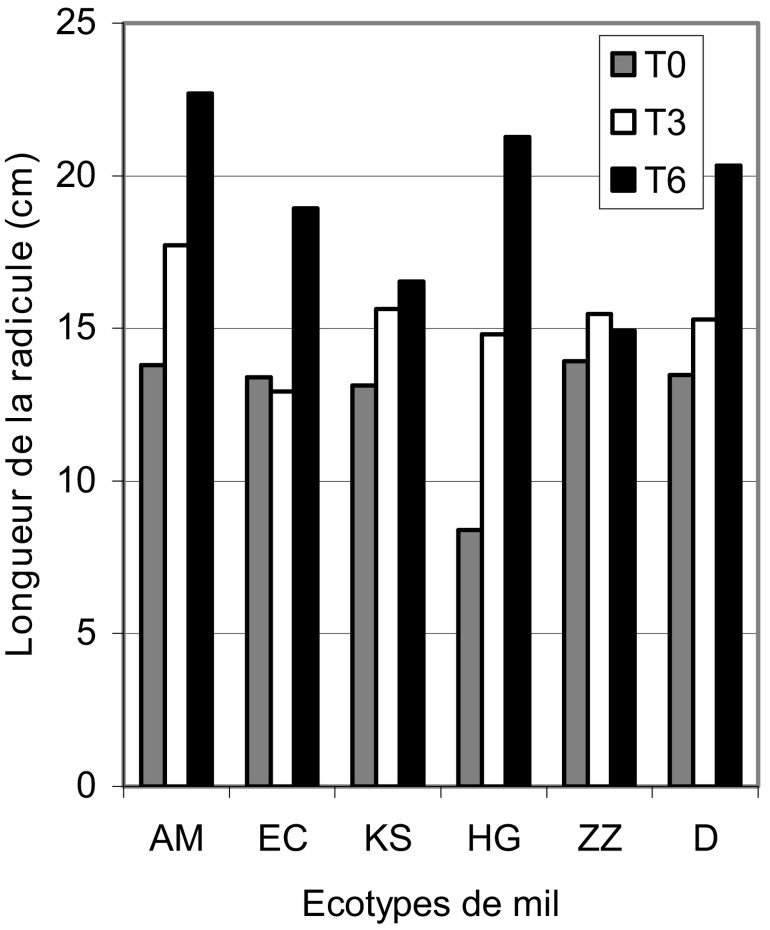
Variation de la longueur de la radicule des différents écotypes de mil en fonction en fonction du stress salin.
Un deuxième groupe, formé par les écotypes KS et ZZ, garde la même dimension radiculaire, quelle que soit l'intensité du traitement salin.
Ce comportement particulier des écotypes AM, HG, D et EC, serait une forme de tolérance au sel. La stimulation de la croissance des racines par NaCl a été décrite chez d'autres plantes, particulièrement tolérantes au sel [17–19].
6.1.4 Corrélations entre salinité et germination
Pour expliquer le comportement germinatif des différents écotypes de mil en présence de sel, nous avons cherché l'existence d'éventuelles corrélations pouvant lier la germination à la croissance du coléoptile et de la radicule. En effet, en présence de salinité, la corrélation trouvée est significative, comme le montre le Tableau 3. Elle est égale à (). Nous pouvons donc dire que, sous contrainte saline, la germination semble dépendre beaucoup plus de la radicule. En effet, la germination et l'installation des plantules sont facilitées lorsque les écotypes présentent de longues radicules.
ANOVA de la corrélation entre le taux de germination et la longueur du coléoptile et de la radicule en conditions de stress
| Source | ddl | CM | F |
| Totale | 53 | 734,44 | 68,73⁎⁎ |
| Régression | 1 | 89,19 | |
| Résiduelle | 52 |
⁎⁎ Seuil de signification 5%.
6.1.5 Conclusion
L'étude effectuée au laboratoire a montré un effet peu marqué du stress salin sur le taux de germination et sur l'émergence du coléoptile. En revanche, il est plus significatif pour la croissance radiculaire, avec des différences entre écotypes, confirmant ainsi d'autres travaux [13,17,20] qui ont montré un effet similaire du stress salin sur la croissance de nombreuses espèces végétales et d'autres graminées céréalières ou fourragères. Cette étude a également confirmé l'adaptation à la salinité du mil et sa tendance à l'halophilie, du moins au stade germination. Un tel résultat a été signalé par Bouraïma et al. [4] sur un mil de la région de Zarzis et par de Villèle [10] sur un mil pakistanais. De plus, tous les écotypes de mil étudiés se sont montrés égaux devant la salinité quant à leur taux de germination. En revanche, des différences génotypiques radiculaires ont révélé que AM, HG, D et EC sont tolérants au sel, alors que les écotypes ZZ et KS sont résistants.
Ces résultats peuvent nous aider à exploiter cette variabilité lors de l'usage d'eaux saumâtres pour la culture du mil, à condition de les vérifier aux stades ultérieurs de développement.
6.2 Résultats des essais de rendement (champ)
6.2.1 Effet du stress salin sur la croissance
Croissance en hauteur. La comparaison des données pour la hauteur des tiges ne montre pas de différences significatives entre les traitements salins, mais plutôt un effet de type écotype (Tableau 4). En effet, la barre de 2 m constitue la limite qui scinde les six écotypes de mil en deux groupes, le premier renfermant, par ordre décroissant, KS et ZZ, et le deuxième étant constitué par EC, AM, HG et D (Tableau 5).
Analyse de variance relative à l'effet du NaCl sur la hauteur des différents écotypes de mil
| Source de variation | Taux de germination | |
| Ecotype | ddl | 5 |
| Test F | 25,17⁎⁎ | |
| ppds | 18,09 | |
| Traitement | ddl | 2 |
| Test F | 1,57 ns | |
| ppds | ns | |
| Ecotype × traitement | ddl | 10 |
| Test F | 1,74 ns | |
| ppds | ns | |
| CV (%) | 4,1 |
⁎⁎ Seuil de signification 1%.
Classification des écotypes de mil en groupes homogènes pour la hauteur des tiges (cm)
| Écotype | Moyenne |
| KS | 221,89 a |
| ZZ | 217,36 a |
| EC | 190,89 b |
| AM | 190,78 b |
| HG | 188,14 b |
| D | 182,00 b |
Croissance de la surface foliaire. La Fig. 3 montre que, du point de vue génotypique, c'est l'écotype D qui possède la feuille drapeau la plus petite. Par ailleurs, les autres écotypes présentent des feuilles statistiquement similaires. Sous l'effet de la contrainte saline, la surface foliaire paniculaire de tous les écotypes, sans exception, est significativement augmentée. Cette augmentation est d'autant plus importante que la charge en sel de l'eau d'irrigation est importante. Elle est respectivement en moyenne de 17% pour le traitement , et de 32% pour le traitement par rapport au témoin. La stimulation de la croissance (tige, feuille paniculaire) observée chez le mil tunisien a été signalée par Bouraïma [4], mais aussi chez le sorgho [21] et le triticale [22,23]. Elle serait due à l'effet bénéfique de Na+ sur l'expansion cellulaire et la balance hydrique [24] ou au maintien de la croissance par une compartimentation vacuolaire efficace de Na+ [25] ou aussi à l'effet bénéfique de l'ABA [26].
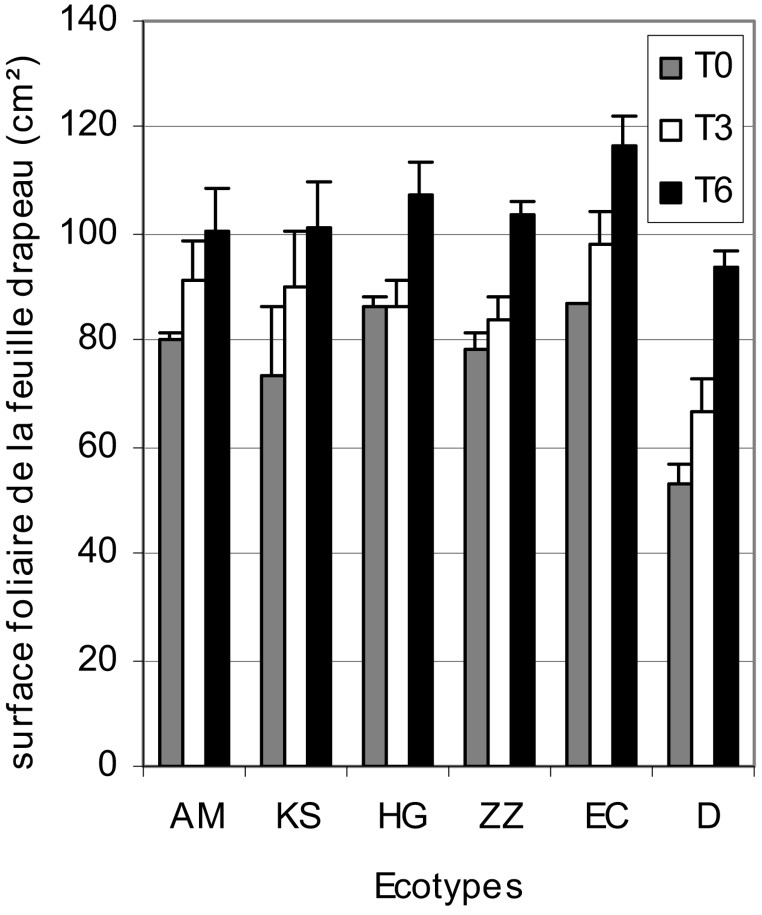
Variation de la surface foliaire paniculaire des écotypes de mil en fonction du stress salin.
6.2.2 Effet du stress salin sur les composantes de rendement
6.2.2.1 Effet sur la longueur de la chandelle principale (LOC).
La longueur de la chandelle n'est pas affectée par le sel (ppds variété = 2,53, ppds traitement = ns, ppds variété × traitement = ns) et semble être une caractéristique variétale. En effet, l'écotype EC possède les chandelles les plus longues, tandis que l'écotype D, ZZ et AM présente les plus courtes (Tableau 6).
Classification des écotypes de mil en groupes homogènes pour la longueur des chandelles (cm)
| Écotype | Moyenne |
| EC | 14,97 a |
| KS | 13,89 ab |
| HG | 13,78 ab |
| AM | 13,00 b |
| ZZ | 12,89 b |
| D | 12,43 b |
6.2.2.2 Effet sur le rendement en grains de la chandelle principale (RGC).
En l'absence de traitement salin, seul l'écotype KS se démarque des autres écotypes par ses chandelles les plus productives. L'écotype D occupe la dernière position, avec les chandelles les moins productives. L'application d'une eau modérément salée a augmenté le RGC de 10% et seulement de 2% en présence de contrainte saline sévère, mais cela n'est pas statistiquement significatif par rapport au témoin (Fig. 4). Cette contrainte n'a eu, en revanche, d'effet positif que sur l'écotype AM, dont le RGC se trouve amélioré de 25% par rapport au traitement modéré. Ce même traitement () a eu un effet dépressif sur KS et ZZ par rapport à , et non par rapport au témoin. En revanche, les chandelles de D, EC et HG n'ont pas subi de changement et donnent les mêmes rendements en grains, quelle que soit l'intensité du stress salin.
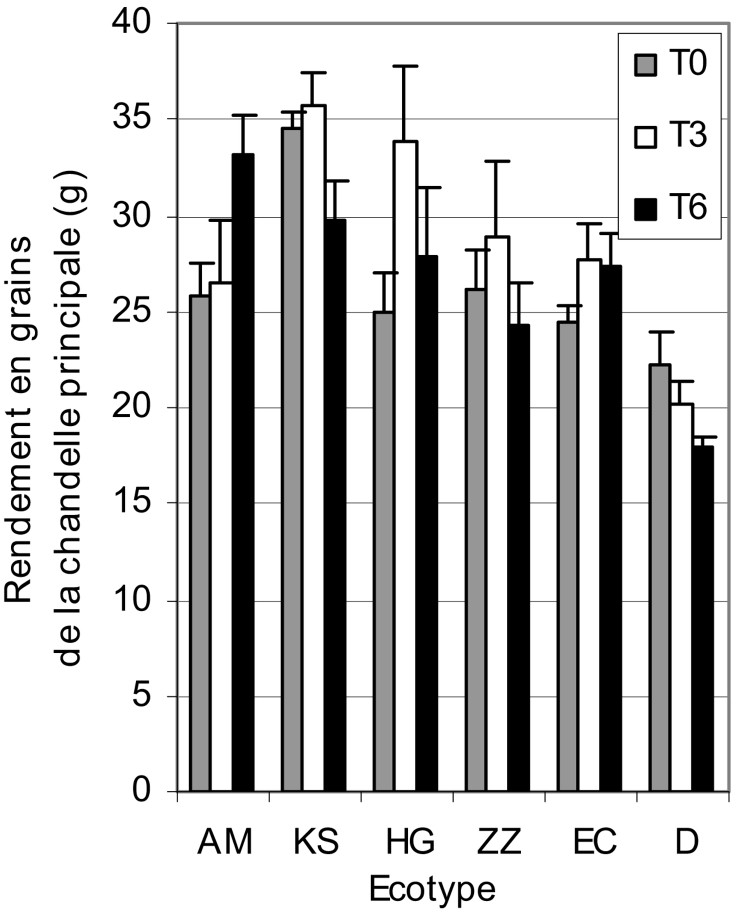
Variation du rendement en grains de la chandelle principale des écotypes de mil en fonction du stress salin.
Notons également que, tous traitements confondus, c'est l'écotype KS qui a été le plus productif, avec en moyenne un RGC de 33,5 g et D le moins productif, avec 20 g de grains par chandelle principale. Les écotypes HG, AM, EC et ZZ ne sont pas statistiquement différents et donnent en moyenne entre 26,5 et 29 g de grains par chandelle.
La présence d'interaction forte entre facteurs ne permet de conclure quant à l'effet de la salinité. En effet, les données montrent des résultats contrastés mais qui, de toute façon, suggèrent que les eaux salines (aux teneurs testées) n'ont pas d'effet nettement négatif. L'irrigation avec ces eaux saumâtres pourrait donc être recommandée pour stabiliser le rendement dans ces régions semi-arides, à la condition de démontrer dans nos prochains travaux que l'application systématique d'eau même légèrement saline ne détériore pas durablement les sols.
Dans ce contexte, certains auteurs [27] ont montré que l'usage des eaux titrant 4 dS/m pour l'irrigation n'entraîne que de faibles réductions de rendement. D'autres [28] ont utilisé des eaux titrant 12 dS/m pour l'irrigation du mil sur une période de 7 ans ; aucune baisse de rendement ni dégradation de sols n'a été constatée. De tels résultats ne peuvent être obtenus que s'ils sont accompagnés d'un système de drainage efficace.
6.2.3 Conclusion
D'après nos résultats relatifs aux essais en plein champ, nous avons vu que, si la salinité modérée a amélioré dans tous les cas la croissance et certaines composantes quantitatives (LOC et RGC) du rendement, la salinité sévère n'a pas engendré d'effets préjudiciables sur ces composantes. Ce même résultat a été signalé par Amazallag et al. [29] sur des populations de mil originaires du Sud tunisien.
L'amélioration du RGC est importante à considérer, car elle renseigne sur la tendance générale du rendement total. En effet, d'après certains auteurs [30], les corrélations qui lient RGC et le rendement total varient entre 0,78 et 0,89.
Une amélioration de rendement enregistrée par un apport d'eau modérément chargée en sel a été signalée sur le blé dur [31] et sur le triticale [23]. Elle trouve son explication dans l'amélioration de l'expansion foliaire. En effet, la feuille étant l'organe le plus actif de la plante, elle constitue le siège de toutes les activités biochimiques (notamment, les hydrates de carbone) ; de ce fait, toute amélioration de l'expansion foliaire semble à l'origine de l'augmentation du rendement [32]. D'ailleurs, quand la surface foliaire est réduite par la salinité, la production de carbohydrates devient insuffisante pour supporter la croissance et le rendement [33]. De nombreux auteurs [34–37] ont trouvé de fortes corrélations positives liant la largeur et la longueur de la feuille paniculaire aux PCP et RGC, ce qui prouve que toute amélioration des dimensions foliaires et, par conséquent, de la surface foliaire entraîne une amélioration du rendement.
L'absence d'effet préjudiciable d'une salinité élevée sur le rendement du mil serait due à la forte tolérance de cette espèce [38,39].
7 Conclusion générale
Notre travail a comporté deux volets.
Le premier a été réalisé au laboratoire et a permis de montrer que le recours à des eaux chargées en NaCl pour l'irrigation des graines de mil exerce une influence variable sur le taux de germination et la croissance des parties aussi bien aériennes que souterraines des différents écotypes. En effet, le taux de germination des semences stressées est supérieur à celui des graines non contraintes (quoique de façon non significative pour la dose ). De plus, la salinité n'a pas exercé d'effet dépressif sur l'élongation du coléoptile, mais a favorisé l'extension radiculaire. Ce test au laboratoire a également montré que les six écotypes de mil se classaient en deux groupes significativement différents. Le premier groupe est formé de AM, HG, D et EC, qui sont tolérants au sel, alors que les écotypes ZZ et KS sont résistants.
Le second volet a été réalisé au champ. Les résultats obtenus ont permis de noter que l'irrigation à l'eau modérément salée a favorisé, dans tous les cas, la croissance et le rendement du mil, alors qu'une salinité sévère n'a pas engendré d'effets préjudiciables vis-à-vis de ces composantes. Ce même résultat a été signalé sur des populations de mil originaires du Sud tunisien [29] ; il serait dû à la forte tolérance de cette espèce [38,39].
De plus, il a été remarqué des comportements différents des six écotypes de mil selon la qualité de l'eau d'irrigation utilisée, qui est due à la différence génétique des variétés dans l'accumulation des sels [40]. En effet, nous avons trouvé que les écotypes KS et ZZ (haute paille) sont les seuls à avoir subi une réduction de leur hauteur et une diminution du rendement en grains de leur chandelle principale face à la salinité sévère. La réduction de la croissance (hauteur) est un indice de la résistance au sel ; elle est d'autant plus importante que la variété est tolérante au sel [41]. En effet, la protection offerte par la croissance rapide (effet de dilution par la croissance du sodium accumulé dans les tissus) explique la relation entre vigueur de croissance et tolérance au sel signalée par plusieurs auteurs [42,43].
Les variétés à croissance foliaire rapide sont capables de retarder l'accès au seuil critique d'accumulation de Na+ et de maintenir un potentiel photosynthétique en produisant de nouvelles feuilles plus rapidement que le sel n'en élimine [44]. Ce mécanisme de dilution par la vigueur prend de l'ampleur chez les plantes en C4, réputées pour leur croissance rapide [45].
D'un autre côté, les écotypes HG, D, AM et EC présentent des similitudes face à la salinité. En effet, ils gardent la même stature et paraissent très stables sur le plan productif, puisqu'aucune composante de rendement n'a pas été pénalisée par l'action de la salinité. Des résultats similaires ont été trouvés sur le quinoa et la fétuque [46,47].
Finalement, nous avons retrouvé le même regroupement signalé dans le premier volet, qui montre que les écotypes AM, HG, D et EC, caractérisés par leur allongement radiculaire dans le même sens que la charge en sel, sont ceux qui ont été les moins lésés par le stress salin. En revanche, les écotypes KS et ZZ, dont la longueur radiculaire n'avait pas varié avec l'intensité du traitement salin, ont été les plus touchés par la salinité.
Ces constatations confirment les résultats trouvés au niveau des corrélations, qui montrent le rôle important que jouent les racines dans l'adaptation et la tolérance à la salinité chez le mil.
De même, ces résultats suggèrent que les tests de germination pourraient constituer des tests de sélection précoce des espèces végétales tolérantes au sel, comme cela a déjà été signalé par d'autres auteurs [48,49].