

1 Introduction
In most of the studied songbird species, males learn the complex syllables syntax of their song through a process of vocal imitation that presents strong similarities with the mechanisms underlying the ontogeny of human language [1,2]. This striking parallel has made songbirds the most relevant biological system to understand language acquisition and its neural correlates [3,4]. Consequently, song perception processes have been extensively explored in male songbirds for many years [5–7]. Conversely, little is known about the perception of calls, which represent the category of acoustic signals that is shared by both males and females. Calls are usually signals composed by a single note and have various roles [8]. Although crucial for understanding acoustic communication in birds, few studies to date dealt with call perception. To get a comprehensive view of acoustic communication in the songbird model, there is thus a strong need of studies focusing on calls.
The sound-induced expression of immediate early genes (IEG) like ZENK and FOS allowed us to identify specialized brain structures that underlie the capacity of songbirds to recognize conspecific vocalizations (reviewed in [9]). The NCM (caudal medial nidopallium [10,11]) and the CMM (caudal medial mesopallium), two regions analogous to secondary auditory cortices in mammals [12], appear to be possible areas for the extraction of biologically relevant information in acoustic signals [9,13–16]. One hypothesis is that the magnitude of the IEG response in these regions does not depend only on sound acoustic structure [17], but also on the relevance of the signal, which can be defined as the stimulus type [18–20] or by the individual experience of the bird [21–23]. Contrary to songs, calls have never been used to investigate whether the neural responsiveness correlates with the sound subjective salience.
Bird vocalizations convey several pieces of information such as sexual and/or territorial motivations, as well as gender, species identity, or individual identity [5]. Individual identity is of particular interest to investigate whether brain responsiveness depends on sound acoustic structure or sound perceptive salience. Indeed, an acoustic-based individual recognition between mates has been experimentally demonstrated in several monogamous bird species [24–31]. In zebra finches (Taeniopygia guttata), which represent one of the favourite model systems of songbird neuroethology, male and female form life-long pair bonds and breed in loose colonies [32]. Among the number of different vocalizations produced by this bird, male and female use distance calls to remain in contact especially when the birds are visually isolated from their mate during flock foraging [33]. Distance calls are known to encode sex as well as species identity and individual identity [34–36]. Because female zebra finches perform a robust recognition of their mate calls among other conspecific male calls [37], this species is a good candidate to investigate whether brain activity in response to calls depends on the acoustic structure of the stimulus (call structure) or its subjective salience (call individual identity).
The aim of the present paper is to compare the brain ZENK expression elicited by mate versus non-mate calls in female zebra finches. We examined neural responsiveness in the caudal telencephalon by measuring ZENK activation following call playback.
2 Material and methods
2.1 Subjects
Six adult female zebra finches (Taeniopygia guttata) served as the subjects for this experiment and were naive to all testing procedures. These birds were bred in our aviary (12L/12D photoperiod with adapted wavelengths; food and water ad libitum; temperature between 23 and 25 °C). Females were paired with their male mates for several months, in separated pair-cages allowing visual and acoustic contacts with all the other birds of the aviary. Each pair raised at least one brood before the tests. All experiments occurred between 8 and 10 a.m. During isolation and stimulation periods, conditions of temperature, food and water were the same as in the aviary. The experimental protocols were approved by the Jean Monnet University's animal care committee.
2.2 Acoustic stimuli
In order to test whether females' brain responsiveness is correlated with the subjective salience of sounds, female subjects were divided in two experimental groups:
- – females exposed to their mate calls (mate stimulus),
- – females exposed to the calls of non-mate males (non-mate stimulus). In order to avoid pseudoreplication, we used a different non-mate male for each female subject in this group.
All stimuli (two or three calls per male) were collected using a Sennheiser MD42 microphone placed 0.3 m above the cage and connected to a Marantz PMD690/W1B recorder with 22,050-Hz sampling rate.
2.3 Analysis of acoustic stimuli
In order to test the potential influence of sounds acoustical characteristics on brain responsiveness, we analysed the acoustic structure of the male calls used as stimuli for exposure. Using Syntana software [38] and Praat version 4.0.19, we analyzed the whole set of calls of the six males (three mates and three non-mates), which corresponds to a total of 16 male distance calls (three per male, except two males with only two calls recorded).
The zebra finch distance call is a complex sound with a fundamental frequency associated with several harmonics (Fig. 1A). This sound is frequency and amplitude modulated. The male distance call has an elevated fundamental frequency (600–1000 Hz) compared to that of the female one (400–500 Hz). It is typically a frequency-modulated downsweep of around 100 ms [34,35,39]. In our population of zebra finches, the typical male distance call can be divided into two segments of different durations (Fig. 1A): a first segment composed by an initial rapid ascending frequency modulation of low amplitude followed by a short stable part (the tonal component defined by [35]), and a second segment defined by a long and loud descending frequency modulation (the noise component or downsweep component defined by [35]). We defined 20 spectral, temporal and intensity cues to describe the calls' acoustic structure.
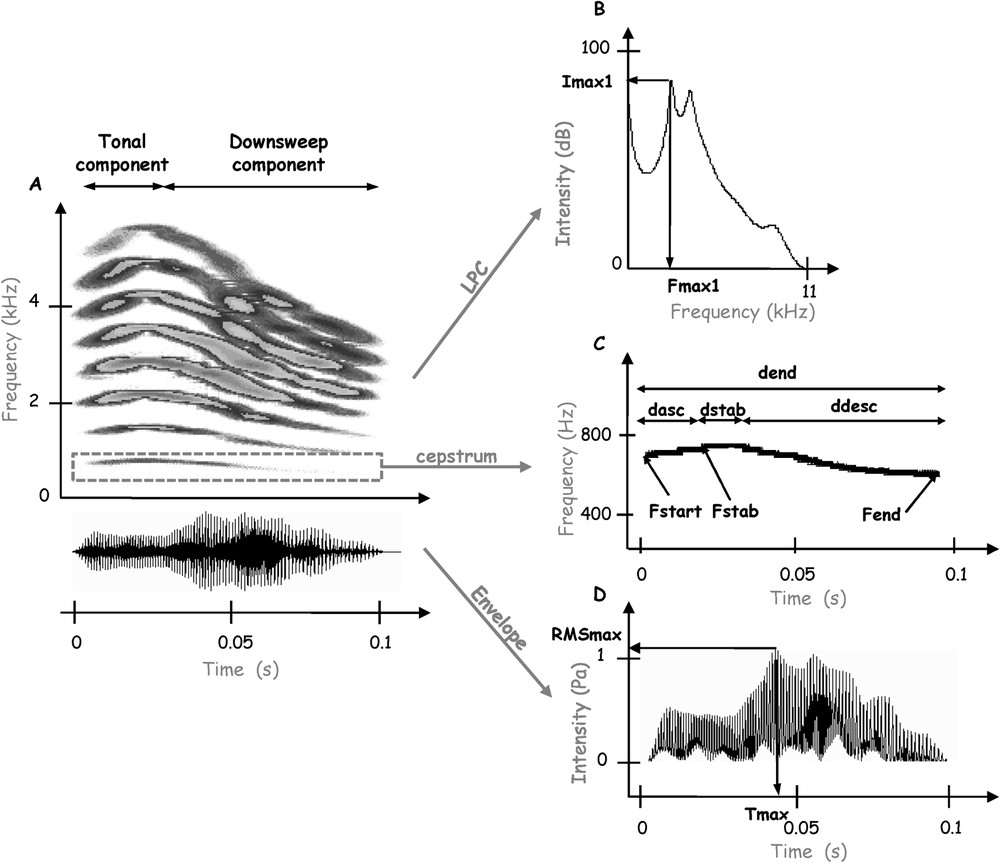
Analysis of a male distance call. (A) Spectrogram and oscillogram. The distance call can be divided into a tonal component followed by a downsweep component. (B) Average power spectrum calculated with the LPC method. The frequency of the first peak amplitude (, Hz), the intensity of this peak (, dB), as well as the frequency of the second peak amplitude (, Hz) and the intensity of this peak (, dB), are measured. (C) Fundamental frequency of the call obtained with the cepstrum method and used to describe the frequency modulation of the call: we measure the duration of the ascending frequency modulation of the tonal component (, s), the duration of the stable part of the tonal component (, s), the duration of the descending frequency modulation of the noise component (, s), the total duration of the call (, s), as well as the start frequency (, Hz), the frequency of the stable part (, Hz), and the end frequency of the call (, Hz). (D) Amplitude envelope of the call. The parameters measured are the mean intensity of the entire call (RMSaver, dB), the loudest intensity in the call (RMSmax, Pa) and the duration between the beginning of the call and the time at which the highest amplitude in the call occurs (, s).
To describe the frequency modulation of the call, we first isolated the fundamental frequency using the cepstrum method [38]. Four temporal parameters were measured from the fundamental frequency (Fig. 1C): the duration of the ascending frequency modulation of the tonal component (, s), the duration of the stable part of the tonal component (, s), the duration of the descending frequency modulation of the downsweep component (, s), and the total duration of the call (, s). Three spectral parameters were also measured from the fundamental frequency (Fig. 1C): the start frequency (, Hz), the frequency of the stable part (, Hz), and the end frequency of the call (, Hz). These parameters were used to calculate the two following parameters: , the slope of the ascending frequency modulation (Hz s−1) calculated as ( – )/, and , the slope of the descending frequency modulation (Hz s−1) calculated as ( – )/().
To describe the amplitude change over time, we measured three parameters from the envelope of the signal (Fig. 1D): the mean intensity of the entire call, represented by the root-mean-square signal level (RMSaver, dB), the loudest intensity in the call (RMSmax, Pa) and the duration between the beginning of the call and the time at which the highest amplitude in the call occurs (, s). We then calculated the parameter RMSmax/RMSaver (Pa dB−1).
In order to describe the energy spectrum of the call, the centre of gravity (CG, Hz) was assessed on the FFT spectrum of the signal. Six parameters were measured from the average power spectrum calculated from the total length of the call with the LPC method (applying linear predictive coding) (Fig. 1B): the frequency of the first peak amplitude (, Hz), the intensity of this peak (, dB), which is normalised by calculating the ratio /RMSaver, the frequency of the second peak amplitude (, Hz), the intensity of this peak (, dB), which is normalised by calculating the ratio /RMSaver.
We then assessed the effect of male identity on call parameters. As our sample failed normality and did not respect homoscedasticity, we performed a non-parametric analysis of variance (Kruskall–Wallis ANOVA, ). Using the parameters that vary significantly among male individuals, we carried out a principal component analysis (PCA) to derive ten composite scores corresponding to the ten principal components (PC1 to PC10). These ten scores derived from the PCA were normally distributed and allowed us to perform a discriminant analysis (DA) in order to investigate whether the male calls of our data set share common acoustic features. The discriminant functions derived from the DA were used in a hierarchical ascending classification (using the method of Ward with squared Euclidean distances). This classification allowed defining groups of calls reflecting their common acoustic cues and defining call types. All statistical tests were performed using Statistica Software version 6.
2.4 ZENK immunocytochemistry
2.4.1 Acoustic stimulation procedure
Each bird () was acoustically isolated in a soundproof chamber the night before the experiment. The birds were housed on a 12L/12D photoperiod, except in the last six hours and during the stimulation period where they were kept in darkness in order to avoid any uncontrolled stimulation of brain auditory areas by spontaneous singing behaviour [40]. All acoustic stimuli were broadcast through a high-fidelity speaker (Triangle Comete 202, sound level: 60 dB at 1 m) connected to a Marantz PMD690/W1B recorder and an amplifier (Yamaha AX-396).
The playback procedure was as follows: one acoustic stimulus consisted of 60 repetitions of a series of 20 calls (2 calls per second, i.e. natural call rate of a male, using two or three different calls of the male individual at random). Two consecutive series were separated by 20 s of silence. Consequently, the total duration of the stimulus was 30 min. The acoustic stimulation was followed immediately by 30 min of silence, during which the animal was kept inside the isolation chamber.
2.4.2 Tissue preparation
The bird was killed by decapitation one hour after the start of the stimulation period, which is within the time of peak ZENK protein expression [40]. The brain was then rapidly frozen on liquid nitrogen. Brain sections (14 μm) were performed in the sagittal plane using a cryostat in the left hemisphere. The sections were mounted onto slides (Superfrost plus, Mewzel Glaser GmbH), air dried overnight and stored at −20 °C.
2.4.3 Immunocytochemistry (ICC)
Each one of the following steps was followed by three washes (10 min each) in 0.01 M phosphate buffer saline (PBS 0.01 M Sigma Laboratories # P-3813). First, slides were fixed in 4% paraformaldehyde for 10 min at room temperature (RT). Next, slides were incubated as follows: (1) 30 min at RT in blocking solution (BS) (0.5% albumin and 0.3% Triton X-100 in PBS 0.01M); (2) biotin blocking treatment (Dako # X0590) at RT; (3) one night at 4 °C in a commercially available Egr-1 antibody (Santa Cruz Biotechnology, catalogue # sc-189) diluted at 1:1000 in BS; (4) 2 h at RT in biotinylated goat anti-rabbit IgG (Sigma Laboratories # B7389) diluted at 1:200 in BS; (5) 1 h 30 min at RT in avidin-biotin peroxidase complex (Sigma Laboratories # E-2886) diluted at 1:150 in PBS 0.01M, followed by a incubation in -diaminobenzidine (DAB) (Sigma Laboratories PAST DAB Tablet sets). Reaction time in the DAB was held constant at 15 min across all different ICC runs. Controls were run by omitting the primary Egr-1 antibody used in step 3. Finally, the sections were dehydrated and coverslipped with Entellan.
2.4.4 Quantification of ZENK expression and statistical analyses
We quantified the level of ZENK expression in each subject using 4 to 9 sections located between 0 and 700 μm sagittal to the midline. Digital images of the sections (NIKON Coolpix 4500 on top of a LEICA DMLB microscope, 10× objective, 10× eyepiece) were captured in grey levels. We quantified ZENK-ICC labelling into a sampling window whose boundaries were defined as a (except for hippocampus: ). We placed this window in four anatomical areas (Fig. 2): the dorsal NCM (NCMd), the ventral NCM (NCMv), the CMM, and the hippocampus (HIP).

(A) Zebra finch brain parasagittal section stained with cresyl violet (right hemisphere). (B) Positions of the sampling areas. The NCM is observed in front of the cerebellum (cb) and is separated from the CMM by a region of weaker ZENK expression, corresponding to a sub-region of Field L, Field L2. The hippocampus (HIP) is separated from these areas by the ventricle. We quantified ZENK labelling into a sampling window of 500×500 μm (except for hippocampus: 250×500 μm) placed in the dorsal NCM (NCMd), the ventral NCM (NCMv), the CMM and the hippocampus (HIP). (C) ZENK expression in the HIP sampling window in response to mate stimulus and non-mate stimulus. The white arrows indicate examples of ZENK-immunoreactive cells.
We counted the number of ZENK immunoreactive cells (NI) within these sampling windows using NIH Image 2000 software (Scion Corporation Beta 4.0.2). The experimenter who counted the immunoreactive cells was blind to all experimental conditions. We defined a threshold corresponding to the grey level above which a cluster of pixels (corresponding to the size of NCM cells nuclei) was considered as signal, and below which it was considered as noise [19]. Because the sampling window used in NCMd, NCMv and CMM presented twice the area of the sampling window used in HIP, the counted NI in HIP was normalized by doubling it. In order to test whether females brain responsiveness is correlated with the subjective salience of sounds, differences in NI values were examined using an analysis of variance for repeated measures with acoustic stimulus (mate vs. non-mate) as between-group factor and anatomical areas as within-group factor (repeated-measures ANOVA, ). The ANOVA was followed by a Fisher PLSD post-hoc test (). In order to test whether females brain responsiveness is correlated with the acoustic structure of sounds, we performed an analysis of variance for repeated measures, with call types as between groups factor and anatomical areas as within group factor (repeated-measures ANOVA, ). To further investigate whether neural responses to calls correlate with the subjective salience or the acoustic structure of calls, we used a multiple regression with NI in each anatomical area as dependent variables and acoustic stimulus and call types as independent factors (). All statistical tests were carried out using Statistica Software version 6.1.
3 Results
3.1 Analysis of acoustic stimuli
Among the 20 acoustic cues measured in the calls, 11 parameters show a significant variation among male individuals (Table 1, Kruskall–Wallis ANOVA, ). All temporal and spectral cues are well individualized, except the frequency at the end of the call () and the frequency of the second peak of intensity (). None of the intensity parameters presents a significant variation among individuals. The individualized parameters allowed performing a PCA to derive composite scores corresponding to the principal components (PC). The discriminant analysis on the ten obtained PC determined five discriminant functions and allowed 100% of discrimination among the six individuals. The hierarchical ascending classification on the discriminant functions allowed defining groups of calls, reflecting their common acoustic cues and defining call types. This method allows quantifying global calls similarities taking into account all the individualized acoustic parameters, but does not tell us on which particular parameters calls can be grouped. At a linkage distance of 30 (squared Euclidean distances) on the tree diagram, the 16 calls can be grouped into four call types (A to D, see Fig. 3), which means four groups of calls that share common acoustic characteristics. Because the limits of these call types do not match the ones of the classes of acoustic stimuli (mate and non-mate), we are now able to test separately the effects of call structure and call identity on ZENK expression.
Statistical analysis of the acoustic parameters of male distance calls
| Variable | Mean | SD | ANOVA Kruskall–Wallis H | p-value | |
| (s) | 0.094 | 0.0416 | 15.84 | 0.014 | ⁎ |
| (s) | 0.014 | 0.0093 | 14.33 | 0.026 | ⁎ |
| (s) | 0.021 | 0.0182 | 13.99 | 0.029 | ⁎ |
| (s) | 0.060 | 0.0333 | 16.96 | 0.009 | ⁎⁎ |
| (Hz) | 642.024 | 318.886 | 15.1 | 0.019 | ⁎ |
| (Hz) | 696.488 | 310.054 | 16.04 | 0.013 | ⁎ |
| (Hz) | 554.231 | 178.229 | 12.15 | 0.058 | NS |
| (Hz s−1) | 54.464 | 37.434 | 12.95 | 0.043 | ⁎ |
| (Hz −1) | 142.257 | 137.449 | 15.23 | 0.018 | ⁎ |
| RMSaver (dB) | 81.3 | 1.319 | 6.34 | 0.386 | NS |
| RMSmax (Pa) | 0.999 | 0.047 | 4.09 | 0.664 | NS |
| (s) | 0.059 | 0.032 | 15.28 | 0.018 | ⁎ |
| RMSmax/RMSaver | 0.012 | 0.001 | 5.75 | 0.452 | NS |
| CG (Hz) | 3438.813 | 446.565 | 14.02 | 0.029 | ⁎ |
| (Hz) | 3105.688 | 591.517 | 13.09 | 0.041 | ⁎ |
| (dB) | 67.144 | 6.3223 | 5.16 | 0.523 | NS |
| /RMSaver | 0.826 | 0.073 | 5.61 | 0.468 | NS |
| (Hz) | 3870.938 | 911.003 | 9.38 | 0.153 | NS |
| (dB) | 61.138 | 7.1102 | 6.33 | 0.386 | NS |
| /RMSaver | 0.752 | 0.087 | 7.73 | 0.258 | NS |
⁎ ;
⁎⁎ ;
NS = Not significant.
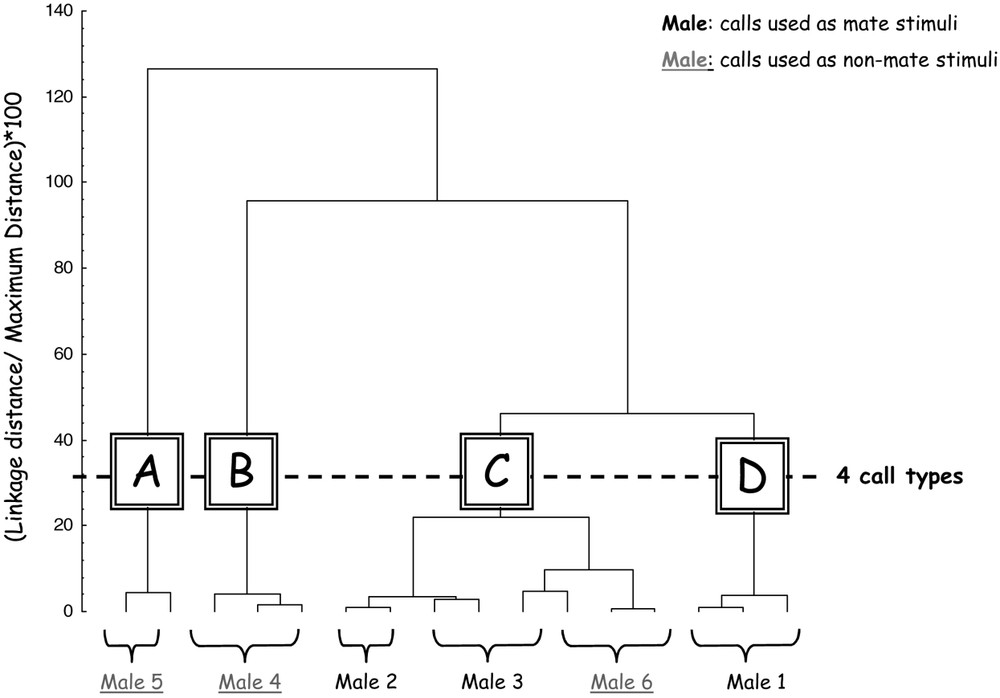
Dendogram obtained by hierarchical ascending classification (method of Ward with squared Euclidean distances). This classification allowed defining four call types (A, B, C, D) reflecting common acoustic cues. Call types do not match the classes of acoustic stimuli (mate and non-mate).
3.2 ZENK immunocytochemistry
The location of the sampling area significantly affects the number of immunoreactive cells NI (within-factor of the repeated-measures ANOVA , ): the ZENK-ICC labelling is stronger in NCMd and CMM than in NCMv and HIP (Fisher PLSD ).
NI is not significantly influenced by the acoustic stimulus (mate vs. non-mate) (repeated-measures ANOVA , ): individual identity of male calls did not influence the global level of ZENK expression in the brain of female subjects. However, the interaction between brain region and acoustic stimulus is significant (repeated-measures ANOVA: , ) (Fig. 4): individual identity of male calls significantly influences the brain pattern of ZENK expression in female subjects. Post-hoc tests reveal that mate stimulus significantly increased ZENK-ICC labelling in comparison with non-mate stimulus (Fisher PLSD ) in HIP, but not in auditory areas (NCMd, NCMv, and CMM). Individual identity of male calls thus affects ZENK expression in hippocampus, but not in auditory areas.

ZENK-ICC labelling (NI) of females challenged with mate stimuli and non-mate stimuli. There is a significant effect of the interaction between the sampling area of quantification and the acoustic stimulus (F=5.38, P=0.002). NI is enhanced in HIP in response to mate calls in comparison with non-mate calls (P=0.002), whereas no influence was observed in other brain areas. Boxes represent standard errors and bars correspond to standard deviation.
Using the call types defined previously based on calls' common acoustic structures, we tested whether female brain responsiveness is correlated with the acoustic structure of sounds used as stimuli. There is no significant effect of call types on the parameter NI (A to D, defined by acoustic cues' similarity in 3.1, repeated-measures ANOVA , ), but a significant effect of the interaction between call type and sampling area (repeated-measures ANOVA: , ) as well as a significant effect of sampling area as within-group factor (repeated-measures ANOVA: , ). As the factor ‘acoustic stimulus’ does, the factor ‘call type’ significantly influences the brain pattern of ZENK expression in female subjects. However, whereas NI is significantly correlated with call type in NCMd and NCMv (respectively , and , ), it is not significantly correlated in the same regions with the acoustic stimulus (respectively, , and , ). Thus, NI in NCMd and NCMv could be predicted by call type (call structure) and not by acoustic stimulus (call individual identity). Although NI in HIP is significantly correlated with call type (, ), it is more strongly correlated with the acoustic stimulus (, ). Thus, acoustic stimulus (call individual identity) allows predicting NI in the hippocampus better than call type (call structure). NI in CMM is not significantly correlated neither with call type (, ) nor with individual identity of male calls (, ).
To sum up, these results give evidence for a genomic response to calls in hippocampus that differentiates more between calls identity (mate versus non-mate) than calls structure.
4 Discussion
In the present paper, we investigated whether male calls with different salience (mate versus non mate) are differently efficient in inducing neural activity in some female brain regions. We observed that mate calls evoked significantly more ZENK expression in female hippocampus than non-mate calls. We did not find out any effect of calls identity on ZENK expression in auditory areas (NCM and CMM). According to the small sample size, our results are preliminary, but give the first evidence to our knowledge that neural responses to calls in hippocampus correlate more with call identity than with call structure.
Previous studies in songbirds have described neuron populations that are selective for particular songs. In male and female adult starlings, operant conditioning results in specific tuning of CMM neurons to learned songs [41]. These CMM neurons show increased electrophysiological response to learned songs compared to novel songs and a subset of this population responds selectively to specific song motifs [42]. In the same operant conditioning design, ZENK expression in CMM is increased in response to novel songs, but also to learned songs, whereas ZENK expression in NCM is enhanced only when the bird hears novel songs [42]. Other studies investigated whether ZENK response in NCM and CMM can be proportional to the behavioural relevance of the stimulus: female white-crowned sparrows show greater ZENK induction in response to their hatch dialect than to foreign dialect, and this induction correlated with female courtship behaviour [20]. Female budgerigars present an increased ZENK expression in NCM and CMM in response to complex songs to which they respond preferentially compared to a simpler song [18]. Female zebra finches learn the characteristics of their father song and form a preference for it that translates into greater ZENK expression in CMM than controls exposed to a novel song [43]. Sexually relevant variations in male song, like song bout duration, induce variations in NCM ZENK response of female starlings [19], and these variations depend on the recent experience of the female [23]. All these results suggested that NCM and CMM could be implicated in the perception processes underlying the behavioural preference for a given song. In the present study, we used female preference for mate call in zebra finches as a paradigm. We found out that neither NCM nor CMM differentiates between the identity of calls, whereas hippocampus does.
In the particular context of tutor song perception in males, it has been previously demonstrated that ZENK expression in NCM and CMM does not depend on the identity of the song stimulus [14,21,44]; conversely, the amount of ZENK expression in the NCM in response to tutor song is correlated with the strength of song learning. Here we found that ZENK expression in female hippocampus is linked to mate recognition. Whereas the NCM of male zebra finches has been implicated in song memory [14,21,44], our results show a possible implication of female hippocampus in retrieval of mate calls memories.
The avian hippocampus appears to be homologous to the mammalian hippocampus [45]. The importance of this brain area in bird memory has been revealed through its implication in spatial memory used for navigation [46] and food retrieval [47]. A role of the hippocampus in acoustic signals' processing [48,49] or in sexual imprinting [50,51] has been previously underlined and stresses the potential modulator effect of hippocampus in fitting the behavioural response to stimulus relevance and context. The hippocampus may be involved in the formation and retrieval of sound memories, and particularly its activation may reflect a contextual memory of the acoustic signal [48,49]. The hippocampus is likely to encode the relevance of the context in which the stimulus appears. Kruse et al. [52] underlined that ZENK activation in the hippocampus could rely on interactions between spatial, acoustic and visual novelty. Contrary to songs, calls often exhibit a highly variable pattern of production [8]. Call emission is regulated by emotional state [53] and we demonstrated in a previous study that call perception is influenced by the current social context [54]. In our experiment, the increased activity of the hippocampus induced by mate calls may sign an increased readiness to respond to mate vocalizations in comparison to the calls of other males.
Acknowledgements
The authors are grateful to Corinne Eyzac, Sabine Palle, Hédi Soula, François Reichardt for their help during the experiments. This work was supported by a Program ‘Émergence’ of the Région Rhône-Alpes and by a grant of the French ‘Agence nationale de la recherche’ (ANR, Birds' voices project). C.V. is supported by a Young Investigator Sabbatical of the Jean Monnet University. N.M. is supported by the ‘Institut universitaire de France’.