

Abridged English version
Introduction
The aim of this work is to conduct a comprehensive review of both experimental evidence and theoretical knowledge of pollen- and seed-mediated dispersal in maize. In 2008, GM maize was the second principal biotech crop grown in the world, occupying 37.3 million hectares at 30% of global biotech crop area. All the GM maize grown around the world exhibits only two major traits, herbicide tolerance (e.g. Monsanto's Roundup Ready) and insect resistance (e.g. Monsanto's MON810, Syngenta's Bt11 and Pioneer's 1507). MON810, Bt11 and 1507 maize varieties have been genetically modified (GM) through the insertion of a gene from the Bacillus thuringiensis (Bt) soil bacterium, to produce an insecticidal protein (δ-endotoxin) for control of some lepidopteran pest insects such as Ostrinia nubilalis (Hbn.), the European corn borer. In 2008, seven of the 27 countries in the European Union officially planted Bt maize on a commercial basis to reach 107,719 hectares. The seven EU countries listed in order of biotech hectarage of Bt maize were Spain (79,269 ha), the Czech Republic (8380 ha), Romania (7146 ha), Portugal (4851 ha), Germany (3173 ha), Poland (3000 ha) and Slovakia (1900 ha). Our work will help to complement the comprehensive review on biological impacts of Cry proteins expressed mainly in insect-resistant Bt maize. An important issue is the possibility of co-existence between GM and non-GM crops. Our review shows that theoretical and experimental data are quite numerous. They lead to propose solutions for co-existence between different cropping systems in a context where the EU sets a threshold of 0.9% of transgenes within food and feed above which all products must be labelled as containing GMOs (Regulation EC 1829/2003).
Methods
Our database is built from selected references from journal sites (such as Ingenta, PubMed, WoK) using the keyword ‘transgene and (plant or crop)’. This data bank is maintained by the analyses of tables of contents of journals received as alerts from collections such as BioOne, Elsevier, Science Direct, Springer, etc. (507 journals), by the monitoring of 352 journals not covered by these collections (these include Chinese journals with automatic on-line translators, proceedings of symposia, etc.), and the by monitoring of 530 other journals. Our database contained 16,703 references as of the 23rd of June, 2009.
For this review the references were sorted using keywords related to agronomical aspects of plant biotechnology. Using the keyword ‘dispersal OR flow OR pollen’, we found 1909 references ranked as follows: 1873 references relate to ‘transgene dispersal’ AND/OR ‘transproteins’ in all GMOs. Among them, 562 publications were selected using the keywords ‘maize OR corn OR zea’. Furthermore, the number of symposia (over 20) shows the interest of scientists for this topic (see Supplementary Electronic Material).
The transgene dispersal
The adventitious presence of transgenes in crops may be due to several factors. The transgene may be:
- • transported by pollen migration. In maize, this migration is due mainly to wind. Indeed, as female flowers do not produce nectar, bees do not visit them. Therefore, they cannot participate in cross-fertilization;
- • transported by seeds that can be mixed with non-GM seeds during harvest and post-harvest processes;
- • perpetuated by the presence of volunteers in fields or by the presence of seeds that have dispersed and germinated out of the GM-field. Volunteers of maize are rare and well controlled by cultural practices.
The pollen-mediated transgene dispersal
Many publications dealt with the dispersal of maize pollen.
The pollen-mediated gene flow from a source-site to a sink-site can be evaluated by counting pollen grains that land on a sticky surface at the sink-site. However, counting pollen grains does not mean necessarily that cross-fertilization occurred due to several factors interfering or not (i.e. mortality, competition, landing of pollen on the female stigma, non-receptiveness of females, abortion in the ovaries, and survival of pollen). Survival of pollen may be limited to less than an hour under certain conditions but in other circumstances, it can survive more than 24 h. To correct these uncertainties, the effective pollen flow can be estimated by measuring the rate of cross-fertilization (crFe) using either morphological or phenotypical markers or the content of the DNA of transgene (transDNA) in plants of the sink-site.
Eventually, since pollen grains can be considered as particles, we can assume that the pollen dispersal is subject to weather conditions (i.e. direction and strength of wind and/or rain).
The European program ‘Sustainable Introduction of GM Crops into European Agriculture’ (SIGMEA) showed that the pollen flow is major in the first meters adjacent to the emission source and decreases greatly as the source-to-sink distance increases. The observed rate of cross-fertilization is 0.98% in the first 10 m from the source-plot, 0.34% between 10 m and 20 m, and 0.11% between 50 m and 60 m.
In the UK, experimental data obtained from 55 plots have be modelled as an exponentially decreasing function with average values less than 0.9% at 25 m, less than 0.3% at 80 m and less than 0.1% at 200 m and beyond. This statistical model allows calculating separation distances between source- and sink-fields to avoid a specific cross-fertilization rate. This helps establish rules of co-existence between GM and non-GM cropping systems.
Thus, in the 2000s, the idea of a mechanistic model came out and models such as MAPOD have been developed. This type of model takes into account cultural practices and climate, and involves GIS data. In a case of intensive presence of Bt maize, two factors must be considered: the source-to-sink distance and synchronized flowering. A rate of presence of fortuitous GM maize (CrFe) is 0% if both factors are considered appropriately. It could reach more than 2% if none of these factors are considered. However, in a recent review (2008) the authors suggested that “models must be retrospectively critiqued based on new field data and knowledge, and not be allowed to become axiomatic. The level of rigor in validating pollen flow models has varied widely, depending in part on the robustness of datasets”. Indeed the authors added “Accurately modelling pollen-mediated gene movement is challenging. Numerous biological, environmental, and crop management factors and their interactions influence the frequency and distance of pollen flow between donor and receptor fields, including type of vector (wind and/or insect), genotype or cultivar, fertility (e.g. male-fertile or male-sterile receptor plants), pollen viability and longevity, synchrony of flowering or pollen production, wind speed and direction, air turbulence/convective air currents, temperature, humidity, and area and plant density of donor and receptor populations. At a landscape or regional scale, additional factors influencing pollen flow are topography, vegetation, distribution and abundance of volunteer and feral populations, and number, shape, and spatial arrangement of pollen donor and receptor fields”.
Concerning man manageable factors modifying the travel and amount of pollen from a source to a sink as well as the cross pollination rates, some data can be cited:
- • an area of separation between source-plots and sink-plots can significantly reduce the amounts of pollen carried into the sink plots. This area is more effective if non-GM maize is sown;
- • discrepancies in flowering dates vary from a cross-fertilization rate of 14.2% with 1 day of flowering lag between source-plot and sink-plot to a cross-fertilization rate of 0.05% with a 20 day flowering lag;
- • respective areas of conventional plots and GM-plots.
Among other factors to be taken into account are the strength and direction of wind which are less manageable. This data is well demonstrated using new methods based on modelling of pollen dispersal. The advantage of modelling using GIS data is that it can predict the possibility of co-existence between cropping systems based on density of GM crops and socio-economic issues, and landscape structure, which allows to prioritize the importance of factors involved in pollen flow and cross-fertilization.
Pollen flow can occur over long distances but obviously with very small amounts of pollen, and with pollen grains which are less and less viable over time of travel. Under certain conditions of wind and temperature, theoretically maize pollen could travel up to 180 km along the upper atmosphere. In models the pollen from a source of a 12-km2 area may emit grains up to 79 km away (0.005 pollen grains/m2). At only a few kilometres (below 5 km) from the source, the cross-fertilization rate is 0.05% if the maize receiver is not castrated, and 0.25% if castrated as no competition occurs with autochthonous pollen. At 5 km from the source, the pollen viability is 30%. This led to highlight the concept of pollen background at the regional scale.
The seed-mediated transgene dispersal
A second way of adventitious presence of transgenes in the production of non-GM maize is the presence of GM-seeds in consignments of non-GM seeds. This mixture can be done through the pollen flow between non-GM seed production and surrounding GM-cultures but also during harvest – transport process which can account for 0% of presence with dedicated harvesters and 0.4% in the case of shared harvesters and lack of cleaning practices.
All this knowledge is used to develop measures that allow co-existence of multiple cropping systems with a specific threshold of cross-fertilization. Thus, it follows that the mixture with GM seeds should be kept under control during the process of harvesting and post-harvesting.
The co-existence between GM and non-GM crops
The first factor used for managing co-existence between GM crops and other cropping systems is the geographical isolation of fields. These separation distances between GM and non-GM maize are widely accepted as an effective measure to reduce cross-fertilization. Several EU countries proposed different separation distances ranging from 25 m to 800 m. However, regulations should be revisited on the basis of scientific criteria including the rate of cross-fertilization. They might help to define separation distances. To keep GM inputs in the final product below the 0.9% labelling threshold prescribed by the EU, separation distances of 20 m and 50 m for silage and grain maize respectively were proposed using statistical data of maize areas and aerial photographs with geographic information systems. Furthermore other important factors that determined cross-pollination are synchrony of flowering and distance between donor-field and receiver-field. In the case of a fully synchronous flowering, a separation distance of 20 m approx. should be sufficient to maintain the presence of GM as a result of pollen flow below the 0.9% labelling threshold.
However, it is necessary to take into account the possibility of adventitious presence of GM seeds in lots of non-GM seed. On the basis of model simulations of case studies, co-existence is technically manageable with a 0.5% threshold of adventitious presence of GM seeds with little or no change of conventional practices.
However, for seed production, the co-existence of non-GM with GM maize should lead to changes in conventional practices, namely the introduction of larger separation distances (from conventional distances of 200–300 m to distances of 400–600 m). For maize, additional measures are necessary to account for climate, landscape and agronomic parameters. The latest versions of models of gene flow take into account spatial distributions of landscapes and agricultural practices.
Based on recent data, some stakeholders require a lower threshold (0.1%) than the 0.9% regulatory labelling threshold to guarantee GMO-free products. For low thresholds (0.1%), isolation distances between GM fields and non GM fields may be difficult to implement and a geographical separation between GM zones and GMO-free zones is viewed as a solution. However, in practice it would be in contradiction with the concept of co-existence mentioned in EU Directive and would restrict the freedom of farmers to opt for GM crops.
However, as on one hand, the situation may vary (climate, winds, area ratio between GM and non-GM fields, landscape and agronomic parameters) and on the other hand taking into account the great progress in models and simulation systems, it seems advisable to replace the national value of separation distance by a value fitting to the local or regional context.
Conclusion
It is essential to conclude a review on mediated-pollen transgene dispersal by indicating methods that avoid this dispersal. We can cite male sterility, restriction technologies, and transgene integration in plastids. Research efforts should be supported to develop these biotechnologies.
1 Introduction
Le maïs (Zea mays L. ssp. mays) est la deuxième plante génétiquement modifiée (PGM) cultivée dans le monde après le soja avec 37,3 million d'hectares représentant 30% des PGM en 2008 [1]. Toutes les variétés de maïs transgénique cultivées dans le monde ont deux traits soit isolés soit en combinaison : la tolérance aux herbicides (e.g. Monsanto's Roundup Ready) ou/et la résistance aux insectes (e.g. MON810 de Monsanto, Bt11 de Syngenta et 1507 de Pioneer). Les variétés MON810, Bt11 et 1507 ont été modifiées par l'insertion du gène cry de la bactérie du sol Bacillus thuringiensis (Bt) qui exprime la transprotéine CryA1b. Elle produit une δ-endotoxine ciblant certains lépidoptères et particulièrement la pyrale (Ostrinia nubilalis Hbn.). En 2008, sept des 27 pays de l'UE ont planté officiellement 107.719 hectares de maïs Bt pour sa commercialisation. Ce sont par ordre d'importance en surface cultivée : Espagne (79.269 ha), République tchèque (8380 ha), Roumanie (7146 ha), Portugal (4851 ha), Allemagne (3173 ha), Pologne (3000 Hha) et Slovaquie (1900 ha) [2].
Une question importante dans l'utilisation des PGM est la possibilité de co-existence entre les systèmes de culture GM et non GM conventionnelle ou biologique. Afin de pouvoir garantir la liberté de choix entre les systèmes de culture PGM et non PGM, et sachant qu'il est impossible de garantir une étanchéité des filières, l'UE a établi un seuil de présence fortuite (PrFo) de PGM de 0,9% dans les cultures ou leurs produits non GM (Régulation EC 1829/2003). Ce seuil qui est un compromis de politiques publiques permet cependant d'asseoir cette question sur des faits scientifiques, théoriques et expérimentaux. En deçà de ce seuil, les cultures ou les produits ne sont pas considérés comme GM, et au-delà ils sont étiquetés comme contenant des OGM. A noter que ce seuil existe également hors de l'UE et est variable suivant les pays. Ainsi l'Australie, la nouvelle Zélande et le Brésil ont un taux limite de 1%, celui du Japon est de 5%, d'autres pays (USA par exemple) n'ont pas de seuil limite parce que ces pays considèrent la culture de PGM au même titre que celle des autres variétés.
Dans le cas de la production de semences, le seuil de PrFo, au niveau européen, est en cours de discussion.
Le but de ce présent travail est de faire un bilan complet des connaissances expérimentales et théoriques concernant les capacités de dispersion des transgènes du maïs Notre travail permet ainsi de complémenter la revue exhaustive de Icoz et Stotzky [3] sur l'impact biologique des protéines Cry exprimées en particulier dans le maïs Bt. Nous nous sommes focalisés sur le maïs Bt, qui est le seul maïs GM autorisé commercialement à la culture dans l'UE. De plus, nous montrons que les données expérimentales et théoriques sont relativement nombreuses et conduisent à proposer des solutions pour faire co-exister les différents systèmes de culture.
2 Démarche méthodologique
Notre banque de données bibliographiques est construite à partir de références sélectionnées dans Ingenta, PubMed, WoK grâce au mot clé « transgen* AND (plant* Or crop*) ». Cette banque est tenue à jour par l'analyse des sommaires de journaux scientifiques reçus en alerte informatique dans les collections telles que BioOne, Elsevier, Science Direct, Springer, etc. (507 journaux), avec un suivi de 352 journaux non pris en compte par ces collections (il s'agit notamment des revues chinoises en traduction automatique, de proceedings de symposium, etc.) et un suivi de 530 autres journaux. Notre banque de données contient 16.703 références en juin 2009.
Pour cette revue systématique, les références sont triées en utilisant des mots-clés relatifs aux aspects agronomiques des biotechnologies végétales. Ainsi, avec le mot-clé « dispersal OR flow OR pollen », nous obtenons 1909 références dont 1873 sont relatives à la dispersion de transgène ( « transgene dispersal ») et/ou aux transprotéines ( « transproteins ») dans toutes les PGM. La répartition des revues dans lesquelles ces 1873 références sont publiées est présentée dans le Tableau 1. Parmi elles, 562 publications sont sélectionnées avec le mot-clé « maize OR corn OR zea » (Tableau 2). Depuis 2005, nous avons recensé plus de 20 symposia et conférences se rapportant entièrement ou en partie à la dissémination des transgènes. Le nombre de symposia montre l'intérêt des scientifiques pour ce sujet. La majorité de ces références est recensée dans le supplément informatique de cet article dans lequel les références comprennent un résumé et des commentaires des auteurs ou autres commentaires des articles citant la référence.
Chronologie de parution des 1873 publications portant sur la dispersion des transgènes dont celles concernant spécifiquement le maïs.
Chronology of publication of 1873 references on the dispersal of transgenes, including those specifically maize.
| Année | Nombre total de publications | Nombre de publications sur le maïs |
| 1938 à 1995 | 120 | 25 |
| 1996 | 28 | 1 |
| 1997 | 22 | 4 |
| 1998 | 61 | 7 |
| 1999 | 108 | 10 |
| 2000 | 66 | 14 |
| 2001 | 83 | 20 |
| 2002 | 127 | 37 |
| 2003 | 167 | 55 |
| 2004 | 170 | 54 |
| 2005 | 221 | 81 |
| 2006 | 216 | 82 |
| 2007 | 330 | 114 |
| 2008 | 154 | 58 |
| Total | 1873 | 562 |
Répartition des 562 publications par type de revue (6 références n'ont pas pu être classées).
Distribution of 562 publications by type of journal (6 references have not been ranked).
| Type de revue | Nombre de publications |
| Revues à comité de lecture | 198 |
| Revues sans comité de lecture | 26 |
| Revues (dont le comité de lecture n'est pas précisé) | 5 |
| Proceedings de symposium ; Conférences | 219 |
| Thèses | 3 |
| Ouvrages | 6 |
| Rapports officiels (ONU, OCDE, UE, Gouvernements, etc.) | 36 |
| Page web (universités, rapports de recherches, etc.) | 35 |
| Pages web (associations, etc.) | 28 |
3 La dispersion biologique des transgènes
La dispersion du pollen préoccupe les chercheurs depuis de longues années, ainsi l'université de l'Oklahoma a publié en 1950 un bulletin technique détaillé [4] souvent cité dans les publications scientifiques récentes (voir supplément électronique). La présence fortuite (PrFo) de transgène dans des cultures non GM peut être due à plusieurs facteurs. Le transgène peut être :
- • transporté par la migration du pollen. Chez le maïs, cette migration est avant tout anémophile mais les abeilles (insectes pollinisateurs) peuvent aussi récupérer du pollen de maïs, cependant, comme elles ne visitent pratiquement pas les fleurs femelles de maïs, qui ne produisent pas de nectar, elles ne peuvent qu'occasionnellement participer à la fécondation croisée (FeCr) en laissant tomber des grains de pollen sur les soies ;
- • transporté par les graines qui peuvent être mélangées aux graines non GM au cours de la filière récolte, transport, stockage ;
- • perpétué par la présence de repousses dans la parcelle ou de graines qui se sont dispersées et qui ont germé hors de la parcelle GM. Cependant, chez le maïs, les repousses sont rares dans les parcelles et hors-champ [5] et peuvent être maîtrisées par les techniques culturales.
3.1 La dispersion des transgènes via le flux de pollen
Le flux de pollen entre un site source qui émet le pollen et un site puits qui le reçoit peut être évalué en dénombrant les grains de pollen qui se fixent sur une surface collante au niveau du puits. Mais le dénombrement des grains de pollen ne signifie pas nécessairement que la FeCr ait eu lieu, à cause d'aléas tels que la mortalité, la compétition, l'atterrissage du pollen ailleurs que sur les stigmates femelles, la non réceptivité des organes femelles et l'avortement dans les ovaires. De plus, dans certaines variétés GM comme le maïs Mon810, un grain de pollen sur deux ne porte pas de transgène cry [6,7]. Pour corriger ces aléas, on peut estimer le flux efficace :
- • en dosant l'ADN du transgène (transDNA) dans les pieds de maïs de la parcelle puits ; dans ce cas le ‘transDNA’, qu'il soit d'origine Bt ou tolérant à un herbicide, renseignera de la même façon pour la problématique de la dispersion des transgènes [8,9]. Pour différentes raisons, il peut être difficile de convertir les analyses quantitatives de transDNA avec les analyses qualitatives [7,10]. De plus, la plupart des mesures actuelles sont faites sur la base d'un seul transgène par PGM. Or, comme on utilise de plus en plus de PGM comprenant un empilement de transgènes, il faut se demander comment tenir compte de cet empilement dans la détermination du taux de FeCr [11].
- • en utilisant des marqueurs morphologiques ou phénotypiques de type couleur de graines [12].
En mécanique des fluides, le grain de pollen est considéré comme une particule, on peut alors concevoir que la dispersion de pollen (flux de pollen) est soumise aux aléas météorologiques (direction et force des vents, régime des pluies), ce qui a été vérifié expérimentalement ([13,14] pour revue).
Parmi les publications, on relève des revues bibliographiques bien documentées [7,15] ou des rapports et comptes-rendus de symposium centrés sur des résultats expérimentaux obtenus par l'intermédiaire de programmes de recherches internationaux. Sans être exhaustifs ici, on peut citer les travaux du programme européen SIGMEA (‘Sustainable Introduction of GM crops into European Agriculture’) [16]. Les résultats obtenus montrent que le flux de pollen est important dans les premiers mètres jouxtant la source d'émission et que ce flux diminue très fortement quand la distance source – puits augmente. Cette expérimentation met en jeu de nombreux sites dans lesquels les parcelles sources (dont la surface peut atteindre 23 ha) sont entourées de parcelles puits, les taux de FeCr sont de 0,98% à 0–10 m, 0,34% à 20–30 m et 0,11% à 50–60 m de la parcelle source [17]. Autre exemple, dans des conditions réelles en Espagne, une parcelle de maïs conventionnel entourée par 4 parcelles de maïs Bt présente un taux de fécondation FeCr de 2,76% dans les 10 premiers mètres, puis de 0,69% entre 10 et 20 m et enfin de 0,3% entre 20 et 30 m [17]. De nombreuses publications donnent des résultats tout à fait concordants, quelques articles sont parus en 2007 [19–22] et en 2008 [23] (pour une vue plus exhaustive, voir le supplément électronique).
Récemment, de nombreuses publications ont été analysées en tenant compte des caractéristiques expérimentales qui peuvent expliquer les plages de variation des résultats enregistrés [24]. Les auteurs ont établi qu'en moyenne la dispersion du pollen mesurée par le taux de FeCr peut être représentée par le graphique (Fig. 1). En résumé, les résultats montrent que jusqu'à 10 m des parcelles sources les résultats sont très hétérogènes avec une moyenne de 5,72% de FeCr. Ensuite, la FeCr diminue fortement pour atteindre 0,35% entre 10 et 25 m, 0,23% entre 25 et 50 m, et 0,19% au-delà de 50 m sans atteindre 0%. Des résultats du même ordre ont été trouvés en Espagne [18]. Un travail relativement voisin d'analyse des données bibliographiques et de résultats expérimentaux a été effectué dans le cadre du programme européen SIGMEA [25]. Celui-ci, qui présente des résultats portant sur des distances plus grandes entre les parcelles GM et non GM, montre que le flux de pollen peut atteindre plusieurs kilomètres (Fig. 2). A ces grandes distances, les quantités de pollen mises en jeu sont très réduites et concernent des grains de pollen qui sont de moins en moins viables avec le temps de parcours. Dans certaines conditions de vent et de température, théoriquement le pollen de maïs pourrait se déplacer jusqu'à 180 km en empruntant la haute atmosphère [26]. Par modélisation il est montré que le pollen d'une source qui s'étend sur 11 km peut émettre des grains de pollen jusqu'à 79 km (0,005 grains de pollen/m2) [27]. On ne devrait plus trouver de pollen à 95 km. En Suisse, dans des conditions de faible pression de parcelles sources et pour des distances séparant les parcelles sources et puits allant de 50 à 4500 m, les taux de FeCr ne sont jamais supérieurs à 0,02% (Fig. 2B) [12]. De plus, dans 89% des cas, le taux de FeCr résulte d'un seul événement de fécondation sur un seul épi [28]. Des résultats du même ordre ont été trouvés en Espagne [18].
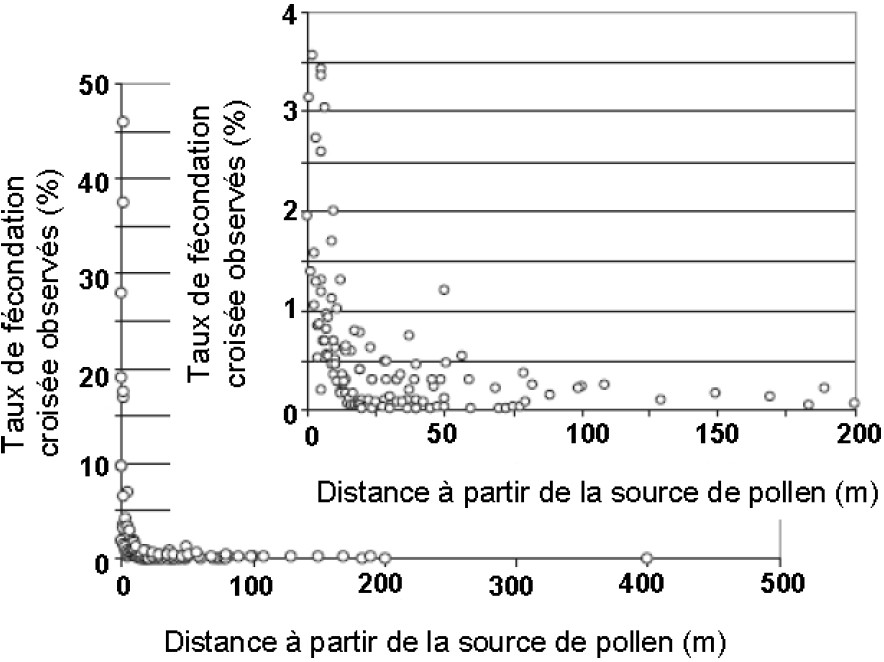
Taux de fécondation croisée observés (FeCr en %) selon la distance à partir de la source de pollen (en mètres) et agrandissement de la figure sur une échelle réduite d'après [24].
Rate of observed cross-fertilization (CrFe in %) depending on the distance from the pollen source (in meters) from [24]. The box represents a magnification of the original graph with a limited scale of the respective axis.
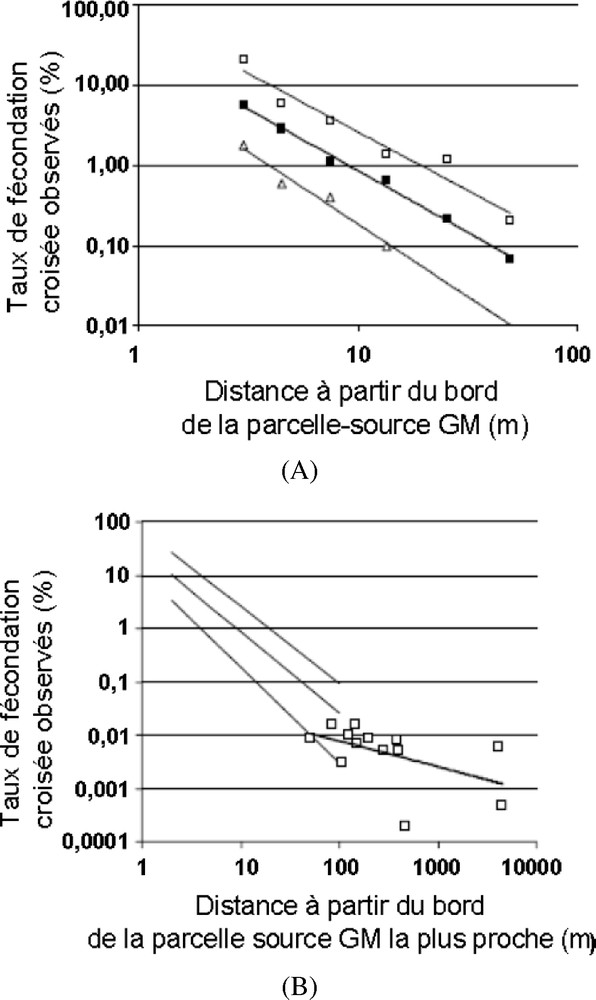
Taux de fécondation croisée observés (FeCr en %) et droites de régression selon la distance (en mètres) du bord de la parcelle source génétiquement modifiée (GM) (A d'après [16] ou de la parcelle source GM la plus proche (B d'après [28]).
Rate of observed cross-fertilization (CrFe in %) and linear regressions according to the distance (in meters) from the edge of the genetically modified (GM) source plot (A from [16]) or from the nearest GM source plot (B from [28]).
Ces chiffres sont à relativiser par rapport au fait qu'en arrivant à destination ces quelques grains de pollen vont être en compétition pour la fécondation avec les grains de pollen autochtone beaucoup plus nombreux. En effet, le maïs produit environ 4,5 à 25 millions de grains de pollen par plante [29], alors que le pollen migrateur prélevé entre 1000 m et 2000 m d'altitude représente une concentration de 1 grain/m3 en moyenne, ce qui donne 10 millions de grains par hectare de culture [30]. Cette compétition indique qu'à quelques kilomètres de la source (moins de 5 km), le taux de FeCr est de 0,05% si le maïs récepteur est non castré et de 0,25% s'il est castré, car il n'y a pas de compétition avec le pollen autochtone [31]. Ces données doivent être rapportées à celles qui estiment qu'à 5 km de la source la viabilité du pollen est de 30% [32]. Ceci a conduit à mettre en évidence la notion de bruit de fond pollinique au niveau régional [13,32]. Bien que ce bruit de fond soit faible [16,33], il faudra en tenir compte si les surfaces en maïs GM deviennent relativement importantes par rapport aux surfaces de maïs conventionnel.
La survie du pollen est aussi un facteur important de FeCr entre parcelles GM et non GM de maïs. Elle dépend fortement des conditions environnementales et en particulier de l'humidité atmosphérique [34,35]. Les travaux expérimentaux ont établi qu'en conditions « normales » le pollen de maïs ne survit qu'entre une heure [36] et quatre heures [37] après sa libération dans l'atmosphère. Cette survie est fonction de la distance de la source émettrice et de la hauteur de capture de ce pollen [38]. Ainsi, au niveau du sol il y a 92% de pollen viable à 0,5 m de la source et 49% à 4 m, en revanche à 2,1 m de hauteur il y a 59% de pollen viable à 0,5 m de la source et 28% à 4 m. Un bilan bibliographique [26] fournit des résultats comparables mais ajoute qu'en conditions très favorables le pollen peut survivre pendant 24 heures et que, conservé au froid, son activité peut être maintenue pendant 9 jours. L'impact de la force et de la direction des vents sur la dispersion du pollen est important [20,39] comme le montre le Tableau 3. Cet impact est bien imagé avec les nouvelles méthodes de modélisation de la dispersion du pollen [36,40–42] comme le montre la Fig. 4 illustrant la déformation du nuage pollinique dans la direction du vent.
Taux de fécondation croisée (FeCr) observé en fonction de la distance source-puits, de la direction du vent et de la synchronisation ou non de la floraison entre source et puits [11].
Cross-fertilization rate (CrFe) observed depending on the source-sink distance, the wind direction and whether or not synchronization of flowering between source and sink [11].
| Parcelle | Année | Distance | Vent | Floraison | FeCr |
| W7 | 2004 | 52 | + | −7 | 0,009 |
| W12 | 2004 | 85 | + | = | 0,015 |
| W2 | 2003 | 105 | - | −4 | 0,003 |
| W3 | 2003 | 125 | - | −1 | 0,010 |
| W10 | 2004 | 149 | + | −1 | 0,016 |
| W5 | 2003 | 150 | + | −1 | 0,007 |
| W1 | 2003 | 200 | - | −1 | 0,009 |
| W9 | 2004 | 287 | - - | −4 | 0,005 |
| W11 | 2004 | 371 | + | −5 | 0,008 |
| W8 | 2004 | 402 | - | −1 | 0,005 |
| W13 | 2004 | 458 | + | −7 | 0,0002 |
| W6 | 2003 | 4125 | + | −1 | 0,006 |
| W14 | 2004 | 4440 | + | −2 | 0,0005 |
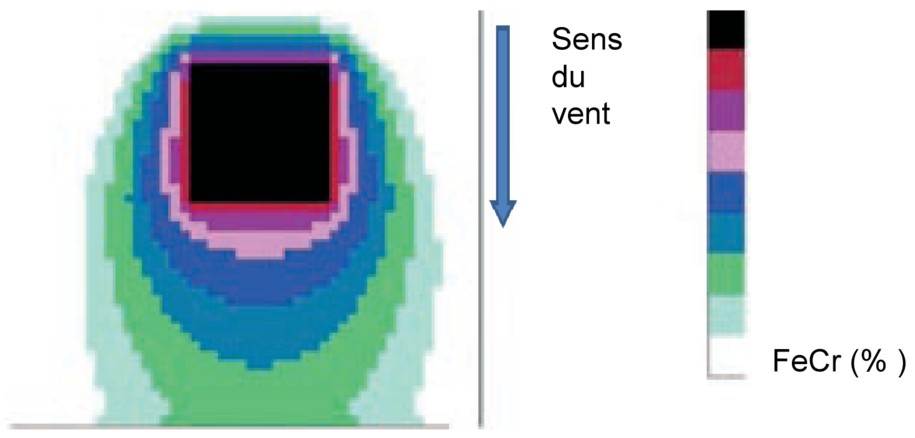
Modélisation du taux de fécondation croisée (FeCr en %) d'une parcelle puits avec une source GM au centre et floraison synchrone [40]. Légende : L'échelle de taux de FeCr (%) va de 50% pour le noir à 0% pour le blanc respectivement (en passant par 20%, 10%, 5%, 2,5%, 1%, 0,5% et 0,1%).
Modelisation of the rate of cross-fertilization (CrFe in %) of a sink plot with a GM source in the center and with a synchronous flowering [40]. Legend : The scale of CrFe rate (%) ranges from 50% to 0% black to white respectively (through 20%, 10%, 5%, 2.5%, 1%, 0.5%, and 0.1%).
Le décalage de floraison entre les parcelles sources et puits est un facteur important influençant la présence fortuite de transgènes cry dans les parcelles de maïs conventionnel. Ce type de données est confirmé par les résultats obtenus dans le cadre du programme européen SIGMEA (voir compte rendu dans [16,43]). Ils indiquent que dans une situation de présence intensive de maïs Bt, deux facteurs doivent être pris en compte : la distance source puits et la synchronisation de la floraison. De nombreuses parcelles puits entourées de parcelles sources ont un faible taux de PrFo, voire un taux de 0%, si ces deux facteurs sont appliqués. En revanche, le taux de PrFo peut atteindre plus que 2% si on n'utilise aucune de ces conditions [44]. D'autres données expérimentales confirment ces résultats [45–47]. Ainsi un taux de FeCr de 14,2% est observé avec un décalage de floraison d'un jour entre parcelle source et parcelle puits, en revanche si ce décalage de floraison est de 20 jours le taux de FeCr chute à 0,05% [47]. Des différences de dates de floraison, allant de 5 à 15 jours, peuvent être obtenues en faisant des semis décalés [48].
3.1.1 Les modèles statistiques
De nombreuses publications montrent qu'on peut modéliser le flux de pollen. On peut citer le travail au Royaume-Uni [49], qui montre que les données expérimentales obtenues dans 55 parcelles sont raisonnablement modélisables sous forme de décroissance exponentielle dont les valeurs moyennes sont inférieures à 0,9% à 25 m, inférieures à 0,3% à 80 m, inférieures à 0,1% à 200 m et au-delà (Fig. 3). En fait, ce type de modèle statistique permet de calculer des distances de séparation qui devraient être respectées entre parcelles sources et parcelles puits pour éviter de dépasser un taux de fécondation croisée donné [31]. Mais les types de données utilisées occultent des facteurs importants qui modifient la dispersion du pollen comme la direction et la force du vent ou qui modifient les capacités de fécondation du pollen comme sa durée de vie après libération par les soies ou le décalage de floraison entre parcelles sources et parcelles puits.
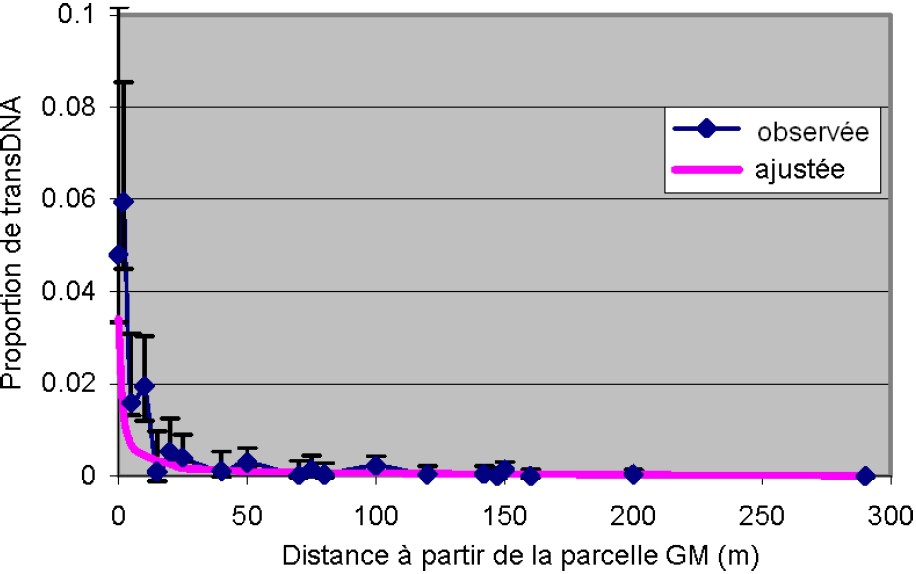
Proportions de transDNA observées dans chaque échantillon et ajustées selon la distance à partir de la parcelle source GM (en mètres) et courbe théorique ajustée (d'après [49]).
Proportions of transDNA (GM DNA) detected in each sample observed and fitted according to the distance from GM source plot (in meters) and the fitted theoretical curve (from [49]).
En établissant un indice fondé sur deux variables importantes de la migration des transgènes (distance entre parcelles sources et parcelles puits et date de floraison), les taux de présence fortuite de maïs GM pourraient être prédits, grâce à des modèles adéquats, avec une précision suffisante pour établir des règles de co-existence [6,18].
3.1.2 Les modèles mécanistiques
La nécessité de prendre en compte en même temps les facteurs biologiques et environnementaux impactant la dispersion des transgènes et la FeCr et la gestion des cultures [43] a fait émerger l'idée, après les années 2000, d'utiliser une modélisation mécanistique [50]. C'est ainsi que des modèles tels que MAPOD (Matricial Approach to POllen Dispersal) ont été mis au point [51]. Ce type de modèle tient compte du système de culture et du climat et peut associer des données GIS (Geographical Information System) [7,52,53]. L'avantage de la modélisation et de son association avec les données GIS est qu'elle permet de faire des simulations pour prédire la possibilité de co-existence entre systèmes de culture en fonction des pressions de cultures GM et de l'environnement socio-économique [15,19,54,55] et de la structure du paysage, ce qui permet de hiérarchiser l'importance des facteurs intervenant dans le flux de pollen et dans la fécondation croisée [56]. Grâce aux progrès de la modélisation et de la simulation, des tableaux de FeCr ont été établis en fonction non seulement de la distance entre parcelles sources et parcelles puits mais aussi en fonction de la proportion des surfaces des parcelles GM et non GM, de la position des parcelles par rapport à la direction du vent et d'un décalage possible de la floraison [7]. Ces tables de décision montrent que pour des parcelles non GM placées contre le vent par rapport à des parcelles GM le taux de 0,9% n'est jamais dépassé et un taux inférieur à 0,1% peut être assuré dans tous les cas (à l'exception d'un seul) avec des distances d'isolement de 20 m. En revanche, pour des parcelles non GM placées sous le vent par rapport aux parcelles GM, le taux de 0,9% ne sera atteint qu'à condition de combiner distance de séparation et décalage de floraison. Voir ces tables de décision dans le supplément informatique à la référence [7].
Toutefois, dans une étude récente [57] les auteurs ont suggéré que « les modèles doivent être critiqués a posteriori sur la base de nouvelles observations de terrain et de nouvelles connaissances, et ne doivent pas devenir axiomatiques. Le niveau de rigueur pour valider les modèles de flux de pollen varie largement en fonction en partie de la robustesse des jeux de données », car ajoutent les auteurs « précisément la modélisation de mouvement de flux de gènes via le pollen est un défi. De nombreux facteurs biologiques, environnementaux et de gestion des cultures et de leurs interactions influencent la fréquence et la distance du flux de pollen entre les champs donneur et récepteur, y compris le type de vecteur (le vent et/ou les insectes), du génotype ou du cultivar, de la fécondité (par exemple, les plantes réceptrices mâles fertiles ou mâles stériles), la viabilité et la longévité du pollen, la synchronisation de la floraison ou de production de pollen, la vitesse et la direction du vent, la turbulence/les courants de convection de l'air, la température, l'humidité, ainsi que la surface et la densité des de la populations donneuses et réceptrices. A l'échelle d'un paysage ou à l'échelle régionale, d'autres facteurs influençant les flux de pollen sont la topographie, la végétation, la répartition et l'abondance des populations hors champ et des repousses, et le nombre, la forme et la disposition spatiale des champs donneurs et récepteurs de pollen ».
Au niveau régional, les rapports des surfaces entre maïs GM et non GM peuvent modifier les résultats de simulations de FeCr obtenus en ne tenant compte que des résultats mesurés à partir de la présence fortuite de champ à champ [32]. Cette prise en compte introduit la notion de bruit de fond régional qui sera d'autant plus important que les surfaces cultivées en maïs GM seront plus grandes.
L'ensemble des connaissances exposées dans les paragraphes 3.1.1 et 3.1.2 permet de déterminer la meilleure façon de cultiver les maïs GM et conventionnel pour qu'un taux de fécondation croisée dû à la migration du pollen soit en deçà du taux autorisé. Elles devraient donc être exploitées pour établir des mesures qui permettent une co-existence de plusieurs systèmes de culture avec une limite donnée de fécondation croisée [11,55,58].
3.2 La dispersion des transgènes via les semences
Une seconde voie de présence fortuite de transgènes dans les productions de maïs non transgéniques est la présence de semences GM dans les lots de semences non GM [11].
Ce mélange peut se faire par l'intermédiaire du flux de pollen entre production de semences non GM et cultures GM avoisinantes (voir données dans le paragraphe 3 et [7]). Depuis longtemps les producteurs de semences de maïs se sont préoccupés de la pureté des semences surtout avec l'utilisation de maïs hybrides dès les années 1950. La séparation des cultures de maïs semences peut être aménagée pour éviter ce mélange [24,46].
Le mélange peut aussi avoir lieu lors du processus de récolte, de séchage et de stockage du maïs. Plusieurs méthodes de séparation des filières récoltes et post-récoltes, dans le temps ou dans l'espace, sont préconisées pour éviter ce problème [59–61]. D'après une analyse bibliographique et des avis d'experts, la filière récolte – transport intervient pour 0% de présence fortuite avec des récolteuses dédiées à la ségrégation des filières GM et non GM et pour 0,4% dans le cas d'un partage des récolteuses et en absence de pratiques de nettoyage.
4 La coexistence entre cultures GM et non GM
Dans les pays qui considèrent que les cultures GM sont différentes des cultures non GM, il est nécessaire d'établir des seuils de PrFo sans lesquels les agriculteurs n'auraient pas le droit de choisir librement leur système de culture. Dans l'UE, la réglementation indique qu'au-dessus d'un seuil de 0,9% de PrFo tout produit, qu'il soit issu de l'agriculture conventionnelle ou biologique, doit être étiqueté comme contenant du matériel génétiquement modifié. Au niveau agronomique et en ce qui concerne le maïs, ce seuil de 0,9% peut être atteint par deux voies complémentaires :
- a) la présence de semences GM dans des lots de semences non GM qui est due d'une part au fait que les parcelles de production de semences se trouvent dans des zones de culture de maïs GM pour la consommation et d'autre part au fait que le matériel de récolte et de transport post-récolte des semences peuvent être les mêmes pour les parcelles GM et non GM ;
- b) la fécondation croisée entre pollen GM et épis non GM due au flux de pollen GM entre parcelles de culture de maïs destiné à la consommation.
4.1 Maîtrise de la présence fortuite due aux semences
L'UE pourrait établir un seuil de 0,5% de présence de graines GM dans les lots de semences conventionnelles pour pouvoir classer ces lots en tant que semences non GM [24].
La maîtrise de la présence fortuite dans les parcelles de production de semences est bien au point actuellement suite aux règles de pureté de la production de semences de maïs hybrides F1. En ce qui concerne le mélange dans le matériel de récolte et de transport, le seuil de 0,5% est facilement gérable, car, sans prendre de précautions particulières [58,60,61], la présence fortuite de semences GM est de 0,4% dans les lots de semences non GM. En adoptant des matériels de récolte et de transport dédiés au maïs GM, cette filière intervient pour 0% de PrFo.
4.2 Maîtrise de la présence fortuite due au flux de pollen entre parcelles de maïs de consommation
Pour satisfaire un taux maximum de 0,9% de PrFo de maïs GM dans des produits de consommation non GM, il faut qu'au niveau des récoltes ce taux soit au maximum égal à la différence (0,9% – le taux de PrFo dans les semences). En admettant que, dans le pire des cas, on ait un taux de PrFo de 0,4% [58,60], le taux maximum de PrFo dans les parcelles puits de maïs conventionnels ne doit pas dépasser 0,5%. Par exemple, dans une région comme Poitou-Charentes, le seuil de 0,9% de PrFo de GM dans les cultures conventionnelles n'est pas atteint dans la plupart des parcelles de maïs en utilisant des pratiques courantes et à condition qu'il y ait au plus 0,5% de semences GM dans les semences non GM [7]. Ce seuil de 0,5% de présence fortuite dans les semences serait la proposition de présence fortuite admise par l'UE [24].
Plusieurs auteurs ont fait une analyse critique des publications concernant la dispersion des transgènes via les flux de pollen [7,12,24,28,46,62,63]. Tous ces auteurs s'accordent à dire que, dans la grande majorité des cas, un taux inférieur à 0,9% de PrFo dans les produits de culture conventionnelle de maïs, pour des maïs hétérozygotes comportant un seul transgène, peut être atteint à condition de laisser une distance de séparation minimale de 10 à 50 m entre parcelles GM et non GM. Ainsi dans 33 publications analysant la présence fortuite de matériel GM au-delà de 50 m, une seule mentionne un taux supérieur à 0,5% (il est précisément de 0,55% à 57 m) [24]. Dans cet intervalle de 10 à 50 m, la distance exacte à laisser entre parcelles GM et non GM dépend de facteurs tels que :
- (i) la pureté des semences non GM ;
- (ii) les caractéristiques des parcelles GM et non GM (formes, surfaces, etc.), leur distribution et leur environnement (haies, etc.) dans le paysage ;
- (iii) le pourcentage de surfaces en maïs GM par rapport à la surface totale cultivée en maïs dans une région donnée ;
- (iv) le type de culture ;
- (v) les différences de dates de semis et de dates de floraison entre parcelles GM et non GM ;
- (vi) et les conditions météorologiques [10].
Plusieurs de ces facteurs sont pris en compte dans les modèles de simulation. En revanche, un certain nombre d'entre eux font encore l'objet de travaux de recherches (hétérogénéité paysagère, etc.) [50]. Malgré ces données expérimentales et scientifiques, les distances d'isolement préconisées officiellement varient entre 25 et 800 m [64] (Tableau 4). Il est clair que l'établissement de distances d'isolement unique pour un Etat-membre oblige à déterminer cette distance comme celle permettant d'éviter une présence fortuite de 0,9% dans les conditions les plus favorables au transport de pollen des parcelles sources de transgènes vers les parcelles puits de culture conventionnelle ou biologique (vent fort en direction des parcelles puits, floraison synchrone, fort pourcentage régional de cultures GM, taille réduite des parcelles puits, etc.). Or, on sait que l'agriculture d'un Etat-membre donné se caractérise par une diversité de situations agronomiques, climatiques et géographiques régionales, voire sub-régionales, importantes. Pour tenir compte de cette diversité, plusieurs auteurs préconisent d'établir des distances d'isolement adaptées au contexte local [10,15,62,65]. La prise en compte du contexte local voire régional permettrait d'exploiter pleinement et habilement la modélisation et les outils d'aide à la décision qui en découlent, ce qui n'est pas le cas avec une distance d'isolement déterminée à un niveau national.
Distances d'isolement des parcelles de maïs GM préconisées dans l'Union européenne par les Etats-membres (d'après [13]).
Isolation distances of GM maize plots recommended by Members State of the European Union (from [13]).
| Etat-membre | Distance d'isolement préconisée (en mètres) |
| Suède (maïs fourrager) | 15 |
| Suède (maïs grain) | 25 |
| Pays-Bas | 25 |
| France | 50 |
| Irlande | 50 |
| Espagne | 50 |
| République tchèque | 70 |
| UK (maïs fourrager) | 80 |
| UK (maïs grain) | 110 |
| Allemagne | 150 |
| Belgique (Wallonie) | 200 |
| Danemark | 200 |
| Pologne | 200 |
| Portugal | 200 |
| Slovaquie | 200 |
| Hongrie | 400 |
| Luxembourg | 800 |
5 Conclusion
La dispersion des transgènes est une question théorique en soi qui a des répercussions sur des questions d'ordre plus pratique comme : (i) l'introgression des espèces sauvages par les espèces cultivées apparentées ; et (ii) la présence fortuite de transgène et vice versa dans des systèmes de culture conventionnelle ou biologique, c'est-à-dire la co-existence des systèmes de culture.
En ce qui concerne l'introgression de transgènes dans les espèces sauvages, cette question ne se pose pas pour le maïs en Europe puisque les espèces sauvages apparentées au maïs telles que la téosinte du pool génique primaire sont absentes de l'aire du culture du maïs. Mais cette question est de portée plus générale parce qu'on devrait la poser pour toute variété ou espèce nouvellement introduite dans un système de culture comme cela a été fait pour le riz [65]. Ainsi, dans certains agro-écosystèmes, comme celui du riz où la forme sauvage vit en sympatrie avec la forme cultivée, les deux formes pourraient alors échanger des gènes, et si les gènes s'intègrent dans le génome hôte, si ils s'y expriment et si les descendants hybrides ont une bonne valeur sélective [66], le génome de la forme sauvage pourrait alors être modifié. Cet échange de gènes pourrait avoir lieu dans le cas de la téosinte et du maïs au Mexique puisque les deux formes y vivent sympatriquement. Depuis longtemps, en tout cas avant l'introduction des maïs GM, on sait grâce à des données moléculaires qu'il existe bien un flux de gènes via le pollen entre le maïs et la téosinte et que l'introgression du maïs et de la téosinte se manifeste dans les deux sens, mais à des niveaux faibles [67,68].
Concernant le taux de présence fortuite de 0,9% de transgène dans les cultures non GM, les données citées dans ce présent travail et dans les publications commentées dans le supplément électronique indiquent que des distances de séparation de 20 m suffisent pour ne pas atteindre ce taux, surtout si (i) dans ces espaces de séparation on cultive du maïs non GM qui sera considéré comme GM [21], et si (ii) on décale les dates de floraison entre parcelles sources et parcelles puits. Ces conclusions ne valent que pour l'Europe. Elles sont appuyées par un rapport CAST [69] qui résume la situation en citant Messéan [46] lequel indique qu'un taux de FeCr de 0,9% est aisément gérable, et que pour ne pas dépasser un taux de 0,5% il faut installer des clauses restrictives de distances de séparation. Ces conclusions sont aussi soutenues par celles de Weekes et al. [31] qui ont synthétisé des données obtenues dans 55 sites expérimentaux et après l'analyse moléculaire du transgène de 1055 échantillons de maïs. Cependant la facilité avec laquelle on maîtrisera le taux de 0,9% pour l'étiquetage dépend de la pureté des semences. Jusqu'à 0,5% de présence de GM dans les semences non GM, le taux de 0,9% est gérable de parcelle à parcelle [7]. Mais certaines parties prenantes de la filière maïs raisonnent sur un taux de 0,1% dans les récoltes ce qui implique bien évidemment un taux identique ou inférieur dans les semences ainsi qu'une gestion avec des zones dédiées soit à la culture de maïs GM soit à la culture de maïs non GM [11]. Si le taux de 0,1% est adopté, le concept de co-existence sera donc vidé de son sens et beaucoup d'agriculteurs seront privés de leur liberté de choix de produire ou pas du maïs GM inscrite dans la Directive 2001/18.
Il nous semble indispensable de conclure un article sur la dispersion des transgènes via le pollen en indiquant des méthodes qui mettent pratiquement à l'abri de cet aléa [11] comme la stérilité mâle cytoplasmique [70], les technologies de restriction [71], l'épigénétique et les RNAi [72] ou/et l'intégration des transgènes dans les plastides [73–75]. Des efforts de recherche, tels que ceux affectés au programme de recherche « Transcontainer » (http://www.transcontainer.wur.nl/uk/), devraient être soutenus pour développer par les biotechnologies ces caractères qui minimiseraient au mieux le flux (ou la dispersion) de transgènes via le pollen.
Supplément électronique
Remerciements
Les auteurs tiennent à remercier les deux correcteurs anonymes pour leurs commentaires pertinents.