

1 Introduction
Much of our present knowledge on the ecology and behaviour of animals is derived from longitudinal studies of individuals. Long-term datasets are essential to study life-history traits (e.g. age-specific survival, age at first reproduction, reproductive lifetime) [1], and to understand how variations in those traits might impact population dynamics and behavioural strategies. However, the collection of such datasets requires the ability to identify individuals repeatedly over time, i.e. by individual markings. For most bird species, monitoring is possible by means of ringing, a method by which observers are able to read an individual identification number on a leg ring, either at distance using binoculars or a telescope, or by recapturing birds (particularly in the case of smaller sized species). For instance, in the current context of increasing anthropogenic pressures, capture-mark-recapture data obtained from banding have enabled researchers to model how seabird populations may be impacted by fisheries and climatic variations through changes in breeding success and survival [2–5]. However, leg rings are not suitable for all bird species. For instance, due to anatomical peculiarities of their leg joint, penguins cannot be banded with traditional leg rings. Nonetheless, obtaining longitudinal data from penguin populations is crucial, as some species are endangered and others live in extreme habitats where climate changes and their impact on marine prey are predicted to be important. They are thus particularly relevant biological models for investigating changes in life-history traits and population dynamics, and may act as bellwethers of climate change [2] and marine ecosystem health (Fig. 1).
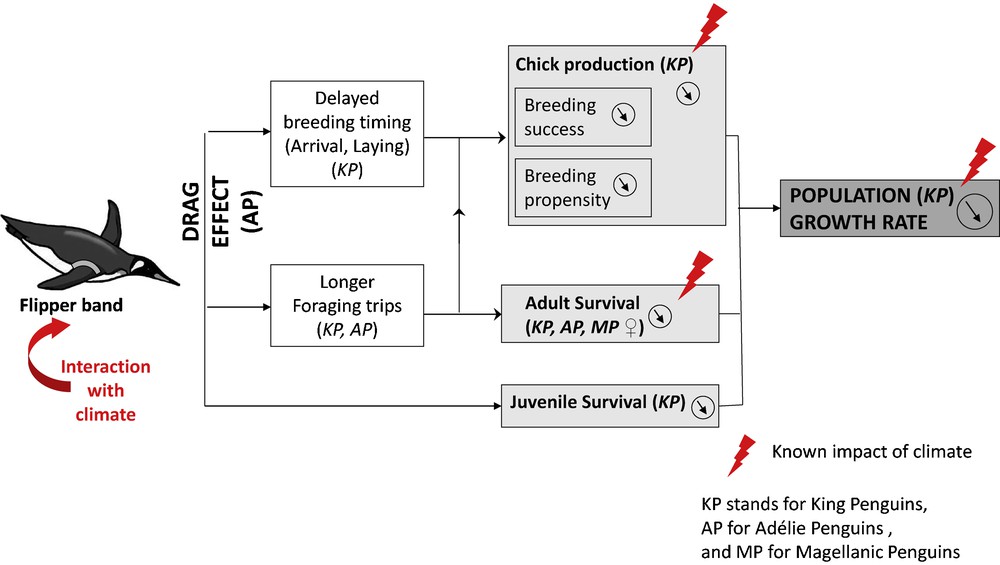
Effects of flipper bands on penguin populations and their interaction with climate. The small black arrows into circles indicate where flipper bands induce a reduction, e.g. such as a reduction in breeding success.
Consequently, since the 1950s, flipper bands have been widely used [2,6,7] to study the life-history traits and behaviour of various penguin species but also to track individuals used in physiological investigations. Data from flipper-banding have also been used to explain observed changes in penguin populations in relation to climate changes [8,9], ultimately predicting the future impact of climate on these populations [10]. However, since the 1970s, detrimental effects of flipper-banding have been reported (for review, see [11,12]). In an early reaction to this, at the end of the 1980s, the observation of tissue injuries induced by flipper bands resulted in the cessation of many penguin flipper-banding projects as a matter of precaution. However, the harmlessness of flipper bands remains a controversial issue and continues to be subject to much debate and dispute. Flipper-banding is still being used in research and conservation projects, such as for investigating the survival of penguins rehabilitated after an oil spill [13]. Some teams, who are still using large scale banding schemes, claim that flipper bands have no significant effects, at least in some species [14,15].
In this context, one may question the reliability of data based on flipper-banding if it has an impact on the life-history traits of penguins. Moreover, “it raises practical and larger ethical questions about costs and benefits of procedures in field studies” [16]. For those teams that have abandoned banding, can data previously obtained from banded birds be used without discussing the various implications of flipper-banding impacts? Is available information sufficient to accurately correct for banding effects?
In the context of global change, obtaining data on penguin biology and ecology is unquestionably necessary for scientists to address future threats. Here, we review the pros and cons of the three penguin monitoring methods known in use today. More specifically, we compare flipper bands to Radio Frequency IDentification (RFID) tags, which appear to be the main alternative to flipper bands since their introduction in 1991 in king penguins, Aptenodytes patagonicus [17]. We will also discuss the advantages and disadvantages of web tags, i.e. small-animal identification tags which are attached to the outer webbing of penguin feet [15].
2 Advantages of flipper bands
2.1 Avoidance of capture stress
Flipper-banding usually requires a single initial capture to fit the band. Since the band number can be read from a distance with binoculars, recapture is not needed for further identification, except [15] when bands become loose and need to be securely reattached. Flipper bands therefore enable the monitoring of penguins throughout their whole life with a single capture, thus avoiding the further stress associated with recaptures, such as in many other bird species (i.e. for which recapture is necessary in order to read tag numbers on smaller scaled rings).
RFID tags share this advantage with flipper bands since identification may be automatically performed using fixed antennas on the typical passageways of the birds. The main limitation of RFID however, is the short reading distance of antennas which is required to identify the tag, i.e. with a maximum of about 1 m for the 31 mm transponders used in the Texas Instruments Radio Frequency Identification TIRIS system (see [18]). Thus, to settle such systems requires the existence of bottleneck locations, where birds will always pass when travelling between the colony and the sea. Further, to identify penguins in the colony, a portable hand reader is needed and the birds must to be approached very closely by observers with the subsequent consequences of human disturbance.
The use of web tags definitely requires both an initial capture of the birds and subsequent recapturing to be able to read the small markings of the tags [15]. Thus, it incurs an even higher disturbance cost to birds and is therefore not favoured as a method for marking individuals.
2.2 An easy and cheap means to identify penguins
Flipper bands, either in plastic or metal, are easily visible for an observer, at least when both flippers of the penguin are visible. Moreover, when the carcass of a banded penguin is found, removing the band or recording the number is simple. Thus, even if the person finding the carcass is not the scientist involved in penguin monitoring, it is likely that the band or the number will be forwarded (the correspondence address is usually indicated on the band itself). In contrast, RFID tags can only be read with dedicated equipment, therefore precluding any identification when such equipment is not available. Moreover, they are not visible and hard to detect as they are implanted under the skin and are small (they weigh less than a gram).
In addition, the cost of flipper bands is extremely low (less than 1 € a piece), as are the small ear tags (0.1 € a piece) designed for rodents that have been adapted for the use on penguins [15]. On the other hand, whereas RFID tags are also affordable (around 2–3 €), the cost of a hand RFID reader is much more expensive (about 500 €) and the cost of a fixed set-up with series of antennas, readers and computers, and spare equipment (such as used for the king penguin colony on Possession Island in Crozet archipelago [18]) can reach 20 to 30 k€, depending on the number of passageways that need to be equipped. Moreover, those figures include spare equipment, but do not account for accessory costs for sheltering the equipment and for maintenance. Yet, to ensure its continuous functioning in the field, the full time presence of an engineer is compulsory, or there is a higher risk of gaps in data collection due to equipment failure. But note that a RFID system generates a huge number of data even if there are some interruptions. For an identification set-up to run at a remote field site, solar cell systems and/or batteries are needed and can also require daily maintenance (with in some cases the added problem of poor light, such as in Antarctica during the winter). Note, however, that the reading of a band number at a distance also requires people to be continuously in the field. But, in contrast to an automatic system, human based informations do not allow the same constant recapture effort and individual birds can be missed.
3 Disadvantages of flipper bands
Hereafter, we will only consider the effects of single flipper bands compared to those of RFID and web tags, as most banding schemes usually involve monitoring penguins with a single flipper band. The main effects of flipper bands are summarized in Fig. 1.
3.1 Tissue injuries
The moulting process causes penguin flippers to swell [19] and, particularly at that time, flipper bands, which can easily be fitted too tightly or not properly secured (with a slight opening), may induce severe injuries to flipper tissues [15,20]. Wound depth in flipper joints may be 1 cm or more, thus damaging tendons and muscles. In extreme cases, presumably due to infections, this can lead to death [21]. The effects of flipper bands, however, depend both on band material and shape; some bands are worse than others. For instance, aluminium bands are known to be more harmful than stainless-steel bands, as they can deform easily, especially under the high pressures encountered while diving [15]. No infections have yet been reported with transponder injection or web tag piercing.
3.2 Increased drag
Culik et al. [22] found in 1993 a 24% increase in the energetic cost of subsurface swimming in Adélie penguins Pygoscelis adeliae in a water canal. This was an important finding, because it revealed the importance of the increased drag resulting from flipper bands. The authors estimated that the increased cost would reduce by half the prey captured by free-ranging banded penguins.
The drag increase associated with any external device fitted to a penguin, depends on the shape and material of the device [23]. Accordingly, Barham et al. [24] found that stainless-steel flipper bands caused more drag on swimming African penguins Spheniscus demersus than experimental silicone rubber bands. RFID tags, since they are implanted under penguin's skin, avoid any possible drag effect. The same presumably applies to web tags since penguins use their flippers instead of their feet to propel themselves when they swim, and web tags are located on the feet positioned within the continuity of a streamlined body. It can therefore be assumed that they cause little if any drag, and thus their energetic cost to a penguin is negligible [15].
3.3 Lengthening of foraging trips
If flipper-banded birds are not able to compensate for the handicap resulting from increased drag, longer foraging trips are to be expected as a consequence of lower swimming and foraging efficiency compared to non-banded birds. By monitoring breeding male and female Adélie penguins with subcutaneous RFID tags over four years (about half of which were banded with stainless-steel flipper bands), Dugger et al. [25] found an 8% significant increase (3.5 hours) in the duration of the foraging trips of banded birds relative to their non-banded conspecifics. Investigating 60 breeding male Magellanic penguins Spheniscus magellanicus over a 13-month period (half of the birds had either stainless-steel flipper bands or a web tag), Boersma and Rebstock [14] found an eight-hour difference in foraging trip length between banded and non-banded birds. However, the difference (almost 20%) was not significant, possibly due to large variability in the data (standard deviation of 13 to 21 hours for an average duration between 43 and 52 hours). Saraux et al. [26], who investigated over 10 years 100 RFID tagged king penguins of which half had a stainless-steel flipper band, found that banded birds made significantly longer trips at sea during all incubation and brooding shifts (that is when both mates alternate on the egg and chick, respectively) than non-banded ones (i.e. of 12.7 versus 11.6 days). The mean increase in the duration of foraging trips was 9% [26], and therefore similar to that observed for banded Adélie penguins [25]. Importantly, the effect was still observed after a decade [26], thus arguing against the assumption advanced in some studies [8] that penguins are ultimately able to adapt to their band.
3.4 Reduced breeding success
The significant impact of flipper-banding on breeding success was first shown in 2004 in a five-year study on king penguins [27], already indicating that the effect of flipper bands on breeding success does not disappear after one or two years [27]. Saraux et al. [26] recently demonstrated that banded birds, even after a 10-year period from initial banding, still exhibited a reduced breeding success compared to non-banded birds. Altogether, chick production over ten years was nearly twice as large for non-banded birds as for banded king penguins: the 50 non-banded king penguins produced 80 chicks whereas the 49 banded birds produced only 47. What are the drivers of such a poor breeding success? We found that banded king penguins arrived later at the colony to breed [26], laid their egg later in the season, and in accordance with previous studies [7,27–29], that delaying reproduction onset resulted in lower breeding success [26]. The delay at the onset of breeding can be attributed to a lower efficiency in storing body fuels. Moreover, the proportion of returning birds engaged in breeding was lower for banded than for non-banded birds, suggesting that those non-breeding banded birds may have been unable to store sufficient body fuels. Note that a bird with a delayed start in breeding also has reduced choice in mate selection.
3.5 Reduced survival in adults
In their study, Dugger et al. [25] found that adult survival was 11–13% lower in flipper-banded Adélie penguins over the 2002–2003 season. Over a longer period, they however observed a high variability in survival, including years of high survival for banded birds, which could be associated primarily with variable environmental conditions. Their data showed that previous studies [30] had underestimated bands’ effect, which likely occurred throughout the lifetime of the animal.
These effects of flipper bands on penguin survival were investigated in more detail in king penguins [26]. Over a decade, adult birds marked with stainless-steel bands had an average survival of 20% compared to 36% for non-banded birds. Plotting residuals of the model suggested that differences in survival increased up to 54 ± 3 months, i.e. 4.5 years. Those birds (banded and non-banded) which died during that period had a lower breeding success than the birds which survived, therefore suggesting that the lower survival in banded birds concerned poorer performers [26].
3.6 Reduced survival of juveniles
The survival rate of non-banded juvenile king penguins is about 75–80% after 2–3 winters [27], which is approximately twice as large as that reported in previous studies of flipper-banded chicks (47% in Brodin et al. [29] and 6–39% in Weimerskirch et al. [7]).
4 Scientific bias due to flipper bands
The impact of flipper bands on the survival and breeding success of penguins obviously introduces a bias in those studies where the changes in these traits are used as indicators of the impact of climate change on populations. The real question concerning flipper-band data is thus whether data obtained from flipper-banded birds may be corrected.
4.1 Uncertainty in the rate of band loss
As reported by Boersma and Rebstock [15], the rate of band loss can vary between 5 and 22% for aluminium bands, which open more easily, compared to a rate of loss of only 0–2% for stainless-steel bands. Moreover, the method used to determine the rate of band loss, i.e. by tracking double flipper-banded birds to calculate the proportion of those losing one band [8], is biased. Indeed, the survival of doubly banded birds is lower than that of single banded birds [31].
In contrast, the rate of RFID tag loss is negligible for 31 mm tags if inserted properly under the skin, i.e. in a way preventing their ejection after initial insertion [26]. However, Boersma and Rebstock [14] found a tag loss of 2.6% for 12 mm RFID tags injected under the skin of penguins’ feet. The small size and the location of these tags may explain their high rate of loss. Indeed, such small tags have a very short reading distance of about 10–20 cm and cannot be implanted under the better-protected abdominal skin if they are to be read by underground antennas. In the feet, transponders may be less likely to get trapped in subcutaneous blubber, and thus are subjected to higher physical stress and ejection as penguins walk. It is therefore preferable to use tags with greater detection distances. Concerning web tags, they may pull out, therefore leaving a small tear in the webbing [15].
4.2 Scientific bias
Since flipper bands can affect adult survival and/or reproduction, any study using raw (uncorrected) data collected through flipper-banding schemes takes the risk of confounding banding effects with the investigated climate or anthropogenic effects, thus leading to incorrect inferences.
Correcting data by accounting for banding effects therefore seems the obvious solution. However, the major problem with such an approach is that flipper-banded penguins are differently affected by climate when compared to non-banded birds. In banded Adélie penguins for instance, the increase in foraging duration varies according to the year [25], whereas African penguins only seem to be negatively affected by banding during periods of reduced prey availability [32]. In king penguins [26], the population growth rates of banded and non-banded birds did not respond similarly to variations in sea surface temperature. Indeed, differences between the two groups were most apparent in “intermediate” years. Food availability at sea could be so poor for a given year that even those birds not handicapped by a band might fail in large numbers. In contrast, in years of very favourable environmental conditions, the environmental pressure on banded king penguins is weak and the extra cost inflicted by banding may then be less visible, explaining the absence of or slight difference observed between banded and non-banded birds in cold and favourable years [26].
A major difficulty also arises for studies investigating the impact of climate change on penguin dispersal. Indeed, a drop in breeding success increases dispersal in seabirds [33]. Thus, the reduced breeding success of banded birds may result in increased dispersal, resulting in an additional bias in ecological investigations. Presently, penguin dispersal is almost exclusively studied based on banded birds because one cannot surround all existing colonies with RFID antennas. However, the significance of such data is therefore put into question, as dispersal based on banded birds may not be representative of actual population dispersal in free-ranging penguins.
5 Conclusions
The long-term impact of flipper bands on the survival of juvenile penguins, and on the breeding success, survival and duration of foraging trips of adult penguins, as well as their relationships with climate, are only known in detail for king penguins. The observed impacts can be essentially attributed to the drag effect of the bands, and possibly also to flipper injuries, since, in accordance with data for Adélie penguins [25], they are associated with longer foraging trips [26], and most likely greater energy expenditure. This result is also in accordance with the pioneering studies of Wilson et al. that highlighted increased metabolic rates for banded swimming penguins, which suggested decreases in foraging efficiency [11,22,23]. In contrast to previous assumptions [8], the impact of flipper bands (at least for king penguins) is not limited in time. Birds which survived beyond the five first years following banding continued to display longer foraging trips and reduced breeding success [26]. Banded birds therefore seem unable to compensate for the handicap resulting from the increased band-induced drag, except when marine resources are unusually abundant [26]. Delays and/or failure/inability to engage in breeding altogether indicate that the main effect of flipper-banding is through a serious weakening of an individual body condition.
Due to the large size of the section of the colony that was surrounded by RFID antennas, we never observed two flipper-banded birds as a mated pair. In a smaller population where the chance of banded pairs would be higher, the effect on breeding would then presumably be even higher. Still, the observed impact certainly cannot be neglected since, for king penguins, the survival of banded chicks after 2–3 winters is approximately half that of non-banded chicks; the survival of banded adults over 10 years is of 20% instead of 36% for non-banded birds and their breeding success is about 40% lower than that of non-banded birds [26,27]. Moreover, it should be emphasized that the results presented were obtained using stainless-steel flipper bands, which are known to be much less harmful than the aluminium bands [17] still widely used in population studies of penguins. We may therefore assume that the bias on data obtained with aluminium bands [7–10] on previous investigations was far greater. However, since this impact has not yet been measured, any correction for the effect of banding that can be made for existing data in population dynamics investigations would be highly speculative. The study of king penguins [26] can at least be used as a benchmark for sensitivity analyses.
An important question is whether the impact of flipper bands is the same in different penguin species. In other words, can the data for king penguins be generalized to other penguins and, particularly to smaller penguins, since the king penguin is the second largest species after the emperor penguin Aptenodytes forsteri?
Since the drag increases as a square of the speed and the power that is required to overcome drag increases as a cube of the speed, the effect is expected to be smaller in smaller penguins if they swim at lower speeds than king penguins. But there is no clear relationship between size and swimming speed in penguins [34], and thus there is no reason for drag effects to be disproportionally higher in the large king penguins. The drag effect might even be higher for smaller penguins because their flipper bands are proportionally larger than for penguins of greater size. Yet, as indicated above, the measured increase in swimming metabolic rate induced by flipper bands in the small Adélie penguin is as much as 24% [22]. A specific problem for the emperor penguin is that it is the only bird to breed during the severe Antarctic winter, and we do not know the effect of temperatures well below freezing on wet flipper bands when the penguins jump out from water, or the impact of the bands during blizzards, i.e. while the birds are standing on sea ice.
It is sometimes argued that using flipper bands is the only way to investigate the dispersal of penguins. However, breeding success is a main trigger of dispersal [33], and its drop induced by flipper bands introduces a bias that raises questions about its costs and benefits. As pointed out by Robert May [16], there are therefore good reasons why we need to think more carefully about some present practices in field investigations.
Altogether, taking into account the important and long-term impact of stainless-steel flipper bands on juvenile and adult penguins, the finding that aluminium bands are even more harmful [15], and considering the uncertainties about the rate of band loss and the dispersal of banded birds, the continuation of banding schemes should be seriously reconsidered. We cannot be sure that there is no effect of RFID tags in penguins [12] but this is very unlikely since no negative effects have been found either in juveniles or adults in tits [35]. The much larger survival and breeding success of RFID tagged penguins is anyway a strong argument for the use of RFID as an alternative to flipper bands and for developing technical innovations to counteract the limitations of RFID. Compared to flipper-banding, another clear advantage of an automated RFID identification based on antennas surrounding a colony is that it enables an identification of penguins day and night at the right time they get in or out this colony. The number of individuals which can be monitored is also quite unlimited, therefore enabling investigations at a real population scale without a huge human involvement. Note that there is also a fully-automated population monitoring of penguins through a computer vision system that is using the natural markings in the chest plumage of African penguins [36]. However, such non-invasive system cannot be used on most species of penguins because they do not have such natural markings.
The data on the breeding success and survival of penguins based on banded birds, and particularly aluminium-banded birds [7–10], should also be reconsidered. Climate change is clearly having negative impacts on some penguin populations [37,38], but developing reliable forecasts requires unbiased estimates of the relationships between climatic variables and penguin demography.
Disclosure of interest
The authors declare that they have no conflicts of interest concerning this article.