

1 Introduction
Wolves (Canis lupus) are the first species known to have been domesticated during the Late Glacial by both European and Asian hunter-gatherers. This occurred somewhere around 17–15 kyrs BP [1–6] or perhaps earlier, around 20–30 kyrs BP [7]. However, this did not bring about major modifications in the way of life for humans, except perhaps small changes in hunting strategies, tactics or techniques. This suggests that at least during the Upper Palaeolithic Homo sapiens were already able to domesticate, but only utilized this behavior sporadically for a restricted number of species in particular circumstances. From ca. 12 kyrs BP several other species of animals and plants began to be domesticated in other parts of the world [8–11]. Though only a tiny fraction of all biodiversity has actually been domesticated [12], domestication of new species continued throughout the next millennia until the present day, where it is still active primarily with fish. Contrary to dog domestication, these domestications were part of a major change in the way of life of an increasing number of human societies throughout the world, in a process called Neolithisation. This process is not only characterized by a slow but drastic techno-economic shift from hunting-gathering to food production, based on cultivation and husbandry of domesticates, but also by a strong demographic transition [13] combined with deep social and spiritual change [14].
This paper aims to summarize the current state of knowledge accumulated by archaeology and archaeozoology during 50 years of studying Neolithic animal domestication. It will briefly present the archaeozoological methods, whilst attempting to emphasize the main trends of this phenomenon against two different scales, namely the continental and regional (Near East) scales to discuss the conceptual issues and the reasons behind the birth of domestication. Many bibliographic references are provided to help readers getting a deeper insight into this fascinating topic of domestication.
2 Archaeological approaches to early animal domestication: concepts and techniques
Archaeological evidence of domestication, such as representations of scenes of husbandry or remains of objects linked with husbandry (e.g. yokes, fessels) are rare and often ambiguous. Thus, the best way to investigate early domestication consists of studying archaeological skeletal remains (archaeozoology [15–19]). These remains provide substantial and important evidence that deserves attention:
- • if they come from well-dated and characterized archaeological contexts they can often be dated with relative precision (± some decades to 2–3 centuries) and as this date can normally be corroborated by direct radiocarbon dating of the collagen from the bones themselves it is therefore possible to analyze the domestication processes with high temporal resolution, even for early period's ca. 12-10 kyrs BP;
- • contrary to paleontological or even Pleistocene collections they often constitute a large series allowing quantitative approaches and statistical appreciation of the observations;
- • as Late Glacial and Holocene archaeological bones are not fossils, histological structures, associated unicellular or helminthic parasites and organic matters are often well preserved, allowing for a large panel of biological analyses, including paleomolecular or isotopic approaches [20];
- • in addition to multiscale and refined analyses of size and shape (e.g. using geometric morphometrics [21,22]), which provide information on size and shape evolutions and on age and sex at death, they constitute a large and very informative panel of pre-mortem (or intrinsic) biological signatures;
- • the archaeological context of discovery (food refuse deposits, human burial, cultural sites) as well as the post-mortem marks on the bones (cut marks, cooking burns) give indications as to the relationship of the species with humans.
Conversely archaeological approaches are limited by:
- • archaeozoological discoveries coming from limited regions or periods, being badly documented or not yet studied;
- • the low rhythm of archaeological analyses, often five to ten yearly excavation sessions are required before the refined chronological or contextual information is available, without which animal bones cannot be used;
- • the loss of most of the biological information with only the skeleton being preserved;
- • the extreme fragmentation of the bones, due to the systematic consumption of marrow and the post-depositional attrition of the collections, which reduces the quality and quantity of archaeozoological information.
Consequently, to fully analyze the preserved archaeological faunal collections, including paleomolecular or isotopic analyses, it is of utmost importance to have an in-depth knowledge of both the archaeological contexts and the taphonomic processes that have degraded the information [23]. This cannot be achieved without a tight and well-balanced collaboration between the excavator, whose scientific approach is as important and difficult as that of the analysts, the osteo-archaeologist, in charge of the general study of the faunal assemblages, and the specialists who undertake the molecular, geo-morphometric or isotopic investigations.
Studying early Neolithic domestication naturally requires a clear theoretical view of precisely what domestication is. Archaeologists generally agree that domestication can be defined as the process whereby the reproduction of a deme (i.e. local sub-population) of animals or plants is appropriated and controlled by human society for material, social or symbolic profit. Domestication, within this definition, is clearly differentiated from the pet-keeping of some Amazonian [24], New Guinean or Japanese Ainu hunters-gatherers, which consists of capturing a young wild animal, for a particular household rather than for a whole society, raising it (and even breast-feeding it) then later releasing or killing it without any offspring as a symbolic offering to nature to guarantee their future subsistence.
As a process dependent on the animal/plant species and on the multiplicity of human behavior domestication takes various forms. These can be arranged on a gradient of eco-anthropological mutualistic relationships between animal and human societies [25,26]; from anthropophily, to commensalisms or control in the wild, the management of captive animals, expansive or intensive breeding, and finally to pets (Fig. 1). As the process depends solely on the dynamic equilibrium between animals and humans, it is possible to achieve sustained stability at any level; with further progression or the retreat back to a less intensive relationship being possible. This is clearly illustrated by the diverse situations observed today (e.g. the reindeer [27]) and by the frequent phenomena of feralization [28–30]. The crucial question for archaeology, therefore, is to decide from which step of this gradient domestication began. It is clear that commensalisms and control in the wild are out of the frame of domestication, while control of captives can be considered as domestication (or not) according to the degree of animal reproductive control (Fig. 1). To draw a more precise line is unachievable, partly because of the continuity of the process but primarily because the delimitation of the concept of domestication depends solely on ethnocentric perception of the limits between nature and culture [27,31].

Domestication can be considered as an ultimate phase of intensification in the relationship between animal or plant sub-populations and human societies. It is comprised of several grades of intensification that, by various means and over varying periods, may end with the emergence of domestic animal as well as plant and microorganism lineages shaped by humans. The control of wild animals is a form of domestication that does not entail any visible morphological modifications, at least from an archaeology point of view.
With this in mind and using (and giving clear definition of) their own cultural concepts and words, a main concern of archaeologists is to accurately describe the characteristics of each of the particular situations, so that observed situations can be positioned in the gradient of Fig. 1. Though tightly connected with this process, the biological effects of domestication on plants/animals must be analyzed apart. The biological consequences of domestication vary according to the particular species. However, for vertebrates (including birds and fish) there are a number of constant modifications including a decrease in aggressiveness, increased fertility, sexual dimorphism (and often body size) decrease, shortening of the face, decrease of the braincase volume, appearance of new coat colors, soft ears (carnivores) and voice changing (barking of dogs) [12,16,32–35]. Some of these modifications result from hormonal changes, due to environmental conditions and the stress of captivity, i.e. without human intent [36,37] whereas others result from epigenetic/developmental changes or are purely genetic mutations possibly selected by humans. The long held belief of archaeo(zoo)logists is that these biological signatures of domestication appeared almost immediately after the breeding of captive animals began. It is now clear, however, that they may have appeared either fast, slow or never, according to the type of modification, the species and the intensity and nature of the relationship [38,39] (Fig. 1).
Using this theoretical framework it appears that the concept of (process of) domestication must be disassociated from that of the domestic animal. To recognize early domestications and early domestic animals archaeozoology has to combine the large panel of information of the archaeological skeletal remains to address the two connected, but mutually exclusive, questions:
- • the description of the level of domestication (i.e. intensity and nature of the relation);
- • the biological modifications due to domestication.
The former can be answered by analyzing:
- • the presence of animals in human burials [2,40];
- • the presence of a species out of its natural area of distribution [41–43];
- • the high frequency of the taxa in food refuse, which provides information about subsistence specialization;
- • the frequency of the skeletal parts, which often reveals the distance between the slaughtering and dwelling places [38,44];
- • and ‘paleodemographic’ data (i.e. a combination of the sex-ratio and frequency of ages at death).
The latter, which is also the most powerful method for addressing these questions [23,45], gives precise, though not always unambiguous, information about the strategy of animal acquisition by various practices of hunting or breeding. For example, J. Peters et al. [46] convincingly evidenced domestication from the drastic changes in slaughtering profiles of sheep and goats ca. 10.5 kyrs BP in the high Euphrates Basin. Using a different technique relying on recording ages and sexes proportions M. A. Zeder [47] also evidenced a strong intensification of the exploitation of the bezoar goat (Capra aegagrus) in the Zagros ca. 10 kyrs BP.
For archaeozoologists, addressing the biological modification question of early domesticates using size decrease [16,46,48] is a well-utilized domestication signal. However, as mentioned above, this modification sometimes occurred well after the initial domestication, if ever, depending on the intensity of the human/animal relationship. For example, J. Peters et al. [46] evidenced a very rapid size decrease in wild boar ca. 10.5 kyrs BP in the high Euphrates Valley, whilst A. Ervynck et al. [49] observed a very slow rate of decrease in the nearby high Tigris basin ca. 9 kyrs BP, the difference between these studies being probably due to different management systems. Another important issue of this technique for dimorphic ungulates such as cattle and goats is the fact that changes in the sex ratio can mimic a change (decrease or increase) in the average size. Based on both bone measurements and the state of long bone epiphysation, M. Zeder [47] proposed a technique that overrides this bias. Another promising and easier technique consists of a separate study of male and female sizes using mixture analyses [50–52]. It allowed detection of the earliest decrease in the sexual dimorphism of cattle, ca. 10.4 kyrs BP, in the Middle Euphrates Valley [51] (i.e. the earliest evidence of cattle domestication) though no decrease in the average size was detectable from the same bone sample.
3 Early animal domestication on a global scale
From the vast knowledge held, a brief statement about the world's earliest mammal domestications will allow us to draw out the main characteristics of this event at a global scale. Except for the dog, see Introduction, the earliest detected domestications are from the Near East. They concern the Oriental mouflon (Ovis orientalis), which gave birth to sheep (O. aries), bezoar goat, which is the ancestor of the domestic goat (Capra hircus), extinct aurochs (Bos primigenius), which generated domestic cattle (Bos taurus), and wild boar (Sus scrofa), the domestic form of which being the pig (Sus domesticus). All these domestications date approximately from the middle of the 11th millennium BP [39,45,46,48,51,52]. This includes the cat (Felis silvestris), an incipient domestication [40] that probably coincided with the start of early agricultural stock keeping and the subsequent development of commensal mice [53]. Goats, however, were also domesticated much later (ca. 10 kyrs BP) in the Zagros using various local different lineages [47]. Cattle domestication also started for a second time (ca. 8.5 kyrs BP) in the lower Indus Valley (Pakistan) with a different lineage, the humped Asian subspecies of aurochs (Bos p. namadicus) [54,55].
The existence of a third independent cradle of cattle domestication in the high Nile valley is still being debated [56–59]. Another independent center of pig domestication has been evidenced in China (ca. 8,000 BP [22,60]). Previous evidence suggested that only the pig (and probably the dog) had been locally domesticated here. However, investigations on the modern genetic lineages of wild and domestic Sus indicated several new potential centres of pig domestication in South East Asia [61], though currently there is no reliable archaeological evidence corroborating this statement. Local wild boar domestications in Europe have also been evidenced based on osteological and genetic data [62], ca. 7-5th millennia BP. During the same period (7th mill. BP) llama (Lama glama) and alpaca (L. pacos) were domesticated in Peru [63], from L. guanicoe and L. vicugna, which were later hybridized [64].
Another wave of large mammal domestications occurred in the Old World during the 6-4th millennia BP. Thus, horse (Equus caballus) was domesticated ca. 5.5 kyrs BP in Kazakhstan [65] and possibly also during the 5th millennium BP in the Iberian peninsula [66]. The origins of ass (Equus asinus) and camels (Camelus bactrianus, Camelus dromedaries) are still debated but it seems that they had not been domesticated before the 6th and 4th millennia BP, respectively. Finally the earliest evidence for the domestic water buffalo (Bubalus bubalus) has been found in the Indus Valley, dating to the 5th millennium PB [60,67]. The domestication of birds (turkey and Barbary duck in America, hen in South-East Asia) came later on.
Though brief, the present overview helps pointing out two of the major trends of early animal domestication. Firstly, it appears that very few of these domestication events were contemporaneous with the main climatic crisis, ca. 12,500 BP for the last cold phase of the Younger Dryas, ca. 11,500 BP for the increased Holocene warming and ca. 8500 BP for the most important (but short) Holocene cooling. Even though some processes of domestication appear to have been initiated during the Younger Dryas [43], all large mammal domestication took place during the Holocene, a period that provided more favorable climatic conditions and that could have, therefore, played an important role in the overall process. Secondly, even though some of these local domestication events could have been induced by contact with other areas where domestication had already been practiced it is clear that at least some domestications (e.g. pig in China, llama in America) began independently. This suggests that very different societies were initiating similar ideas in completely different parts of the world, not at precisely the same time, but most presumably under similar general climatic conditions, and during the same relatively short period of time.
An understanding of the potential reasons for this major and unique change requires examining these processes on a much smaller scale.
4 Early Neolithic domestications at a regional level: the Near East
Current knowledge states that the Near East formed the earliest cradle of Neolithisation [8,9,11–14,26,38]. In an area comprised of the Middle Euphrates Valley and South Palestine, a section of the Late Glacial population became sedentary (ca. 16–15 kyrs BP), a culture defined as the Natufian [14]. These societies appear to have lived in rather modest villages composed of semi-buried round houses and were still hunter-gatherers exploiting a broad spectrum of resources similar to their Paleolithic ancestors [68]. The introduction of the wild boar to Cyprus, before 11.4 kyrs BP [50], during the end of the Late Glacial (Fig. 2) indicates that these societies practiced a form of wild ungulate control that included a sophisticated form of hunting or incipient domestication. Shortly after the climatic Holocene transition (ca. 11.5 kyrs BP) these populations began to cultivate wild cereals and legumes [69,70] and gradually evolved toward a new culture defined as the Pre-Pottery Neolithic A (PPNA). Thus, PPNA villages were much larger and more sophisticated than the Natufian ones. In the Northern Levant, they were composed of standardized rectangular houses arranged centrally around a large, circular and semi-buried house probably used for common storage and for political/religious meetings [71]. A large PPNA religious site, such as Göbeckli in the high Euphrates basin, with its impressive megalithic architecture and animal rock art, testifies to this highly organized society [72]. Thus, these established and widely distributed PPN societies were responsible for the earliest ungulate domestications in the world.
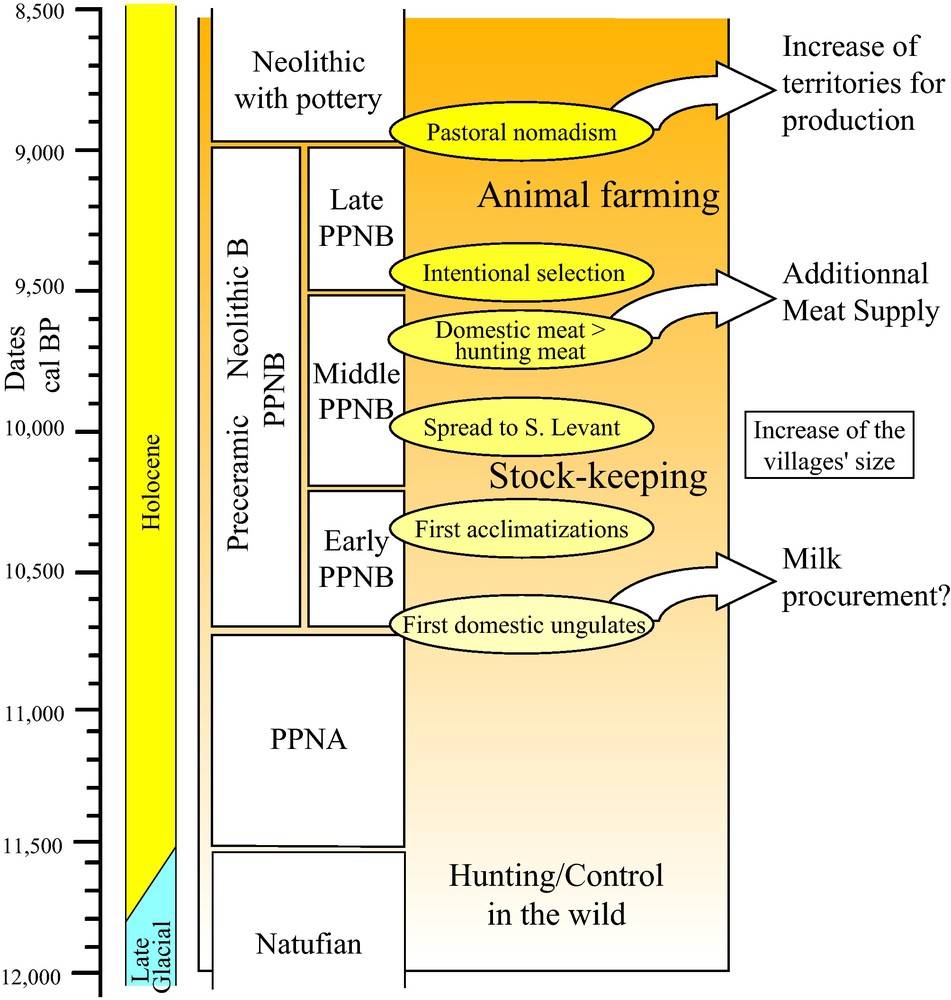
The different phases in the emergence of animal farming in the Near East [41].
As previously mentioned, the earliest known evidence for both new management strategies and morphological modifications (i.e. size/sexual dimorphism decrease) of ungulates dates from ca. 10.7 to 10.5 kyrs, in the high (sheep, goats, pigs [46]) and middle (cattle [51]) Euphrates basin on the high slopes of the Taurus Mountains. Approximately this coincides with the beginning of the early PPNB (Fig. 2) characterized by the cultivation of domestic cereals and legumes and by a newly sophisticated technique of producing long and rectilinear flint hunting weapons [14]. Shortly after, early domestic ungulates were transported far from their areas of origin toward the south, to the Damascus region [73], and even overseas to Cyprus where goats and cattle appeared (ca. 10.4–10.3 kyrs BP) followed by sheep, pigs and the Mesopotamian fallow deer (a failed attempt to domestication) ca. 10 kyrs BP [42,48,52].
These transportations necessarily entailed acclimatization of the early domesticates and brought about a further level of animal control [39]. Notably the earliest evidence of milk exploitation also dates to approximately this time [39,48,74,75] (Fig. 2). Around 10 kyrs BP, further to the east in the Zagros Mountains, people were still mobile hunter-gatherers (Zarzian culture [14]) but had possibly controlled wild goats for some time [76], being locally and probably independently domesticated from the Anatolian cradle [47]. During the same period, the earliest domestic ungulates appeared further to the east in Central Anatolia [77] and to the south in the Jordan valley and the Dead Sea area [78]. From ca. 10 kyrs BP, domestic sheep, goats, cattle and pigs were being bred across an expansive area stretching from Cyprus and Central Anatolia to the Iranian Plateau from west to east and from the high Euphrates and Tigris valleys to the southern borders of Palestine. This area was mainly composed of high mountainous slopes (Taurus, Zagros) and included some of the largest Near Eastern valleys (high and middle Tigris, Euphrates, and Jordan). It seems improper, therefore, to call this zone the Fertile Crescent (i.e. the society that appeared 4000 kyrs later) because its geographical entity was restricted to the large valleys that is, from the Nile to the Lower Euphrates-Tigris (Mesopotamia) [79]. More importantly it appears increasingly likely that animal (and plant) domestication did not occur solely in one place then diffuse outwards across a larger area, but that this domestication appeared simultaneously in numerous different and geographically distant places, evolving at different speed in each of them [80] and spreading outwards from each of these to other communities. The distance between these probably varied greatly and would have required a dense and complicated network of both short and long distance relationships. This period corresponding to the first half of the 10th millennium (Middle PPNB, Fig. 2) is characterized by a strong demographic increase in all the Levantine and Anatolian areas as attested by the drastic widening of the tell villages [14]. Nevertheless, during this time hunting of large game (animals hunted for food and not normally domesticated: hemions, gazelles, aurochs, bezoar goats, deer, ibex) still provided most of the meat consumed [39,75]. In this respect, the middle of the 10th millennium represents a new important turning point (Fig. 2). This was the moment (ca. 9.6–9.5 kyrs BP) when husbandry definitively became the primary source of meat and when important ungulate morphological modifications (namely horns) began to appear, suggesting the start of intentional selection and crossbreeding [81,36]. This was the birth of animal husbandry in the Near East. It was followed several centuries later by the start of pastoral nomadism [82], which appeared not as a primitive form of husbandry but as an innovative way of seasonal exploitation of the sub-desert areas, which even today is visible across vast areas of the Old World.
Nearly at the same time, the Neolithic new ways of life, including pottery and domestic plants and animals, began to spread to the Eastern Asia, to North-Eastern Africa and South-East Europe [83,84]. The scenario in the Near East, as with all other centres of Neolithisation, is that they cannot be generalized as a single event. Each must, and can, be defined separately by its own unique succession of events, be they with or without pottery, agriculture or sedentism. However, they all followed a slow and complex process, which involved small interconnected communities with demographic growth, in a zone where lived one or several animal/plant species able to be domesticated.
From this examination of early animal domestication at the global and regional scales we discuss below the conceptual framework for the study of the Neolithisation and its possible causes.
5 The conceptual framework for the study of early domestication
Neolithisation is characterized by the start of an unprecedented increase in human activity and its subsequent impact on the environment. It is marked by the birth of new types of ecosystems including those strongly impacted by humans (man-modified ecosystems, e.g. exploited forests) and those distinctly artificial (man-made ecosystems, e.g. agrosystem, village, city) [85,86]. This new ecological organization required the redeployment of plants and animals into new biotopes according to their specific capacity for adaptation to these new ecosystems and to the intentional human decision of rejecting undesirable species or taking advantage of their material, social or spiritual properties [86] (Fig. 3). Several authors analyzed this phenomenon in terms of “human niche” construction, i.e. “the process whereby organisms modify selective environments, thereby affecting evolution” [87–89]. Others analyzed the birth of domestication as a phenomenon of biological co-evolution [90]. Here, human niche construction would have favored biological changes in proto-domesticates as ensuring increased reproduction in a “disturbed” environment would make them more attractive as a human resource. In turn humans would have been selected (via cultural evolution) or motivated (via rational choice) to modify their behavior so as to favor the propagation and growth-multiplication of these proto-domesticates. However, as stressed by M. A. Zeder [10] “what makes human niche construction and the human/plant and animal mutualistic relationships involved in the process of domestication different from these classic biological processes is the element of human intent” (see also [27]). It is therefore necessary to consider Neolithisation, and more importantly plant and animal domestication, not as a simple additional episode in the biological evolution but as a discontinuity in the evolution of the biosphere. That is, one special species became able to modify its environment according to long term predictions, fed by a multi-generation memory of successes and failures and using socialized (i.e. flexible) techniques. In other words, the pure neo-Darwinian conceptual framework cannot account for the anthropological and biological complexities of the Neolithisation and/or the birth of domestication, notably concerning the techno-economic and symbolic uses of animals by human societies as being part of their technical, social and symbolic systems (sensu M. Mauss and C. Lévi-Strauss [91,92]) and subsequently being a characteristic part of their cultural system [93]. We prefer, therefore, to use the structuralistic conceptual framework of the anthroposystem, i.e. a meta-system constituted by the culture system (with its own anthropological functioning), the ecological system (with its own biological functioning), and their interactions and dynamics through time [15,85,86,94] ( Fig. 4). For the same reason we keep the term ‘co-evolution’ for true biological interactive evolution and use the term ‘co-development’ where human cultures are involved. For example, most of plant or animal domestications are co-developments because they involve human intent.
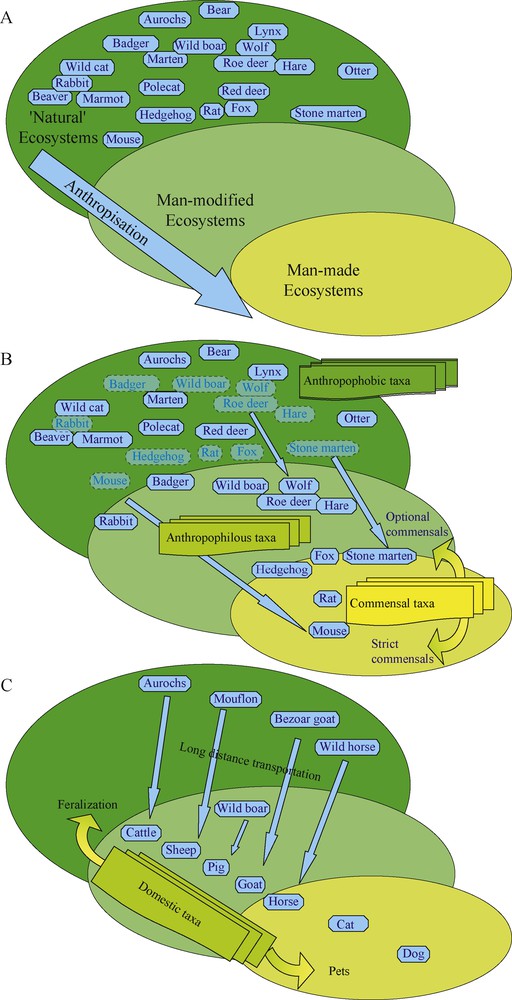
Schematic representation of the effects of anthropisation on the ecosystems of animal communities (example of the large mammals of Western Europe). A. Apparition of new man-modified and man-made ecosystems. B. Redistribution of taxa in this modified ecological structure according to their respective niche and apparition of a new ecological structuration of the communities into three new groups: anthropophobous, anthropophilous and commensal. C. Proposal of an ecological interpretation of domestication as a component of animal communities anthropisation, though with intent from humans [15].
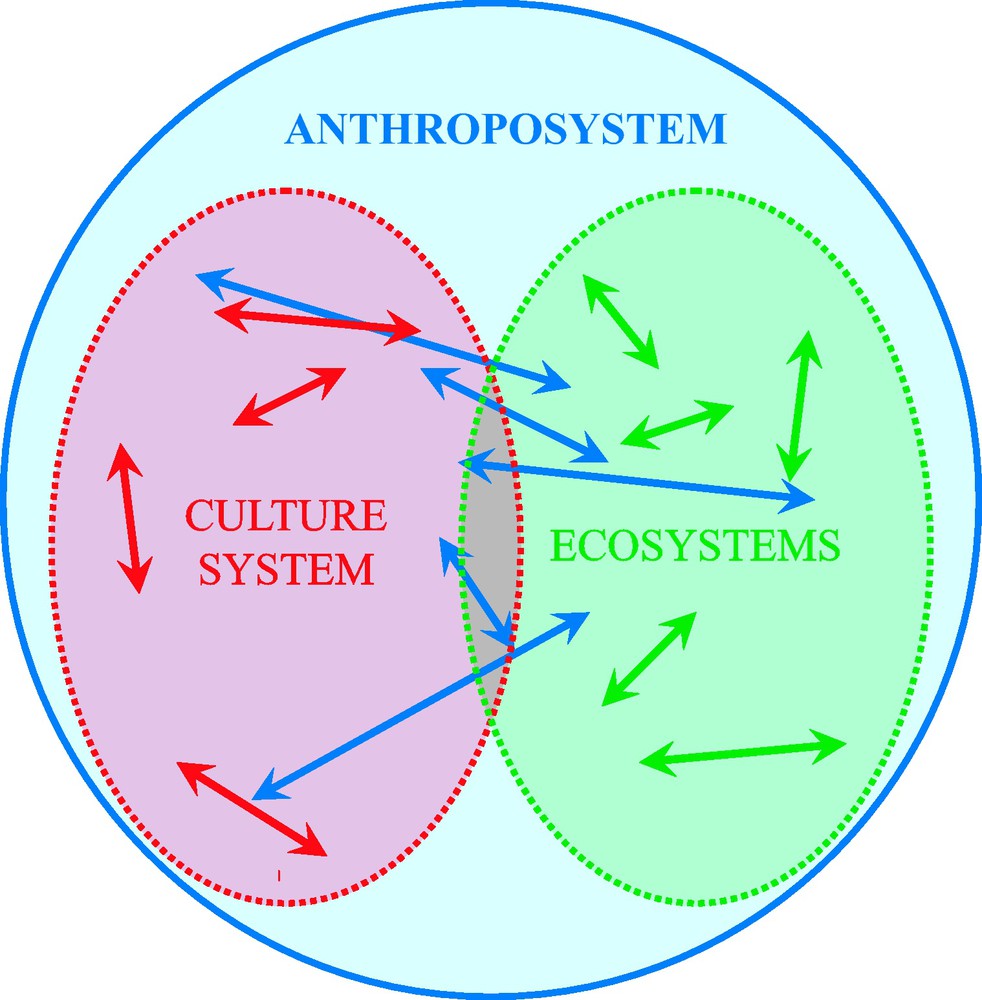
Schematic representation of the anthroposystem, a meta-system constituted by the cultural system (red) with the interactions between its proper components (red arrows), the ecosystems (green) in which the human society lives and the biological interactions between its components (green arrows), and the interactions between these two (groups of) systems (blue arrows).
6 The possible causes of Neolithisation and domestication
Neolithisation was not a revolution in the sense of an abrupt historical phenomenon. Across the Near East, it lasted at least 4–5 kyrs from the earliest evidence of wild animal control (ca. 14 kyrs) to the emergence of farming (ca. 9.5 kyrs). Thus from a global perspective it lasted more than 10 kyrs. However, as emphasized earlier, the Neolithisation was most certainly a revolution in the sense that it opened up an unprecedented era for the biosphere allowing creating numerous new regimes of functioning environments not solely ruled by the physical and biological factors but also by socio-cultural ones due to the acceleration of anthroposystems development.
Climate has long been considered the principle cause of Neolithisation, namely the cooling during the Younger Dryas, as it would have forced some human societies to diversify into new modes of subsistence [95–97]. However, as already stated, recent refinements to the chronology of climatic changes, at the end of the Late Glacial and during the early Holocene, disclosed the absence of strict correlations between them and the Neolithisation process. Therefore climate change cannot be considered to have been the principal instigator of Neolithisation [14,39,86,98], though the increasingly stable, and thus more predictable, warm Holocene climate did create more favorable conditions for human demographic increase and in turn for successful Neolithisation.
Although the demography of Homo sapiens continued to increase since the emergence of the species [13], it is clear that Neolithisation would not have been possible before a certain demographic density and therefore could not have occurred earlier in its history. Yet the fact that the demography suddenly and drastically increased just after the beginning of agriculture in numerous regions of the world (Europe, North America, Near East [13]) indicates that, as with climate change, demography alone cannot solely explain Neolithisation.
Sedentism is another factor that has long been considered as the first step, or as one of its omnipresent components, of Neolithisation. In the Near East it began during the Late Glacial and preceded the earliest domestications by at least 4000 yrs. In central Asia the societies who domesticated horses or camels several millennia later were clearly nomadic and stayed so. The last step in the birth of husbandry in the Near East is the return, by some sectors of the population, to a more pastoral nomadic way of life. This confirms that even though it constantly increased worldwide during the Holocene and undeniably provoked an increase in birth rate, hence a strong demographic increase [13], sedentism does not appear as a compulsory component of the Neolithisation. Also the fact that PPN people continued to obtain most of their meat diet from hunting during the thousand years after the appearance of domestic ungulates does not support the hypothesis that Neolithisation was a way to increase meat supply [39]. At the most, domesticates would have provided complementary resources during the less productive hunting seasons [39,86]. However, recent evidence of milk exploitation, from as early as the PPN in the Near East, suggests that the domestication of sheep, goats and cattle could have been stimulated by the quest for this new food [75]: it was only available from captive animals and its symbolic value as a figure of the maternal link should have already been high [99,100].
Therefore taking into account the anthropological, as well as the biological dimension, it appears that ownership of an animal, especially if it had a high symbolic value (such as wild boars or bovids, among the most frequent animal representations during the PPNA [72,101]) was a mark of social prestige, or even, as it is still the case in numerous present traditional societies, a sign of wealth. Prestige and wealth are of course hardly detectable through archaeological finds but we must keep in mind that there may be symbolic, as well as practical, reasons that stimulated the earliest appropriations of animals.
Aside from “pet keeping” [35], animal appropriation (i.e. domination) is almost impossible in animist or totemic societies as in their “horizontal”, cosmogonic conception of the world they consider animals, plants and humans as existing within the same hierarchical level [31]. For these societies, humans have no rights above those of other beings. In sharp contrast, to start the process of animal domestication implies that at some stage humans gave themselves permission to control nature. This suggests that to become dominant species humans had to radically change their “horizontal” conception of the world into a “vertical” one. In the Near East, this “verticalization” of the cosmogony is confirmed and strengthened by the archaeological observations of J. Cauvin [14]. This author evidenced that the Near Eastern PPN was characterized by the birth of divinities, with representations showing them standing above human figures or with humans praying toward the sky. This suggests that humans were beginning to see themselves within a hierarchical order dominated by divinities, placing themselves above animals and plants, which in turn allowed them to dominate or even kill them, while being pardoned of this unbearable act of murdering familiars “turning towards a higher entity, to whom [they] will […] offer the victim” [99].
This added dimension to Neolithisation highlights the importance of spiritual and symbolic human thought within the origins of animal domestication.
What is not yet completely clear is why, during the first half of the Holocene, so many distant human cultures throughout the Old and New World changed their beliefs, their techno-economic practices and their social organization. What seems clear is that Neolithisation was not a reaction to a single but rather numerous different factors (e.g., climate, biogeography, environment, demography, techno-economic practices, diet and health, social structure and mentality) simultaneously evolving at different degrees according to the local, regional or continental environment and scales [86], furthermore interacting and culminating in a snowball effect [9]. One possible scenario for the Near East would be that the demographic growth due to three millennia of sedentism combined with the more predictable Holocene climate would have increased both the hierarchy and the specialization of society, which in turn created the mental and social conditions necessary for animal appropriation and that these appropriations were then further stimulated by the social prestige of animal ownership, by the quest for new foods as milk, and/or by the buffering effect of domestic animals for smoothing of the seasonal irregularities of subsistence. In turn it is possible that these boosted demographic growth (e.g. at the beginning of the Middle PPNB) leading to the final adoption of husbandry as the main socio-economic mode of animal resources procurement. However, such a scenario must be stepwise tested with more archaeological evidence. Also more research outside of the Near East should be done, in order that scenarios for other parts of the world can be proposed and to stimulate comparative studies and a common model.
The lasts 60 years of archaeological studies showed that Neolithisation was not just another step in the history of humanity and the biosphere but corresponded to a pivotal change in their historical trajectories. It is then of utmost importance to increase the archaeological information held to better understand the factors playing major roles during each step of the Neolithisation process, not only for the benefit of archaeological researches, but to help answering the many questions facing our present society on the sustainability of its own future.
Conflicts of interest statement
The author has not declared any conflict of interest.