

Abridged English version
The study of the relationships between plants supposes that we consider the plants as indicators of environmental parameters. This concept is widely accepted by botanists, ecologists and phytosociologists. The criterion of fidelity, introduced by the fathers of phytosociology, is applied in this paper in order to characterize the relationships between plants. The numerical application of the criterion of fidelity across France, by using a database containing 215,000 phytosociological surveys, gives an original result concerning the ecological characterization of phytosociological surveys: the “probable flora”, which represents the probability for a plant to find, in a given locality and with other plants having a close ecological behavior, the type of environment that allows its survival.
The calculation of the probable flora includes three steps: the first one is used to determine the apparent dependency between plants by using a calibration of taxa with the use of the criterion of fidelity. In this way, each taxon is ecologically situated close to the other taxa. The second step is to determine, for a given survey, which plants are the most ecologically discriminant, that is to say, those that explain the environmental factors of the phytosociological survey. Among the discriminant plants of the survey, there are some that have not been observed in it: they are considered as “probable flora”. The third part concerns the mapping of the probable flora in order to identify the ecological gradients.
The application of the calculation of the probable flora is considered, at the scale of France, by the use of the botanical and ecological database SOPHY. Currently, the database contains more than 215,000 phytosociological surveys, which represents more than four million elementary observations made by more than one thousand botanists. The used taxa represent a number of 8000 herbaceous and woody species, taking into account their levels of abundance/dominance, divided into six classes.
The authors present three examples of plant distributions that have a very distinct ecological behavior: a Mediterranean plant (Pinus halepensis Mill.), an Atlantic bioclimate plant (Ulex europaeus L.), and a mountain plant [Eritrichium nanum (All.) Schrader].
The distribution of fidelities of Pinus halepensis Mill. shows the Mediterranean behavior of this taxon that is concentrated near the littoral of the Mediterranean Sea and it has few occurrences outside of this area. In Corsica, where this species would have been introduced by humans, its probabilities of occurrence show that it should colonize the littoral and the hills of this island.
Ulex europaeus L. is a taxon that grows in acidic and deep soils, relatively humid in sub-Atlantic bioclimate. Its probability of occurrence in France clearly shows the sub-Atlantic character of this species and, at the same time, the localities with favorable environmental requirements for Ulex europaeus L. Its spatial distribution underlines that the averages fidelities of Ulex europaeus L. are higher on the front of the Atlantic coast than of the Channel coast. It also shows two main areas of high probabilities: one in Brittany and Loire-Atlantique and another in Aquitaine and the Basque Country.
Eritrichium nanum (All.) Schrader is an endemic species of the Alps and the Caucasus. Its probable distribution confirms the mountain behavior of this taxon and it also shows that it could grow in the Pyrenees mountains because of its ecological behavior. The probability of Eritrichium nanum (All.) Schrader in the Pyrenees is related to its high fidelity to Saxifraga aspera L. and Festuca halleri (All.) that are commonly found in the Alps and whose geographical distribution extends to the Pyrenees.
The mapping of probable flora highlights some more pronounced ecological gradients instead of the observed flora due to the intermittent nature of botanical observations. The probable flora reduces, in a certain way, the intermittence of botanical observations that may result of phenomena that are independent of the ecology of plants. Therefore, its use can be considered to improve the methods of characterizing bio-indicators, and to help the decision makers and the public utilities in their activities of territorial management, because determination of probable plants can participate in defining the vegetation dynamics in the short and medium terms.
1 Introduction
L’étude des relations entre plantes a conduit les botanistes, les phytosociologues et les écologues à considérer les plantes comme des indices de variables environnementales [1]. Tout botaniste de terrain s’attend à rencontrer telle ou telle plante selon le milieu qu’il prospecte, en compagnie des autres plantes qu’il rencontre. Cette supposition est liée à l’expérience du botaniste qui, au fur et à mesure des prospections qu’il réalise, rencontre des taxons botaniques dans différentes situations écologiques. Plus il rencontrera fréquemment un taxon dans un milieu donné et en compagnie d’autres plantes de comportement écologique semblable, plus il s’attendra à le retrouver dans de nouvelles localités de cortège floristique semblable et de conditions environnementales proches. Cette démarche empirique a été traduite numériquement comme étant la notion de fidélité entre plantes et milieu [2].
Originellement, la fidélité a été utilisée par les phytosociologues dans le but de rattacher les taxons aux groupements phytosociologiques, afin de dégager les plantes caractéristiques des différents niveaux hiérarchiques phytosociologiques, comme l’association. Avec l’arrivée des centres de calculs ont été développées les méthodes numériques fondées sur des critères de ressemblance de flore entre relevés [3]. Il faudra attendre les travaux de Brisse et al. [4,5] pour que cette notion soit utilisée pour définir le comportement écologique des espèces et pour caractériser écologiquement les milieux.
Le concept de fidélité des plantes aux plantes appliqué aux relevés permet de calculer les plantes discriminantes pour un relevé donné. L’intérêt de ce procédé est d’estimer, pour chaque relevé, les facteurs du milieu dont ils représentent un échantillon et dont les plantes représentent les témoins. Parmi les indices de variables du milieu, il existe de nombreuses plantes à forte fidélité moyenne vis-à-vis du relevé, mais qui n’ont pas été observées dans ce dernier. Ces taxons indicateurs portent le nom de plantes probables et constituent la « flore probable » des relevés de la banque SOPHY. La flore probable représente la probabilité de trouver une plante dans un lieu donné en compagnie d’autres plantes de comportement écologique proche [4].
Le but de cet article est de présenter l’apport du concept de « flore probable » en écologie végétale, à la fois pour la caractérisation des milieux et pour la mise en évidence de gradients écologiques traduits par la probabilité de rencontrer un taxon dans une localité ayant des facteurs environnementaux qui lui sont favorables.
2 Principe et application à l’échelle de la France
L’application du calcul de la flore probable est envisagée à l’échelle de la France par l’utilisation de la banque de données botaniques et écologiques SOPHY [6,7]. Actuellement, cette banque de données regroupe plus de 215 000 relevés phytosociologiques (Fig. 1), ce qui représente plus de quatre millions d’observations élémentaires réalisées par plus d’un millier de botanistes. Les taxons utilisés sont au nombre de 8000 espèces herbacées et ligneuses, en prenant en compte leur niveau d’abondance/dominance réparti en six classes (Fig. 1).

Carte de répartition des relevés phytosociologiques en France et dans les pays limitrophes.
La détermination de la « flore probable » comprend trois étapes :
- • la première détermine les liens de dépendance apparents entre plantes, en procédant à un étalonnage entre taxons par le calcul de leur fidélité sur l’ensemble des relevés de la banque SOPHY. De cette manière, chaque taxon est écologiquement situé par rapport à tous les autres taxons de la banque SOPHY ;
- • la deuxième consiste à déterminer, pour un relevé donné, quelles sont les plantes qui lui sont le plus écologiquement discriminantes, c’est-à-dire celles qui permettent d’expliquer les facteurs écologiques dont le relevé phytosociologique est un représentant. Parmi les plantes discriminantes du relevé, apparaissent un certain nombre de plantes qui n’ont pas été observées : elles constituent la flore probable du relevé ;
- • la troisième concerne la cartographie proprement dite de la flore probable.
2.1 Calcul de la fidélité des plantes aux plantes
Soient deux taxons notés respectivement T1 et T2, la fidélité notée FID (T1/T2) de T1 vis-à-vis de T2 s’exprime comme suit : [1,2] (Fig. 2)
| (1) |
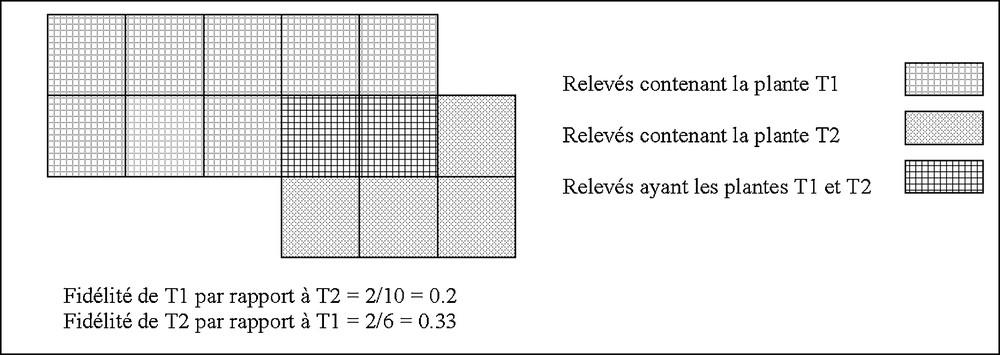
Calcul de la fidélité deux plantes T1 et T2.
Freq(T1,T2) = nombre de relevés ayant simultanément les plantes T1 et T2 dans la banque SOPHY.
FreqT1 = fréquence de la plante T1 dans la banque SOPHY.
Comme ce calcul est appliqué aux taxons de la banque SOPHY, un taxon possède des valeurs de fidélité avec les 8000 autres taxons de la banque. En regardant attentivement le calcul de la fidélité entre taxons, la fidélité du taxon T1 vis-à-vis du taxon T2 correspond à la probabilité de rencontrer T1 en présence de T2, et inversement. Cette probabilité rejoint les hypothèses empiriques avancées par les botanistes sur le terrain concernant la probabilité de rencontre d’un taxon dans un milieu donné. L’application de ce calcul donne un tableau de 8000 lignes sur 8000 colonnes où, pour chaque taxon étudié, une valeur de fidélité à l’égard de 8000 autres taxons est donnée. Ce calcul permet ainsi de considérer les plantes comme des indices de variables (IV) du milieu.
2.2 Caractérisation écologique des relevés
Si l’on considère les plantes comme des indices de variables environnementales, un relevé phytosociologique apparaît comme un échantillon de milieu donné par la liste d’indices du milieu que sont les « plantes discriminantes ».
La caractérisation écologique d’un relevé consiste à déterminer par le calcul les plantes discriminantes, c’est-à-dire celles qui vont participer le plus à l’originalité écologique du relevé. Le relevé est situé dans l’espace des fidélités des plantes de la banque SOPHY. Trois principales étapes permettent de caractériser un relevé par ses plantes discriminantes.
2.2.1 Calcul du centre de gravité de la banque SOPHY
Le calcul du centre de gravité de la banque a pour objectif de fournir un référentiel pour situer les plantes dans l’espace des fidélités. Ce centre de gravité résume l’information écologique contenue dans la banque SOPHY.
| (2) |
FID(T,IV) = fidélité de chaque taxon T à l’égard de tous le indices de variables IV.
FreqTot(T) = fréquence totale de chaque taxon T dans la banque SOPHY.
On obtient un vecteur Vsophy contenant 8000 valeurs de fidélités moyennes pour la banque.
2.2.2 Calcul du centre de gravité du relevé
Pour situer le relevé par rapport à l’ensemble des situations écologiques contenues dans la banque SOPHY, le calcul consiste à déterminer le centre de gravité du relevé à l’aide de la formule suivante :
| (3) |
NbObs le nombre d’observations de plantes dans le relevé.
Ce calcul donne le vecteur VRelevé contenant 8000 valeurs de fidélités moyennes pour le relevé.
2.2.3 Calcul du pouvoir discriminant des plantes
Ce calcul consiste à identifier les plantes écologiquement discriminantes du relevé, c’est-à-dire celles qui représentent des IV du milieu. Dans un premier temps, le calcul consiste à mesurer la distance entre le centre de gravité de la banque, qui est un référentiel, et celui du relevé.
| (4) |
Puis, le pouvoir discriminant du relevé est calculé par la formule suivante :
| (5) |
Ce calcul est appliqué aux 8000 IV de la banque. Le résultat du calcul du pouvoir discriminant DISCR est un dernier vecteur de 8000 IV, rangés selon leurs valeurs de pouvoir discriminant. Comme le relevé est caractérisé par l’ensemble des indices de la banque, il est indispensable de choisir ceux qui participent le plus à décrire l’originalité du milieu qu’il représente. Les indices sélectionnés pour expliquer le milieu du relevé sont ceux qui participent à expliquer la moitié de la distance entre le centre de gravité du relevé et celui de la banque.
Les plantes présentes du relevé ont une fidélité maximale à l’égard de ce dernier, ce qui représente, en fait, un artéfact, car elles font double emploi. Pour savoir si elles aussi sont discriminantes du milieu qu’elles sont censées représenter, le calcul élimine les indices dont la fidélité est égale à 1, car il s’agit des plantes fidèles à elles-mêmes. De cette façon, seuls les indices réellement discriminants vis-à-vis du milieu du relevé sont représentés. En général, les plantes discriminantes ainsi sélectionnées représentent un effectif compris entre 30 et 60 indices de variables et comprennent à la fois des plantes observées dans le relevés et des plantes ayant une probabilité d’occurrence dans le relevé (Tableau 1).
Comparaison entre la composition floristique d’un relevé et ses plantes discriminantes. Relevé effectué par A. Gaston en 1962 dans la montagne de la Clape, Aude, France. La première colonne correspond au code du taxon observé dans le relevé, la deuxième au nom latin du taxon observé, la troisième au niveau d’abondance de ce dernier, qui s’échelonne de 1 à 6. Les colonnes suivantes correspondent quant à elles au code de l’indice, au nom latin de l’indice et à sa fidélité à l’égard du relevé. En gras figurent les plantes probables du relevé.
| id | Nom latin des taxons observés | Abondance | id | Nom latin des indices de variables | Fidélités |
| 7795 | BRACHYPODIUM RAMOSUM (L.) R. & S. | 6 | 3402 | THYMUS VULGARIS L. | 0.59366653 |
| 9034 | PINUS HALEPENSIS MILL. | 4 | 5623 | RUBIA PEREGRINA L. | 0.49403283 |
| 3283 | ROSMARINUS OFFICINALIS L. | 4 | 7795 | BRACHYPODIUM RAMOSUM (L.) R. & S. | 0.46102807 |
| 1914 | STAEHELINA DUBIA L. | 4 | 3360 | TEUCRIUM CHAMAEDRYS L. | 0.45495004 |
| 7456 | CAREX HALLERIANA ASSO | 3 | 8452 | APHYLLANTHES MONSPELIENSIS L. | 0.4094805 |
| 2845 | QUERCUS COCCIFERA L. | 3 | 9034 | PINUS HALEPENSIS MILL. | 0.38873154 |
| 5375 | POTENTILLA VERNA L. | 3 | 2847 | QUERCUS ILEX L. | 0.38080251 |
| 5623 | RUBIA PEREGRINA L. | 2 | 7758 | AVENA BROMOIDES GOUAN | 0.37452588 |
| 1019 | FUMANA CORIDIFOLIA (VILL.) P. F. | 2 | 4205 | ONONIS MINUTISSIMA L. | 0.36838808 |
| 4205 | ONONIS MINUTISSIMA L. | 2 | 3283 | ROSMARINUS OFFICINALIS L. | 0.36739924 |
| 8453 | ASPARAGUS ACUTIFOLIUS L. | 2 | 9010 | JUNIPERUS OXYCEDRUS L. | 0.36358206 |
| 9013 | JUNIPERUS PHOENICEA L. | 2 | 7456 | CAREX HALLERIANA ASSO | 0.35346618 |
| 3360 | TEUCRIUM CHAMAEDRYS L. | 1 | 7872 | DACTYLIS GLOMERATA L. | 0.34906434 |
| 1002 | CISTUS MONSPELIENSIS L. | 1 | 1019 | FUMANA CORIDIFOLIA (VILL.) P. F. | 0.33734803 |
| 997 | CISTUS ALBIDUS L. | 1 | 8453 | ASPARAGUS ACUTIFOLIUS L. | 0.32947073 |
| 3225 | LAVANDULA LATIFOLIA (L.) VILL. | 1 | 3965 | DORYCNIUM SUFFRUTICOSUM VILL. | 0.32539503 |
| 3279 | PHLOMIS LYCHNITIS L. | 1 | 6428 | ERYNGIUM CAMPESTRE L. | 0.31482244 |
| 1717 | LEUZEA CONIFERA (L.) DC. | 1 | 7790 | SUBSP. PHOENICOIDES R. & S. | 0.29798528 |
| 3370 | TEUCRIUM POLIUM (L.) RY | 1 | 8813 | SMILAX ASPERA L. | 0.28843654 |
| 3402 | THYMUS VULGARIS L. | 1 | 3633 | PHILLYREA ANGUSTIFOLIA L. | 0.28576829 |
| 3633 | PHILLYREA ANGUSTIFOLIA L. | 1 | 7810 | BROMUS ERECTUS HUDS. | 0.28428441 |
| 3984 | GENISTA SCORPIUS (L.) LAM. | 1 | 1584 | HELICHRYSUM STAECHAS (L.) DC. | 0.27129514 |
| 437 | LONICERA IMPLEXA AITON | 1 | 2848 | QUERCUS LANUGINOSA LAM. | 0.26907918 |
| 4944 | CLEMATIS FLAMMULA L. | 1 | 5172 | RHAMNUS ALATERNUS L. | 0.26713033 |
| 6428 | ERYNGIUM CAMPESTRE L. | 1 | 4476 | PISTACIA LENTISCUS L. | 0.2648843 |
| 6586 | SESELI MONTANUM L. | 1 | 997 | CISTUS ALBIDUS L. | 0.26473602 |
| 7758 | AVENA BROMOIDES GOUAN | 1 | 437 | LONICERA IMPLEXA AITON | 0.25701117 |
| 8452 | APHYLLANTHES MONSPELIENSIS L. | 1 | 1628 | HIERACIUM PILOSELLA L. | 0.25536045 |
| 9010 | JUNIPERUS OXYCEDRUS L. | 1 | 4944 | CLEMATIS FLAMMULA L. | 0.24990898 |
| 4633 | POLYGALA RUPESTRIS POURRET | 1 | 5529 | ASPERULA CYNANCHICA (BAUHIN) L. | 0.23650879 |
| 3917 | CORONILLA MINIMA L. | 0.22230804 | |||
| 7925 | FESTUCA DURIUSCULA L. | 0.22162997 | |||
| 1914 | STAEHELINA DUBIA L. | 0.21629782 | |||
| 5567 | SUBSP. CORRUDAEFOLIUM VILL. | 0.20630064 | |||
| 3902 | CALYCOTOME SPINOSA (L.) LINK | 0.20164433 |
Comme le montre le Tableau 1, parmi les plantes discriminantes sont présentes des plantes qui n’ont pas été observées par le botaniste. Ces plantes, dites probables, sont considérées comme des témoins de variables environnementales. Leur affectation à ce relevé montre dans quelle mesure le milieu représenté par ce relevé peut leur être favorable.
2.3 Expression cartographique de la flore probable
La cartographie de la flore probable représente les fidélités moyennes des plantes discriminantes, c’est-à-dire des indices de variables du milieu aux relevés. Cependant, pour atténuer le sur-échantillonnage de taxons dans certaines régions mieux explorées que d’autres, on comptabilise les coexistences des indices dans un quadrat d’1 km2. Ce principe présente l’avantage de montrer d’une façon plus originale la répartition probable des milieux favorables à un taxon. L’ensemble des 215 000 relevés phytosociologiques localisés représente 30 000 quadrats. La flore probable, ainsi cartographiée, permet de définir des gradients écologiques de flore sur le territoire français.
3 Résultats : identification de gradients écologiques de flore
Le concept de fidélité des plantes aux plantes appliqué aux relevés permet de calculer les plantes discriminantes pour un relevé donné. L’intérêt de ce procédé est d’estimer, pour chaque relevé, les facteurs du milieu dont ils représentent un échantillon et dont les plantes représentent les témoins. Parmi les IV du milieu, il existe de nombreuses plantes à forte fidélité moyenne vis-à-vis du relevé mais qui n’ont pas été observées dans ce dernier. Ces taxons indicateurs portent le nom de plantes probables et constituent la « flore probable » des relevés de la banque SOPHY. La flore probable représente la probabilité de trouver une plante dans un lieu donné en compagnie d’autres plantes de comportement écologique proche. La cartographie des fidélités de plantes aux milieux montre les gradients écologiques de flore de façon plus accentuée qu’en utilisant la flore observée. Les exemples suivants montrent la comparaison entre les répartitions observées et probables de trois taxons de comportement écologique distincts : Pinus halepensis Mill. (Fig. 3), Ulex europaeus L. (Fig. 4) et Eritrichium nanum (All.) Schrader (Fig. 5).
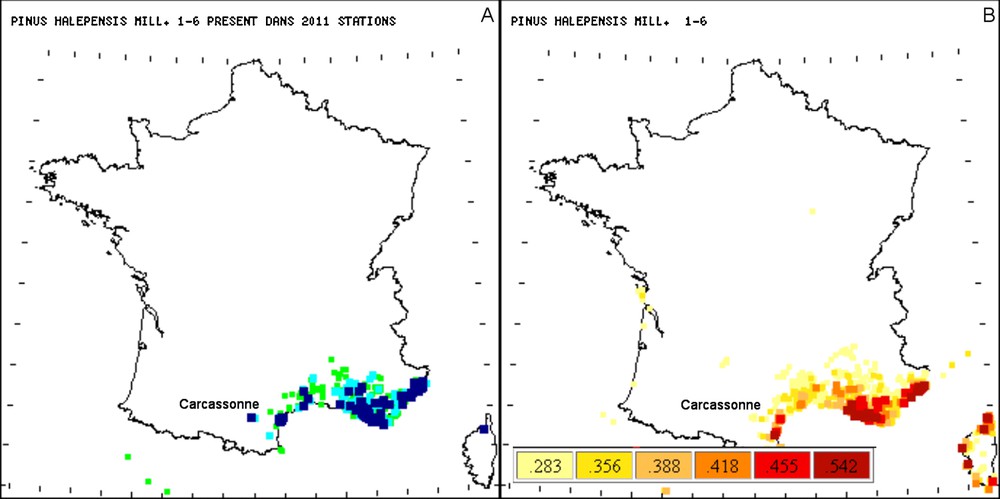
Comparaison entre les répartitions observées (A) et probables (B) de Pinus halepensis Mill. Les gradients d’intensité des couleurs correspondent pour les observations botaniques (vert et bleu) aux classes d’abondance croissantes 1–2, 3–4 et 5–6. Pour les fidélités moyennes aux relevés (jaune, orange et rouge), les variations de couleurs correspondent aux gradients croissant de fidélités. Pour l’interprétation des couleurs, le lecteur est prié de se référer à la version en ligne.
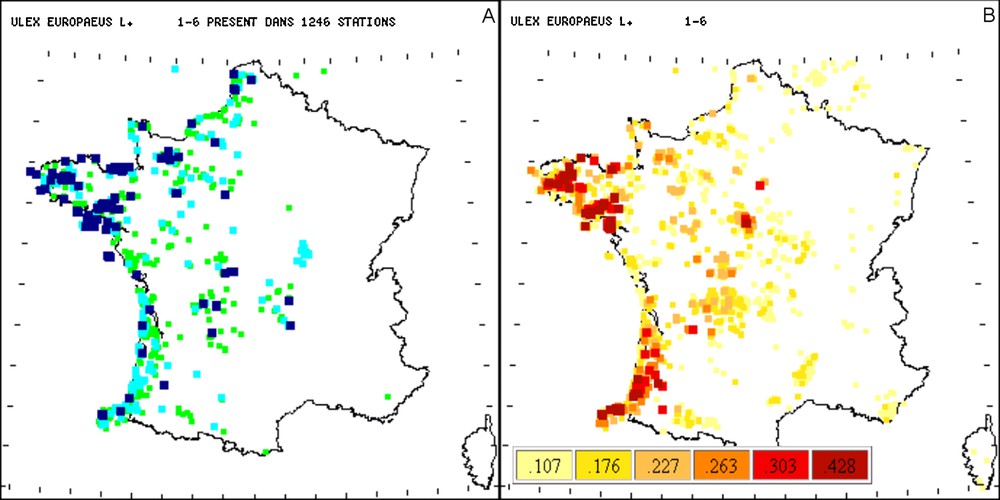
Comparaison entre les répartitions observées (A) et probables (B) d’Ulex europaeus L. Pour l’interprétation des couleurs, le lecteur est prié de se référer à la version en ligne.
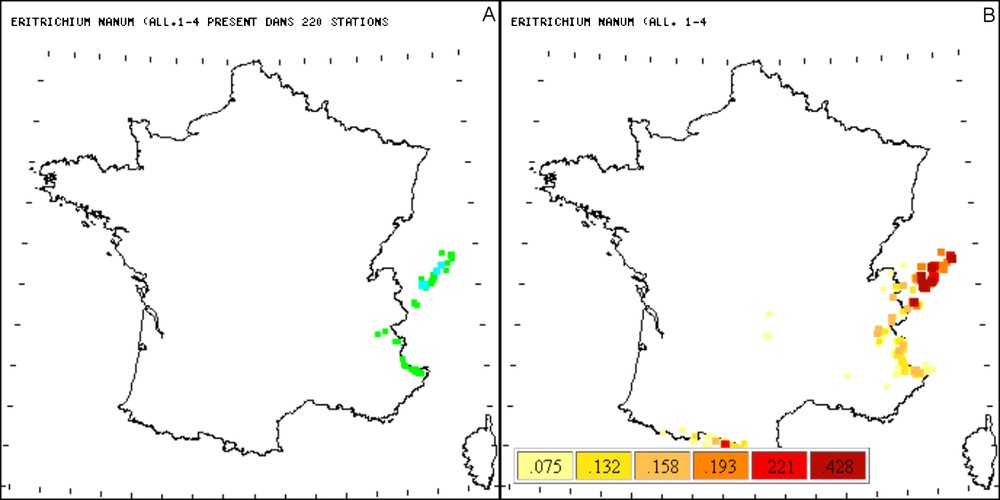
Comparaison entre les répartitions observées (A) et probables (B) d’Eritrichium nanum (All.) Schrader. Pour l’interprétation des couleurs, le lecteur est prié de se référer à la version en ligne.
La répartition observée montre les occurrences du taxon sur le terrain dans les localités prospectées par le botaniste. La flore probable témoigne des fidélités moyennes du taxon considéré comme un indice de variable du milieu. Cette distinction en tant qu’indice du milieu n’est possible qu’à partir du moment où l’on a mesuré le lien de dépendance apparent entre Pinus halepensis Mill. et tous les taxons de la banque SOPHY. La carte de répartition des fidélités moyennes du pin d’Alep montre que ce taxon est bien méditerranéen et qu’il est très peu présent au-delà du littoral méditerranéen (Fig. 3a). En Corse, où ce taxon a vraisemblablement été introduit [8], ses probabilités d’occurrence (Fig. 3b) montrent qu’il pourrait coloniser des localités situées plus au sud, où des conditions écologiques lui seraient favorables. La comparaison des répartitions observées et probables montre que, parfois, le taxon, présent ou bien abondant, est observé dans des localités où sa probabilité d’occurrence reste faible. Ce point n’entre pas en contradiction avec la réalité, mais montre simplement que, même si le taxon est abondant dans la localité, il ne présente peut-être pas forcément un grand avenir et pourrait être remplacé par d’autres taxons dont le milieu leur convient encore plus. Cet aspect peut dans certain cas rendre compte de la dynamique du milieu à moyen terme. En effet, le pin d’Alep est considéré comme un taxon pionnier appartenant au modèle expansionniste [9] et pouvant donc être remplacé par des essences plus stables dynamiquement parlant. Par exemple, Pinus halepensis Mill. abondant a été observé dans la région de Carcassonne (Fig. 3a), mais ses fidélités moyennes aux relevés de cette région restent relativement faibles. En revanche, les valeurs de fidélité moyenne du pin d’Alep sont importantes sur le littoral et à sa périphérie. Elles montrent ainsi l’aspect thermoméditerranéen de ce taxon, sans prendre en considération des observations qui peuvent résulter de facteurs contingents comme la pression anthropique.
Ulex europaeus L. est un taxon qui croît dans des localités à sol acide assez profond, relativement humide et de bioclimat subatlantique [10]. Ses probabilités d’occurrence en France montrent bien le caractère atlantique à subatlantique de ce taxon et, par la même occasion, les stations ayant des facteurs environnementaux proches des exigences d’Ulex europaeus L. Notons aussi (Fig. 4a) que les fidélités moyennes d’Ulex europaeus L. sont plus fortes sur la façade atlantique que sur la façade de la Manche et montrent deux principales zones de probabilités : une en Bretagne et en Loire-Atlantique et une autre en Aquitaine et Pays basque.
Eritrichium nanum (All ;) Schrader est une espèce endémique des Alpes et du Caucase [11]. Sa répartition probable confirme le caractère montagnard de ce taxon et montre aussi qu’il pourrait croître dans les Pyrénées en raison de son comportement écologique. La probabilité d’occurrence d’Eritrichium nanum (All ;) Schrader dans les Pyrénées est liée à sa forte fidélité avec des taxons comme Saxifraga aspera L. et Festuca halleri All. qu’il rencontre fréquemment dans les Alpes, et dont la répartition géographique s’étend jusqu’aux Pyrénées.
4 Discussion
4.1 Intérêt de la flore probable en géographie botanique
La répartition géographique d’un taxon est le plus souvent discontinue, en raison de son comportement écologique, mais aussi en raison de facteurs contingents autres comme la pression anthropique. Il en résulte qu’un taxon ne colonise jamais toutes les localités que son comportement écologique pourrait lui permettre. Cependant, les écologues s’attendent souvent à trouver tel ou tel taxon dans une formation végétale donnée. Par exemple, lorsque Paul Fournier [12] décrit Oreochloa disticha (Wulf.) Link. il précise que cette espèce, fréquente dans les Pyrénées et très rare dans les Alpes, est « à rechercher dans nos Alpes, parmi Carex curvula et Festuca varia ». En observant la répartition d’Oreochloa disticha (Wulf.) Link. (Fig. 6a) d’après la banque SOPHY, on remarque que ce taxon est bien représenté dans les Pyrénées et très peu observé dans les Alpes. L’utilisation du critère de fidélité des plantes aux relevés montre en revanche que ce taxon pourrait être plus fréquent dans les Alpes (Fig. 6b) en raison de ses co-occurrences avec des taxons comme Chrysanthemum alpinum L., Gentiana alpina Vill. ou Carex curvula All., eux-mêmes considérés comme étant des taxons très fidèles à Oreochloa disticha (Wulf.) Link. Par la même occasion, comme Fournier précise qu’Oreochloa disticha (Wulf.) Link. se rencontre dans les alpages et les rocailles sur substrat granitiques, la répartition probable de ce taxon montre alors des gradients des types de milieu dans lesquels Oreochloa disticha (Wulf.) Link. pourrait être rencontré. Notons que ces gradients sont nettement plus visibles en utilisant la flore probable plutôt que la flore observée. Aussi, la flore probable représente une solution originale pour ceux qui souhaitent chercher des localités abritant un taxon donné ou bien un type de milieu particulier.

Comparaison entre les répartitions observées (A) et probables (B) d’Oreochloa disticha (Wulf.) Link. Pour l’interprétation des couleurs, le lecteur est prié de se référer à la version en ligne.
Par ailleurs, Festusca varia (Haencke) Hack., considérée comme écologiquement très proche d’Oreochloa disticha (Wulf.) Link. selon Fournier, ne fait pas partie des plantes les plus fidèles de ce taxon, qui en a rencontré 387 autres dans 197 relevés de la banque SOPHY. Ce résultat souligne l’intérêt d’utiliser une banque de données phytosociologiques pour calculer avec précision les fidélités des plantes entre elles et entre les relevés pour caractériser, de la meilleure façon possible, le comportement écologique des taxons et mettre en évidence des gradients écologiques de flore en observant uniquement les fréquences des plantes, indépendamment de tout préjugé empirique. La flore probable permet dans ce cas de rendre plus continu le gradient écologique montré par un taxon que ne le font ses observations directes.
4.2 Intérêt de la flore probable pour caractériser écologiquement les milieux en France
La probabilité d’occurrence des plantes probables représente ainsi un apport d’information pour caractériser le milieu du relevé de manière encore plus précise que les plantes observées. En effet, lors du travail de terrain, le botaniste observe le plus grand nombre possible de taxons. Cependant certains peuvent avoir échappé à sa vigilance ou bien ne se trouvent pas dans la localité prospectée pour des raisons anthropiques ou bien liées à l’histoire de leur répartition. Bien souvent, le botaniste s’attend même à trouver des plantes dans les milieux qu’il connaît bien et qui parfois sont absentes. C’est pourquoi les occurrences probables des indices de variables du milieu apportent un complément d’information quant à la caractérisation écologique du relevé.
En 1953, Gaussen [13] fait remarquer que certaines plantes caractéristiques d’associations végétales peuvent être absentes de l’association, ce qu’il traduit comme étant la « hêtraie sans hêtre ». Dans cet article, l’auteur montre que l’attribution d’Asperula odorata L. en tant que plante caractéristique du Fagetum silvaticae est ambiguë, car cette espèce peut aussi coloniser des formations à Picea excelsa (Lam.) Lk., desquelles Fagus silvatica L. est pourtant absent. En appliquant l’étalonnage socioécologique de la flore, les résultats montrent, au contraire, qu’Asperula odorata L. figure parmi les plantes les plus écologiquement proches de Fagus silvatica L. (Tableau 2), ce qui conforte le choix des phytosociologues dans l’attribution de ce taxon comme indicateur de milieu à Fagus silvatica L., même si ce dernier en est absent [14].
Liste des 30 plantes les plus écologiquement similaires à Fagus silvatica L. La première colonne (PDCUM) correspond au pouvoir discriminant cumulé de la plante vis-à-vis de Fagus silvatica L. ; la deuxième colonne présente le nom de plante discriminante ; la troisième colonne (Fid) correspond à la fidélité et la dernière colonne (Frq) à la fréquence de la plante discriminante dans les quadrats.
| PDCUM | Fid | Frq | ||
| 14 | FAGUS SILVATICA L. | 1–6 | 1.00 | 7393 |
| 16 | CORYLUS AVELLANA L. | 1–6 | .45 | 7884 |
| 18 | HEDERA HELIX L. | 1–6 | .44 | 8924 |
| FAGUS SILVATICA L. | 5–6 | |||
| 22 | QUERCUS SESSILIFLORA SA | 1–6 | .35 | 4568 |
| 24 | DESCHAMPSIA FLEXUOSA (L.) | 1–6 | .36 | 7034 |
| 25 | VIOLA SILVESTRIS (LAM.) | 1–6 | .35 | 5268 |
| 27 | OXALIS ACETOSELLA L. | 1–6 | .31 | 3707 |
| 28 | POLYSTICHUM FILIX-MAS | 1–6 | .31 | 4214 |
| 29 | ASPERULA ODORATA L. | 1–6 | .30 | 2898 |
| 31 | ILEX AQUIFOLIUM L. | 1–6 | .29 | 3802 |
| 32 | LAMIUM GALEOBDOLON (L.) | 1–6 | .29 | 3898 |
| 33 | PTERIDIUM AQUILINUM (L.) | 1–6 | .30 | 6358 |
| 34 | ANEMONE NEMOROSA L. | 1–6 | .29 | 4010 |
| 35 | ABIES ALBA MILL. | 1–6 | .27 | 3099 |
| 36 | ATHYRIUM FILIX-FEMINA | 1–6 | .27 | 3774 |
| 37 | QUERCUS PEDUNCULATA EHR | 1–6 | .29 | 6336 |
| 38 | VACCINIUM MYRTILLUS L. | 1–6 | .27 | 4514 |
| 39 | CARPINUS BETULUS L. | 1–6 | .27 | 4099 |
| 40 | SORBUS AUCUPARIA L. | 1–6 | .26 | 3815 |
| 41 | FRAXINUS EXCELSIOR L. | 1–6 | .28 | 5486 |
| 42 | LONICERA PERICLYMENUM L. | 1–6 | .26 | 4857 |
| 44 | SOLIDAGO VIRGA-AUREA L. | 1–5 | .32 | 3917 |
| 44 | PRENANTHES PURPUREA L. | 1–6 | .26 | 5170 |
| 45 | FRAGARIA VESCA L. | 1–6 | .24 | 2616 |
| 46 | MILIUM EFFUSUM L. | 1–6 | .26 | 5034 |
| 47 | ACER PSEUDOPLATANUS L. | 1–6 | .23 | 2753 |
| 48 | EUPHORBIA AMYGDALOIDES | 1–6 | .30 | 3335 |
| 49 | LONICERA XYLOSTEUM L. | 1–6 | .23 | 3335 |
| 49 | CAREX SILVATICA HUDS. | 1–6 | .22 | 3392 |
| 50 | SORBUS ARIA (L.) CRANTZ | 1–6 | .22 | 3549 |
Les plantes les plus fidèles à un relevé représentent, d’une certaine façon, l’équivalent des plantes caractéristiques des phytosociologues. Ces plantes sont aussi des indices de facteurs du milieu et permettent de cartographier les milieux les plus propices à un taxon donné. Ces indices montrent alors des gradients écologiques de flore en raison de la répartition des valeurs de probabilité. Appliquée à un échantillonnage réparti sur un territoire comme la France et avec un lot important de relevés phytosociologiques, la flore probable constitue un moyen dans la compréhension des liens entre les plantes et leur milieu.
Enfin, le concept de « flore probable » se démarque aussi de celui de « végétation potentielle » habituellement défini par les phytosociologues, les écologues et les biogéographes, car il ne repose pas sur une démarche empirique issue des approches synchroniques ou diachroniques d’analyse de la végétation [15,16], mais, au contraire, intègre avant tout traitement numérique des relevés phytosociologiques l’application du critère de fidélité des plantes aux plantes à partir d’un vaste lot d’observations recueillies au sein d’une banque de données. L’application du critère de fidélité des plantes aux plantes à l’ensemble des observations de la banque de données permet alors de connaître numériquement la dépendance apparente d’un taxon vis-à-vis d’un autre, ce qui n’est pas le cas avec la démarche de détermination de la « végétation potentielle ».
5 Conclusion
La flore probable, résultat de l’application du concept de fidélité des plantes aux plantes énoncé par les pères de la phytosociologie, est un moyen simple et précis pour rendre compte des liens entre flore et milieu. Sa cartographie permet d’identifier des gradients écologiques de flore à l’échelle du territoire français et permet de retourner à la géographie des plantes avec un regard plus écologique. Elle réduit, dans une certaine mesure, les intermittences des taxons qui peuvent résulter de phénomènes indépendants de leur écologie et, par conséquent, son utilisation peut être envisagée pour améliorer les méthodes de caractérisation de bio-indicateurs, ou encore pour aider les botanistes dans leurs prospections floristiques et les décideurs dans le cadre de l’aménagement du territoire, car les plantes probables peuvent fournir des indications sur la dynamique de végétation à court et moyen termes.
Déclaration d’intérêts
Les auteurs n’ont pas transmis de déclaration de conflits d’intérêts.