

1 Introduction
Le premier impératif d'une culture satisfaisante consiste en son implantation convenable et donc la réussite de la phase de germination est déterminante de celle de toute la période végétative. En effet, la germination est considérée comme un stade critique dans le cycle de développement des végétaux [1–3]. C'est elle qui conditionne l'installation de la plante autotrophe et probablement sa production ultérieure [4,5].
En Afrique, la culture du mil est pluviale. Les agriculteurs sont souvent obligés d'attendre les pluies qui n'arrivent pas au bon moment pour engager les semis ; d'où l'obligation de semer souvent en terre insuffisamment humidifiée ou même sèche, dans l'espoir de profiter des averses dès les premières gouttes. Les graines sont donc soumises à un déficit hydrique dès la germination.
La tolérance d'un bon nombre d'espèces au stress hydrique au moment de la germination constitue un avantage, car elle autorise, d'une part, l'établissement de ces espèces dans des zones que les taxons sensibles à la sécheresse ne peuvent coloniser et, d'autre part, l'occupation rapide de l'espace [6].
Le mil (Pennisetum glaucum (L.) R. Br.) est une céréale qui peut répondre à cette alternative grâce à un système radiculaire très développé, et aussi grâce à divers mécanismes adaptatifs efficients [7–9].
L'étude de l'influence des contraintes hydriques, simulées par l'utilisation d'agents osmotiques, est l'une des méthodes utilisées par les chercheurs pour étudier la résistance à la sécheresse des plantes au cours de la phase germinative. En effet, il a été démontré que la germination de l'arachide arrosée avec une solution de sucrose était en harmonie avec la résistance à la sécheresse de cette plante dans les conditions naturelles [10]. De plus, des relations étroites entre la résistance à la sécheresse du sorgho et sa capacité à germer à une pression osmotique de 10,88 bar ont été mises en évidence [11]. Dans une étude comparative sur l'effet du PEG sur la germination du mil et du sorgho, Saint-Clair [12] a démontré que la germination du mil est beaucoup moins affectée que celle du sorgho, puisque l'effet dépressif de la pression osmotique de 10,88 bar est comparable à celui que connaît le mil à une pression comprise entre 7 et 18,5 bar. D'ailleurs, dans la littérature, on a toujours signalé la supériorité du pouvoir germinatif du mil à des pressions osmotiques plus élevées que celles utilisées pour le sorgho, ainsi que sa remarquable résistance à la sécheresse [13–16].
De plus, la réponse des semences au stress hydrique pourrait également être un indicateur de la tolérance des plantes à la sécheresse pour les stades ultérieurs du développement, d'où l'intérêt de notre étude dont les objectifs principaux sont :
- – l'identification d'écotypes de mil tolérants à la sécheresse,
- – la recherche d'éventuelles corrélations pouvant lier le comportement au stade germination à celui de stades ultérieurs et qui pourraient constituer un test rapide à cet égard.
2 Matériel et méthodes
2.1 Matériel végétal
Cet essai a porté sur six écotypes autochtones de mil collectés à travers le territoire tunisien. Ces écotypes appartiennent à des étages bioclimatiques différents, sont cultivés sur des sols de textures diverses et sont irrigués avec des eaux de qualité médiocre. Ces écotypes sont :
- – D, écotype originaire de l'île de Djerba. Il est précoce, à tige courte et possède des chandelles petites, légères et peu productives ;
- – ZZ, écotype ramassé à Zarzis, présentant une longue tige et des chandelles peu larges à structure lâche. Son cycle est assez long, et il est productif ;
- – HG, écotype collecté au cap Bon (Hammam Laghzaz), possédant une courte tige et des chandelles larges et compactes. C'est un mil à cycle court et moyennement productif ;
- – KS, mil de la plaine de Kairouan, très vigoureux, à haute paille, chandelles de fort calibre très productives. C'est un écotype à long cycle ;
- – AM, écotype trouvé dans la zone de El Amra (Sfax). Il est du type mixte (précoce à semi précoce), avec des chandelles larges et assez productives ;
- – EC, mil collecté à Echaba (Mahdia). C'est un écotype moyen qui a, à la fois, les caractéristiques des écotypes à cycle long (tige longue) et celles des écotypes à cycle court (chandelles peu productives et peu vigoureuses). Il est aussi semi précoce et moyennement productif.
2.2 Protocole expérimental
Les essais sont conduits au laboratoire et au champ.
Au laboratoire, il a été procédé à des essais de l'effet du polyéthylène glycol (PEG 4000) sur le taux de germination et sur les longueurs de l'épicotyle et des racines des différents écotypes étudiés. Pour chaque écotype, les grains sont lavés à l'eau du robinet, désinfectés pendant quelques minutes dans de l'eau de Javel diluée à 10%, puis rincés à l'eau distillée. Ils sont placés à raison de 50 grains dans des boites de Pétri sur du papier filtre imbibé avec, soit 20 ml d'eau du robinet (témoin), soit avec des solutions de PEG 4000. Pour créer la contrainte hydrique, nous avons préparé des solutions développant des potentiels osmotiques de 1 MPa (stress modéré) et de 2 MPa (stress sévère) à partir de la régression polynomiale liant le potentiel osmotique (π en MPa) à la concentration en PEG (C en mol/l) [17].
Après une semaine d'incubation à l'obscurité et à 30 °C, nous avons compté les graines germées et mesuré les longueurs respectives de leur épicotyle et de leur radicule.
Les essais en plein champ ont été conduits dans une zone semi-aride. Les semis ont été réalisés le 23 mai 2005. L'eau d'irrigation utilisée est celle courante, titrant 1 g/l de NaCl.
Pour réaliser le stress hydrique, nous avons adopté un dispositif permettant d'apporter la dose d'eau souhaitée.
Les besoins théoriques en eau du mil sont estimés, d'après [18,19], à 580 mm ().
Les doses choisies sont :
- – T100, témoin irrigué à 100% ETM ;
- – T70, traitement recevant 70% ETM ;
- – T35, traitement irrigué à 35% ETM ;
Les différents écotypes ont été semés en cinq répétitions. Chaque répétition est représentée par un carré de 25 m2. La densité de semis est de 200 000 plants/ha.
Les traitements sont espacés de 3 m et sont entourés, dans toutes les directions, de lignes de bordure.
2.3 Mesure de la croissance des plantes
La hauteur des plantes (HAT), en centimètres, est déterminée à l'aide d'une règle graduée du collet de la plante à l'insertion de la chandelle, au stade maturité.
La surface foliaire de la feuille drapeau ou feuille paniculaire (SFD) en centimètres carrés, est mesurée à l'aide d'un planimètre CI-202, au stade pâteux.
2.4 Mesures des composantes de rendement
À la récolte, les chandelles principales sont coupées, mesurées, pesées, puis battues.
Les caractéristiques propres à la chandelle principale sont :
- – longueur de la chandelle (LOC) en cm ;
- – poids de la chandelle (PCP) en grammes ;
- – rendement en grains de la chandelle (RGC) en grammes.
2.5 Analyse statistique
Les résultats sont soumis à l'analyse de la variance à un ou à 2 facteurs, et les moyennes sont comparées selon la méthode de Bonferonni, basée sur la plus petite différence significative.
Chaque moyenne est affectée d'une lettre, les moyennes suivies d'une même lettre ne sont pas significativement différentes.
3 Résultats et discussion
3.1 Résultats des essais de germination (laboratoire)
3.1.1 Taux de germination
Les différentes concentrations de PEG ont eu un effet significatif sur la germination des semences de mil. Le taux de germination varie entre 100% pour le témoin et 73% pour le milieu le plus concentré en PEG. Toutefois, une légère carence hydrique () n'a pas eu de conséquences notables sur le taux germinatif et a même amélioré le taux de germination pour certains écotypes (Fig. 1). En moyenne, les taux de germination les plus élevés sont enregistrés chez les populations D et ZZ (92%) ; il est similaire chez les écotypes KS et HG (87%) et il est le plus bas pour les écotypes EC et AM (environ 83%).
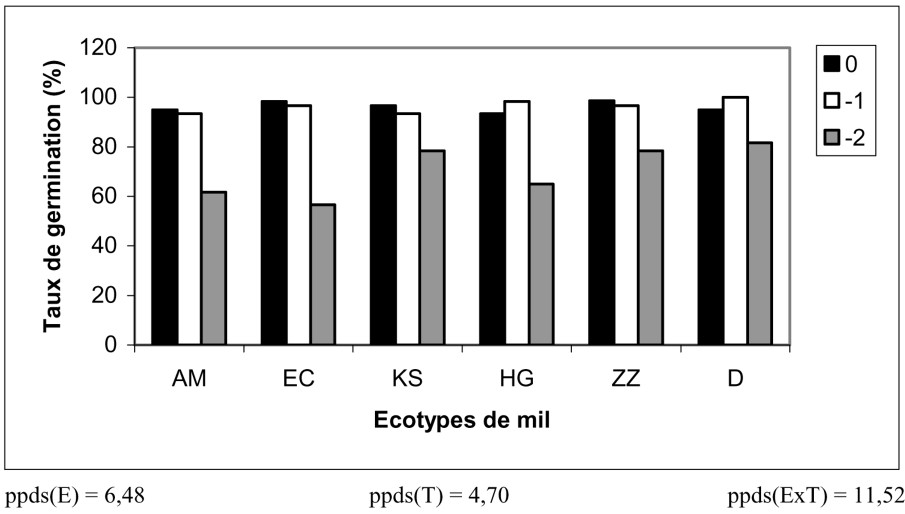
Influence du stress hydrique sur le taux de germination des six écotypes de mil.
L'exposition des graines de mil à une carence hydrique sévère a diminué le taux de germination. En effet, à , le taux de germination du mil est de 73%, ce qui est proche de la valeur donnée par [20], à savoir 75% pour un potentiel de . Cette valeur constitue, d'après cet auteur, une limite pour laquelle la quasi-totalité des espèces ne germent plus et qui confirme le caractère adaptatif de cette espèce à la sécheresse, tout au moins au cours de la phase de germination. Cette inhibition de la germination résulterait en particulier d'une difficulté d'hydratation des tissus, qui se répercute sur le processus d'élongation de la radicule [21], et d'une difficulté de la pénétration de cette molécule dans les graines, ce qui ne favorise pas l'ajustement osmotique [22]. Cependant, il est à remarquer qu'un léger déficit hydrique améliore cette faculté germinative. Un tel résultat a été mentionné par de nombreux auteurs et sur diverses cultures [23–26].
Par ailleurs, des différences génotypiques de sensibilité au stress hydrique, au stade de la germination, ont été signalées [27]. En effet, les écotypes issus du sud (D et ZZ) ont présenté des taux de germination supérieurs à ceux des autres écotypes, ce qui montre l'importance de l'origine de la semence quant à la résistance à la sécheresse. Il a été trouvé que les cultivars d'origine indienne supportent plus la sécheresse que ceux originaires des conditions humides de la Géorgie (États-Unis) [20].
3.1.2 Longueur de l'épicotyle
Plus la contrainte hydrique est sévère, plus le coléoptile est court. En effet, un potentiel hydrique de diminue la longueur du coléoptile de 84,5%, alors qu'un stress modéré de réduit cette dimension de 44% seulement (Tableau 1). Ce résultat est corroboré par [28] chez le blé. De plus, il a été constaté que c'est la population D qui a le plus court coléoptile () et que les écotypes ZZ et KS possèdent les coléoptiles les plus longs.
Effet du stress hydrique sur l'élongation du coléoptile, exprimée en % du témoin
| Potentiel osmotique (bar) | Taux de réduction de l'élongation du coléoptile en% du témoin |
| 0 | 100,0a |
| −10 | 44,0b |
| −20 | 84,5c |
a Les valeurs caractérisées par la même lettre ne sont pas significativement différentes entre elles au seuil de signification (5%).
b Les valeurs caractérisées par la même lettre ne sont pas significativement différentes entre elles au seuil de signification (5%).
c Les valeurs caractérisées par la même lettre ne sont pas significativement différentes entre elles au seuil de signification (5%).
3.1.3 Longueur de la radicule
Une légère carence hydrique améliore la longueur de la radicule chez tous les écotypes (15,8%). En revanche, un stress hydrique sévère la réduit de plus de 88% (Fig. 2). Un tel résultat a été mentionné [29]. Cette réduction serait due à un arrêt de la division et de l'élongation cellulaire au niveau de la racine, conduisant à une sorte de « tubérisation » [30]. La tubérisation et la lignification du système racinaire permettent à la plante une « entrée en vie » ralentie, en attendant que les conditions redeviennent favorables [31]. Par ailleurs, aucune différence significative entre les divers écotypes n'a été enregistrée.
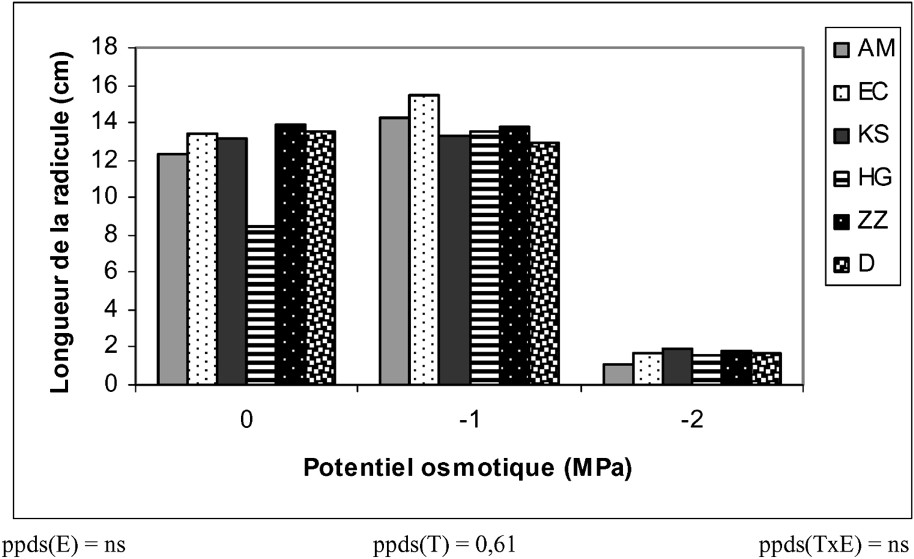
Variation de la longueur de la radicule des différents écotypes de mil en fonction du stress hydrique.
3.1.4 Conclusion
L'exposition des graines de mil à une carence hydrique () sévère a diminué le taux de germination des graines de mil d'environ 25% seulement, ce qui constitue, à notre connaissance, une limite pour laquelle la quasi-totalité des espèces ne germent plus et qui confirme le caractère adaptatif de cette espèce à la sécheresse, tout au moins au cours de la phase de germination. Cette espèce, qui a pu germer même en présence d'un potentiel de suscite plusieurs questions sur son importance en tant que semence « fourragère », capable de s'installer même en conditions sèches. Ce caractère d'adaptation est très souhaité, puisqu'il détermine le nombre de plantes par unité de surface et permet d'étendre l'aire de culture de cette plante vers des régions plus sèches. De plus, nous avons remarqué qu'un léger déficit hydrique améliore la faculté germinative de cette espèce et stimule l'élongation radiculaire. Au contraire, lorsque la contrainte hydrique est sévère, la radicule paraît plus sensible que le coléoptile. Par ailleurs, des différences génotypiques de sensibilité au stress hydrique, au stade de la germination, ont été signalées. En effet, les écotypes issus du sud (D et ZZ) ont présenté des taux de germination supérieurs à ceux des autres écotypes, ce qui montre l'importance de l'origine de la semence quant à sa résistance à la sécheresse.
3.2 Résultats des essais de rendement (champ)
3.2.1 Effet du stress hydrique sur la croissance
Croissance en hauteur. L'analyse de la variance à un facteur a montré que les écotypes de mil sont divisés en deux grands groupes, selon la hauteur de la tige principale. Le premier groupe, constitué par les populations ZZ et KS, mesure plus de 2 m. Le deuxième groupe représente le reste des écotypes mesurant moins de 2 m. À l'intérieur de ce groupe, la population D occupe la dernière place, précédée par HG. De plus, le traitement modéré n'a pas eu d'effet sur la hauteur des plantes (Fig. 3), alors que le traitement sévère a raccourci les tiges principales d'environ 22% ; ce sont ZZ et KS qui ont été les plus touchés (raccourcissement de plus de 40% pour ZZ). Ce même résultat a été signalé sur le mil [8,32]. De plus, ils ont remarqué que les réductions les plus importantes sont celles qui touchent les variétés à tige très longue.
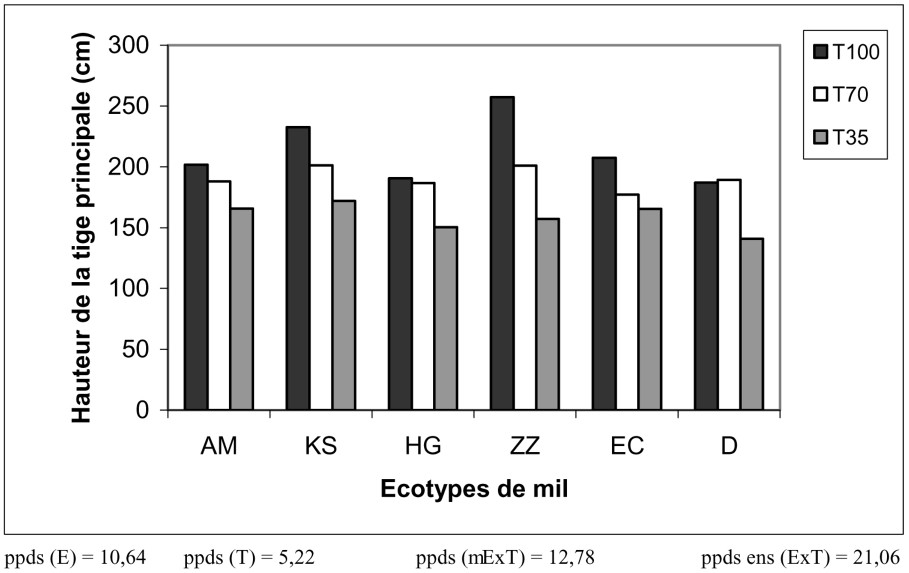
Évolution de la hauteur de la tige principale des écotypes de mil en fonction du traitement hydrique.
Croissance de la surface foliaire. Les plantes contraintes présentent des feuilles drapeaux d'autant plus petites que le stress hydrique est plus intense (Fig. 4). Les réductions de la surface foliaire paniculaire sont respectivement de 11% pour T70 et de 34,7% pour T35. Les résultats consignés sur la Fig. 4 montrent également que les surfaces foliaires de tous les écotypes n'ont pas été touchées par le manque d'eau modéré. En présence de la contrainte hydrique sévère, ce sont les écotypes HG et D qui ont présenté les réductions foliaires les plus importantes (38% de réduction par rapport à leurs témoins respectifs bien irrigués). En revanche, KS et ZZ occupent une position intermédiaire avec une réduction foliaire d'environ 32%. Une réduction de la surface foliaire du mil a été signalée par plusieurs travaux [33–39]. Elle serait due à un ralentissement de la division et de l'expansion cellulaire [40].
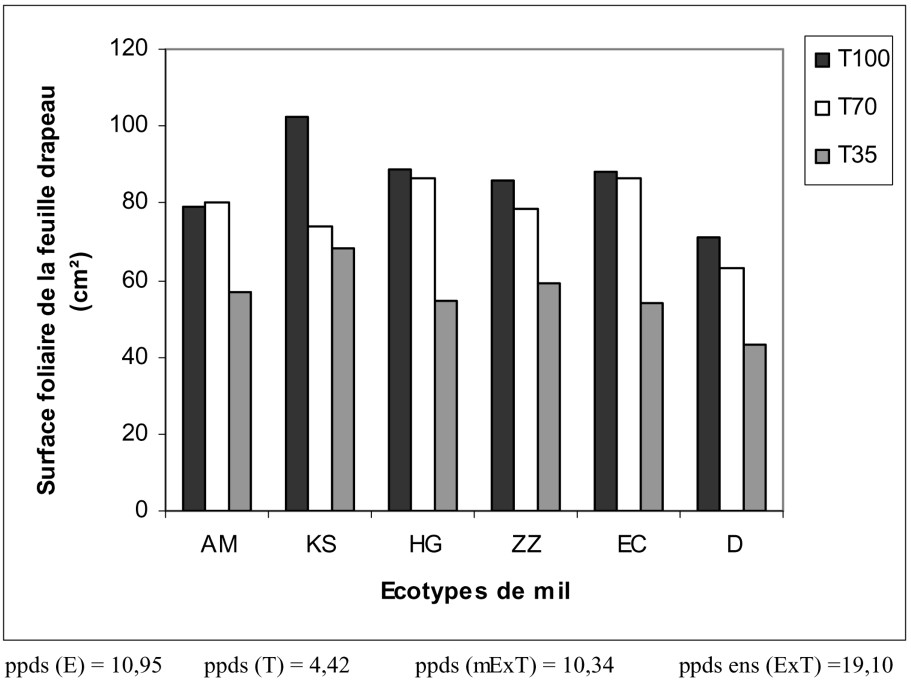
Variation de la surface foliaire paniculaire des écotypes de mil en fonction du traitement hydrique appliqué.
3.2.2 Effet du stress hydrique sur les composantes de rendement
Effet sur la longueur de la chandelle principale (LOC). La variable LOC présente un effet génotypique significatif et permet de classer les écotypes en trois groupes (Fig. 5) :
- – les chandelles principales 1 de ZZ et KS mesurent plus de 14 cm ;
- – EC et AM donnent des chandelles d'une longueur moyenne de 13 cm ;
- – HG et D présentent des chandelles d'environ 12 cm de longueur.
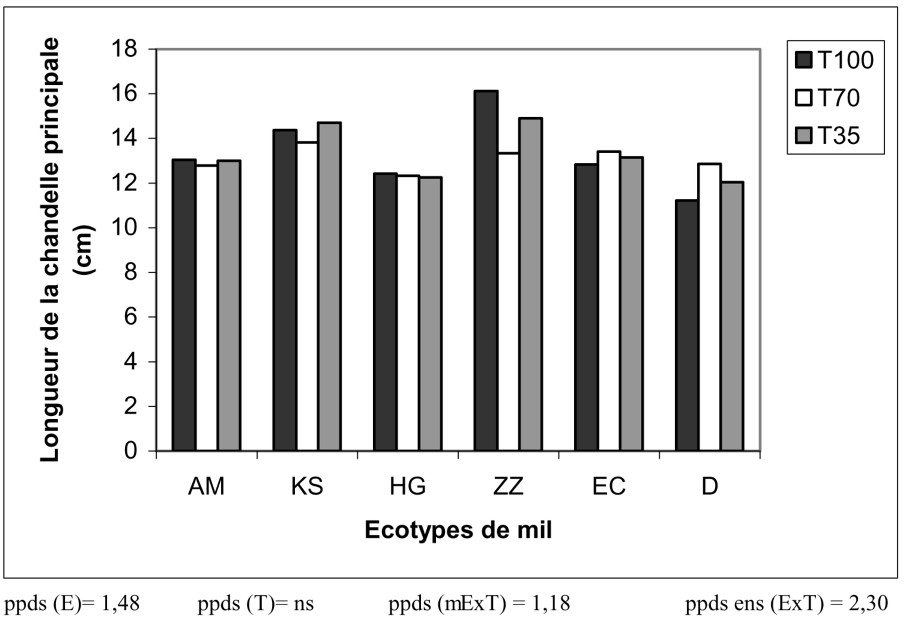
Variation de la longueur de la chandelle principale en fonction du traitement hydrique appliqué et des écotypes de mil.
La contrainte hydrique n'a pas eu de conséquence notable sur cette composante de rendement.
3.2.3 Effet sur le poids de la chandelle principale (PCP)
Génétiquement et selon le poids de leur chandelle principale, les écotypes sont classés en quatre catégories. Par ordre décroissant, KS occupe de loin le premier rang, il est suivi par ZZ et EC ; puis viennent en troisième place AM et HG. L'écotype D occupe le dernier rang, avec une chandelle pesant la moitié de celle de KS.
Les résultats présentés sur la Fig. 6 montrent également que seule la dose très restrictive a agi défavorablement sur le poids des chandelles principales. La perte en poids est dans ce cas de 25%. De plus, les écotypes de mil les plus touchés par la sécheresse sont KS et ZZ (environ 35% de pertes).
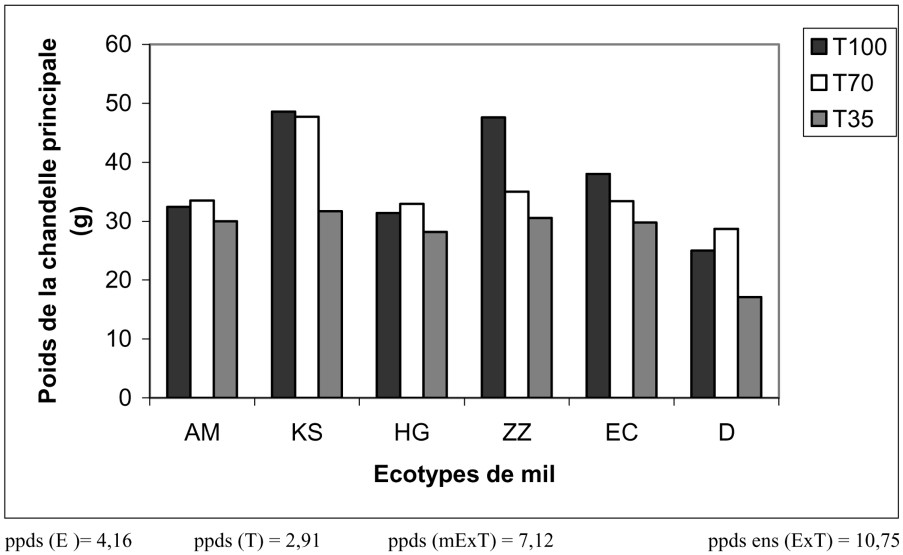
Variation du poids de la chandelle principale en fonction du traitement hydrique appliqué et des écotypes de mil.
3.2.4 Effet sur le rendement en grains de la chandelle principale (RGC)
Selon le rendement en grains de leur chandelle principale, les écotypes sont classés comme suit : KS avec en moyenne 30,5 g, ZZ avec 27 g, puis EC, AM et HG avec des rendements variant entre 23,5 et 25 g, et finalement D, avec seulement 18,5 g.
Comme pour le poids de la chandelle principale, le rendement en grains de ces mêmes chandelles est diminué seulement en présence du traitement sévère (la diminution est de 25%) ; ce sont les écotypes KS et ZZ qui ont été les plus touchés (Fig. 7).
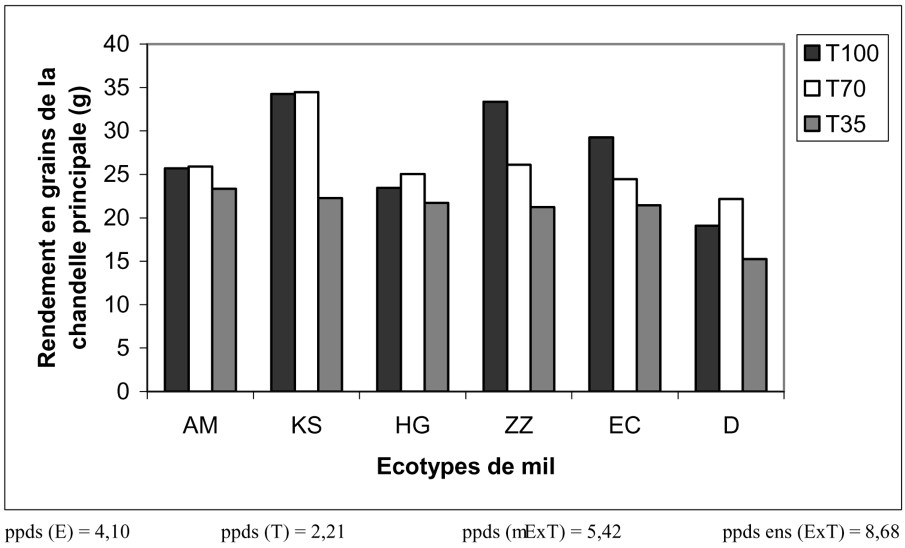
Variation du rendement en grains de la chandelle principale en fonction du traitement hydrique appliqué et des écotypes de mil.
3.2.5 Corrélations entre stade de germination et stade adulte
Nous avons cherché à mettre en évidence si certains paramètres mesurés au stade adulte seraient en relation avec le stade de germination. En effet, certains auteurs [41–43] ont trouvé que la longueur du coléoptile et de la radicule est liée à la taille finale du plant de mil. Ainsi, d'après ces auteurs, la haute stature est liée à un long coléoptile, alors qu'un court coléoptile est caractéristique d'une population naine ou à courte paille. De plus, les écotypes à long coléoptile donneraient ultérieurement des chandelles longues et productives.
Le Tableau 2 résumant les corrélations trouvées montre que la hauteur de la tige est corrélée positivement à la longueur du coléoptile et, à un degré moindre, à celle de la radicule en présence de stress hydrique ; autrement dit, plus un coléoptile et une radicule sont développés sous contrainte hydrique, plus la stature est importante.
Lors de l'étude de la germination, nous avons trouvé que ce sont les écotypes KS et ZZ qui présentaient les coléoptiles les plus longs. À maturité, ce sont ces deux écotypes qui ont présenté les hauteurs les plus importantes ().
Ce genre de corrélation est important à établir, car il peut constituer un test précoce de sélection pour le choix d'écotypes de mil à développer dans les programmes d'amélioration. D'ailleurs, de nombreux programmes de recherche basés sur les critères de sélection précoce au stress hydrique sont basés sur les caractéristiques de la germination, essentiellement la longueur du coléoptile [44].
3.2.6 Conclusion
Nos résultats relatifs aux essais en plein champ ont montré que l'application d'une contrainte hydrique modérée n'a pas affecté les caractéristiques propres à la chandelle principale. Ce résultat est corroboré par Yazar et al. [45] et Crépin et al. [46]. Au contraire, un effet bénéfique de ce traitement a été parfois observé chez le mil comme chez d'autres cultures [47,48]. En revanche, le traitement hydrique sévère a pénalisé les composantes de rendement. En effet, les chandelles sont devenues effilées, moins compactes et moins garnies, plus légères, et leur rendement en grains a diminué.
Les chandelles principales semblent être les seules qui contribuent significativement au rendement final [36]. Elles sont si importantes qu'elles fournissent à elles seules souvent autant, et quelquefois plus de grains que l'ensemble du tallage secondaire [49] ; donc toute modification les concernant se répercute sur le rendement final.
4 Conclusion générale
Notre travail a comporté deux volets : le premier a été réalisé au laboratoire et a permis de montrer qu'un léger déficit hydrique avait amélioré la faculté germinative de cette espèce et stimulé l'élongation radiculaire. Au contraire, lorsque la contrainte hydrique est sévère, la radicule paraît plus sensible que le coléoptile. Ce test au laboratoire a également montré la supériorité des écotypes originaires du Sud, ce qui montre l'importance de l'origine de la semence quant à la résistance à la sécheresse.
Le second volet a été réalisé au champ. Les résultats obtenus ont permis de noter que la croissance du mil ainsi que ses composantes de rendement ont été réduites lorsque l'irrigation était très déficitaire. En revanche, une sécheresse modérée n'a eu de conséquence notable, ni sur le développement, ni sur les composantes productives.
De plus, il a été remarqué des comportements différents des six écotypes de mil selon l'intensité du stress hydrique. Ainsi, la sécheresse sévère a provoqué une réduction de la hauteur des plantes pour les écotypes les plus vigoureux (KS et ZZ) et un rétrécissement des feuilles drapeaux pour les écotypes de vigueur moyenne à faible (EC, HG et D). De même, ce sont les écotypes les plus productifs (KS et ZZ) qui ont été les plus pénalisés par la contrainte hydrique.
Nous avons également pu établir des corrélations importantes, qui montrent que la longueur du coléoptile et de la radicule sont liées à la taille finale du plant de mil. Tout ceci montre que l'étude de la germination est un outil scientifique nécessaire, qui peut avoir des répercussions agronomiques pratiques et exploitables. En effet, certains paramètres mesurés au stade adulte seraient en relation avec le stade germination. C'est selon le mode d'irrigation déployé que le choix des semences à employer doit être décidé. Ainsi, et en cas d'irrigation déficitaire, le recours à des écotypes de mil à coléoptile vigoureux est-il à conseiller. Dans la pratique et indépendamment de la germination, c'est la levée de la jeune plante de mil qui intéresse le plus l'agriculteur. Plus celle-ci est vigoureuse, plus elle supporte les risques du déficit hydrique à ce stade, et mieux elle s'acclimate, ce qui lui permet de mieux résister aux stress hydriques survenant après l'épiaison–floraison.