

1 Introduction
The Mediterranean basin hosts about 12 endemic vascular plants per 100 km2 and has been recognized as a priority region for conservation in Europe, as well as one of the 34 major “biodiversity hotspots” on the planet [1,2]. Besides its thousands of species of flora, the Mediterranean region is home to some 455 million humans, who depend heavily on its natural resources, especially water. Historical and present-day human disturbance is ubiquitous, leading to degradation and physical modification of riparian landscapes [3] at accelerating speed and on an increasingly large scale. Riparian landscapes are currently under increasing pressure to supply superficial and groundwater for irrigation of intensive crops and for tourism in coastal areas [4]. The rapid loss and conversion of floodplains and threats to remaining areas make it urgent to improve our understanding of their internal dynamics in order to optimise conservation of what remains. Rapid methods of assessing plant species diversity and distribution along rivers floodplain are therefore valuable [5] and could be applied in areas, particularly Mediterranean riparian habitats, where high levels of biodiversity coexist with significant human impact and landscape transformation [6–8]. Such areas are considered highly vulnerable to invasion by alien species [9,10]. In natural and semi-natural ecosystems, native plants may be threatened by invasive alien species [11], introduced deliberately to meet human needs (food, pest control) or accidentally (often due to the globalization of transport). These invasive species can cause enormous damage to ecosystems, livelihoods and human health, and are a major cause of biodiversity loss, especially on Mediterranean islands [11].
Many studies have focused on shifts in dominance between native and alien species, only rarely including endemic species and generally remaining on a local scale, as in experimental patches of vegetation [12,13]. However, other studies provide a landscape perspective of interactions between native/endemic and alien species, and emphasize the importance of such an approach to the quantification of threats from alien species and the establishment of management guidelines [14–16].
Alien species have been demonstrated to be an indicator of natural and human disturbance [17], especially in riparian areas, while endemic species are an indicator of persisting ecological integrity of rivers and can therefore be considered a “quality metric” for watershed planning [18]. Endemic species are native species with unique biological and ecological features [19] and are thought to adapt to natural in-site conditions better than other native flora. However, few studies have focused on the coexistence of important groups of species, such as aliens and endemics and on their distributional pattern along human-impacted rivers [18], despite their recognized importance in defining conservation priorities along rivers [20].
Extensive field surveys to determine plant species composition and distribution over large geographical areas along rivers are not a viable approach because they have many practical limitations and are costly in terms of time and money. Faced with the need for more information and a lack of resources to acquire it, we propose a way of rapidly obtaining information on conservation status at landscape scale in Mediterranean biodiversity hotspots, specifically river floodplains rich in endemic species [21,22].
Longitudinal gradients are seen as the main drivers of vegetation in riparian landscapes and altitude is known to be the main physical factor affecting riparian vegetation along river corridors and directly influencing the erosive power of river flow in the upper and lower reaches [23–25]. This is also true of Sardinian rivers [26]. In this study, we evaluated the relationship of the altitudinal gradient with alien and endemic species along two Mediterranean rivers in Sardinia, in order to model the distribution patterns of these two groups along the main gradient. The Mediterranean island of Sardinia is particularly rich in endemic taxa [27,28] by virtue of its insularity. Together with Corsica and Sicily, it is considered a biodiversity hotspot [21,29]. However, the island has a long history of human settlement, closely linked to water for its survival, as in other semi-arid basins [30,31].
Our basic hypothesis was that endemic and alien species have different patterns of distribution along rivers in relation to different degrees of human/non-natural disturbance, and that their coexistence indicates human impacts on fluvial systems. These impacts can cause loss of natural habitat. If so, the resulting niche segregation and species overlap can be useful preliminary information on riparian zones susceptible to alien invasion or to hosting endemic plants. This could help to identify river sections in which to focus management efforts and areas in need of protection.
2 Materials and methods
2.1 Study areas
The rivers chosen for the study were Rio Leni and Rio Santa Lucia in southwest Sardinia, Italy (Fig. 1). The rivers originate in two of the higher reliefs of southern Sardinia (Monte Lattias 1086 m and Monte Linas 1234 m, respectively) at elevations of 402 m and 541 m, respectively, and are the major rivers of the area. The catchment basins measure 110 and 130 km2, respectively. The Leni and the Santa Lucia have features typical of Mediterranean rivers, including flow regimes subject to erratic fluctuations during the year, with declining flow and almost total drying of the middle sections in late spring and summer, bringing harsh habitat conditions, followed by floods, usually in autumn or early winter. The flora is also typical of these rivers, being rich in terrestrial species related to harsh climate and intermittent flows and with few hydrophytes and helophytes [24,32–34] compared to mesic fluvial systems [35,36].
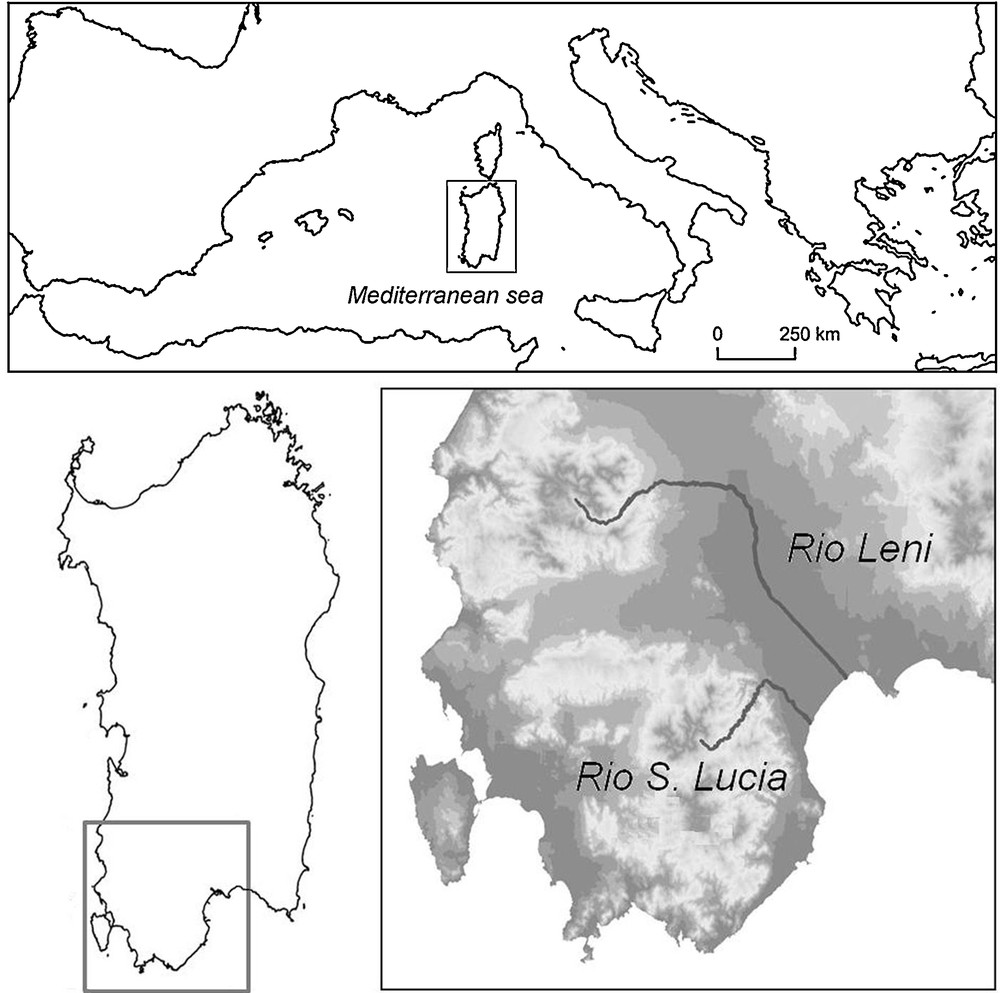
Study area, showing the location of the rivers sampled. In the lower right figure, the Digital Elevation Model (DEM) is shown in the background in grey scale in order to suggest the morphology (white areas have higher elevations).
The rivers flow through land with similar geological and environmental features. The study areas were classified [37] as Mediterranean pluviseasonal oceanic bioclimate, with upper thermo- to lower supra-Mediterranean thermotypes and upper dry to lower humid ombrotypes [22,38].
The rivers were divided into three sections of approximately equal length, based on distinguishing geological and topographic features [39,40] to enable identification of variations in endemic and alien species richness and distribution. The sections were:
- • zone 1 = upper section (from almost 400 to 100 m a.s.l.) composed of granite and metamorphyte outcrops, mostly with single-thread channel morphology and narrow riverbeds (about 25 m);
- • zone 2 = middle section (from 100 to 20 m a.s.l.) where the riverbed was about 60 m wide, including locally braided channels dominated by cobblestones and sands;
- • zone 3 = lower section, with riverbeds about 40 m wide and sediments dominated by clays (from 20 m to sea level).
The upper reaches of both the rivers are in protected areas and Sites of Community Importance (SCI), namely “Monte Linas - Marganai” (ITB041111) in the case of the Leni, and “Foresta di Monte Arcosu” (ITB041105) in the case of the Santa Lucia. These areas are sparsely populated, and human disturbance is associated with low intensity agro-pastoral systems. Woodlands are dominated by Quercus ilex with Q. suber, and riparian forests of Alnus glutinosa. In the middle–lower part, where villages, industries and agricultural land can be found, the riparian woods are dominated by Salix alba, Populus nigra and Nerium oleander subsp. oleander [22], while the fluvial terraces host garigues of Helichrysum microphyllum subsp. tyrrhenicum, Teucrium marum subsp. marum and macchia type vegetation [41,42].
More than two thirds of the Leni flows across the Campidano graben, the largest area of level land in the whole island, while the Santa Lucia is almost equally distributed between granite and sedimentary substrates. Irrigation crops, such as citrus orchards, artichokes and tomatoes occupy the rich alluvial soil of the lowlands. Agriculture and grazing are common along both rivers, although the Leni suffers the greater impact. Human settlements are small and scattered, especially in the Santa Lucia basin. Small industries, mostly related to livestock (goats, sheep, pigs) and cork processing, are dotted throughout the basin. Grazing and water abstraction are the main impacts on the stream channel and adjacent riparian areas, while in the past, there were also mining activities. Both rivers flow into a wetland which is part of the SCI “Stagno di Cagliari, Saline di Macchiareddu e Laguna di S. Gilla” (ITB040023).
2.2 Sampling design and field work
We used a stratified random sampling design. Since we were interested in analysing riparian vascular plant species, the sampling only considered floodplains (identified by selecting areas with alluvial lithology and subtracting cultivated land). The resulting floodplain area was divided into river segments sensu Poole [43], drawn at 2 km intervals along the river from source to mouth, excluding any areas occupied by dams or lagoons. Using the segments as strata, plots were selected along all the river corridors to include variations in species assemblages and environmental features. We randomly selected two 10 × 10 m plots in each segment, obtaining a total of 73 plots (the last segment of one river was less than 2 km, so only one plot was selected there). This plot size was found to be adequate to detect the vegetational mosaic of the floodplain, as it allowed us to explain the relationships between species richness and environmental features in forests and grasslands [44,45]. For each plot, we recorded the presence of all vascular plants from April to July 2010. For species nomenclature, we used the recent checklist of Italian flora [46,47], and other references for endemic species [28], and alien species [48]. Alien status was allocated to species from outside Western and Central Europe and the Western Circum-Mediterranean region and Eastern Europe, whereas endemic species were prevalently Sardinian and Sardo-Corsican, with few Tyrrhenian insular endemics, Tyrrhenian endemics sensu strictu and W-Mediterranean endemics [27–29,49]. Altitude was recorded for each plot using the Digital Elevation Model (DEM).
2.3 Statistical analysis
Species data (presence/absence) from all plots was ordered using Detrended Correspondence Analysis (DCA). This unconstrained ordination was used to find the axis with maximum variation in floristic composition and thus, described the general pattern of species distribution along the gradients [50]. The options chosen for DCA were down weighting rare species and inter-species distances by Hill's scaling [51]; otherwise, the default options were used. The DCA diagram, in which plot distribution revealed a longitudinal gradient, was subsequently projected passively on altitude to show variations across the main gradient.
Generalized Additive Models (GAMs) were then used to construct species response curves along the first DCA axis for endemic and alien species. GAMs are semi-parametric extensions of Generalized Linear Models (GLMs), which assume no a priori responses of a species to an environmental gradient [52]. Options were chosen for the presence/absence data (binomial distribution and maximum value). Two degrees of freedom and stepwise selection by Akaike Information Criterion (AIC) were employed to select the best GAMs. Canoco 4.5 for Windows [51] was used for the analyses.
3 Results
A total of 428 taxa were found during the study, of which 22 were endemic species (8.4% of the island's total endemic flora) [28], and 14 were alien species (about 3% of the species found during this study). About half of the total endemics and aliens (16) were common to both rivers (see Table 1) and most endemics found are linked to riparian zones (Carex microcarpa, Mentha suaveolens subsp. insularis, Hypericum hircinum subsp. hircinum, Eupatorium cannabinum subsp. corsicum, Salix arrigonii, Apium crassipes, Aristolochia navicularis, Euphorbia meuselii, Torilis nodosa subsp. nemoralis, Leucanthemum flosculosum, Polygonum scoparium) [22,53]. There were also some endemic species not strictly linked to riparian habitats but frequent in floodplains with xeric substrate, being well adapted to harsh habitats, like river alluvial terraces and incoherent soils: they were prevalently chamaephytes, such as Teucrium marum subsp. marum, Genista corsica, Stachys glutinosa, Dephinium pictum, Rumex scutatus subsp. glaucescens and Helichrysum microphyllum subsp. tyrrhenicum.
Endemic and alien species of the two rivers.
| Endemic species | River |
| Apium crassipes | L |
| Aristolochia navicularis | L |
| Arum pictum subsp. pictum | S |
| Bellium bellidioides | L, S |
| Bryonia marmorata | L, S |
| Carex microcarpa | L, S |
| Crepis bellidifolia | L |
| Cymbalaria aequitriloba subsp. aequitriloba | L, S |
| Delphinium pictum | S |
| Eupatorium cannabinum subsp. corsicum | L |
| Euphorbia meuselii | L, S |
| Genista corsica | L |
| Helichrysum microphyllum subsp. tyrrhenicum | L, S |
| Hypericum hircinum subsp. hircinum | L, S |
| Mentha suaveolens subsp. insularis | L, S |
| Leucanthemum flosculosum | L, S |
| Polygonum scoparium | L, S |
| Rumex scutatus subsp. glaucescens | L |
| Salix arrigonii | S |
| Stachys glutinosa | L |
| Teucrium marum subsp. marum | L, S |
| Torilis nodosa subsp. nemoralis | L |
| Alien species | |
| Acacia mearnsii | L |
| Acacia saligna | S |
| Asclepias fructicosum | S |
| Arundo donax | L, S |
| Avena sativa | L |
| Brassica napus | L |
| Chenopodium ambrosioides | L |
| Conyza bonariensis | L, S |
| Eucalyptus camaldulensis | L, S |
| Oxalis pes-caprae | L, S |
| Senegalia visco | S |
| Symphyotrichum squamatum | L, S |
| Xantium spinosum | L |
| Xantium strumarium | L |
The longest gradient of the DCA (Fig. 2) performed on the whole species data set was 4.09 SD, underlining high floristic heterogeneity. The first axis, explaining 8.6% of variability, represented the main gradient of the rivers. This was highly negatively correlated with altitude (r = –0.79; P < 0.01) and clearly represented the longitudinal gradient of the rivers. The pie charts at the bottom of Fig. 2 show white slices of alien species much wider towards the right side of the first axis, while the black slices of endemic species were wider towards the left. In the middle part of the main gradient, endemic and alien species occurred together with quite similar high frequencies.
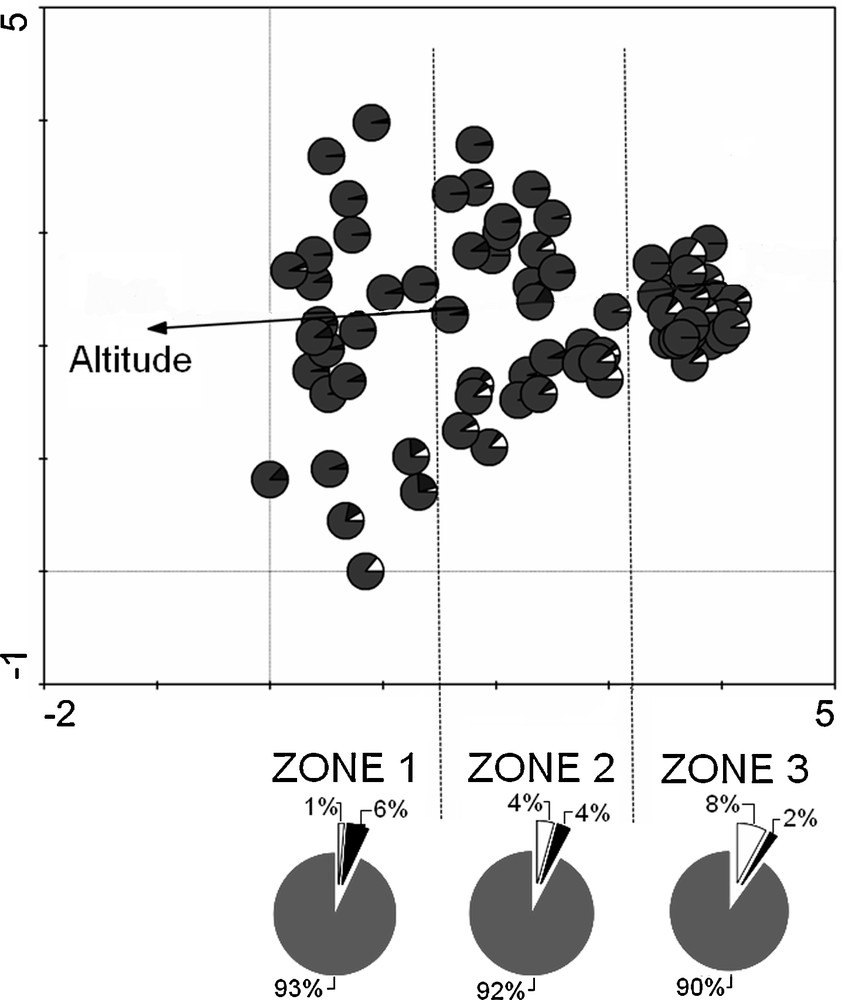
DCA ordination diagram with pie charts for plots, showing the distribution of endemic (black slice) and alien (white slice) species in relation to the other plant species in the plot (grey slices). Arrow indicates altitude. The pie charts (below) show the relative percentage frequency of occurrence of alien and endemic species in the three river sections (zone 1 = upper; zone 2 = middle; zone 3 = lower).
In order to investigate how endemic and alien species varied along the main gradient, species response curves were performed along the first DCA axis. Many endemic and alien species presented a significant response to the GAMs (P < 0.01) and were distributed all along the longitudinal gradient, although the distribution pattern differed between groups. The response curves for endemic species (Fig. 3A) showed that these species have the highest probability of occurrence in correspondence with the medium and lower values of the first axis, except for Aristolochia navicularis, which increased in correspondence with the highest values. The response curves for alien species (Fig. 3B) showed that most have a higher probability of occurrence corresponding with the medium and higher values of the first axis, except for Acacia mearnsii, which decreased in correspondence with lower values.
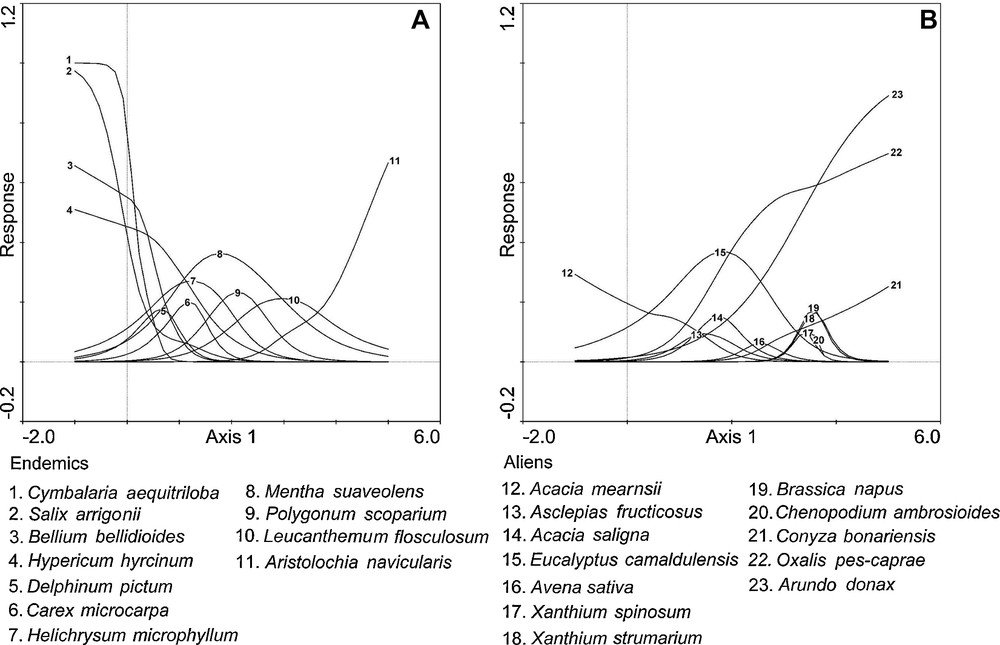
Response curves of endemic (A) and alien (B) species to the first DCA axis. All species were significant at the 0.01 level.
4 Discussion
The approach proposed here, defining the niches achieved by endemic species along Mediterranean riparian corridors with respect to alien species, allowed us to obtain general information about the main criticalities of the rivers, which in most cases were directly due to human impact.
Endemic species are usually scarce in riparian areas, as found by several authors in studies regarding rivers in the Mediterranean basin [18,54]. In contrast, our results indicate a large number of endemic species along both rivers, as already observed by others [22,27,49]. This result is probably due to the isolation and high geological diversity of the island [21] and can be considered specific to insular regions, confirming that distributional patterns of endemism can be used to establish conservation priorities in the riparian habitats of biodiversity hotspots [21]. This study highlighted that the largest group of endemics, in terms of number of species, included entities linked to riparian zones. However, there were also a variety of terrestrial entities with occasional occurrence in riparian corridors. These species are a result of lateral connections between riparian and terrestrial ecosystems at higher altitude, due to the fact that riverbanks are small and zonal vegetation is in close contact with riparian vegetation, while nearby Mediterranean shrublands play a major role in their diversity [8]. Examples include the sclerophyllous and evergreen shrubs Cistus sp., Lavandula sp. and Cytisus striatus (Hill) Rothm.
Alien richness was substantially low, a pattern already found in other Mediterranean rivers in Portugal and Spain [55–57], even if their frequency was higher in the lower stretches. The ordination carried out on the whole floristic data set, with superimposed supplementary variables, confirmed the longitudinal gradient to be the main driving force in the distribution of floodplain flora. The importance of the longitudinal gradient was further underlined by the endemic and alien species, which revealed GAMs, almost all of which had high statistical significance along this gradient. As expected, the distribution patterns of most aliens and endemics along the longitudinal gradient were quite well differentiated. In fact, the response curves showed that endemic species had their maximum occurrence probability in the upper and middle stretches of the rivers, while alien species had theirs in the middle and lower stretches. The only exceptions to this trend were Aristolochia navicularis and Acacia mearnsii: the former more likely to be found in the lower parts of the rivers, where it is threatened by invasion of its habitat by Arundo donax [58]; the latter found mostly in the upper stretches where it presumably escaped from nurseries. The distribution of alien species in lowlands can be regarded as a consequence of substantial human impact and high nitrogen availability, mainly due to agriculture [48,59]. In line with many other authors, we found that invasion of riparian areas by aliens was closely linked to habitat degradation by human activities, which is always greater in the vicinity of residential areas and in cultivated lowlands [2,60], such as those of our rivers [58,61]. These conditions reduce the competitiveness of native species, which are very sensitive to environmental modifications [62,63]. Our findings may indicate that endemics and aliens compete in such riparian zones, and that some alien species may displace native flora. Nevertheless, we highlight that:
- • in the Mediterranean basin, including near-natural rivers, it is possible to observe a natural pattern with endemic species prevalently distributed in mountain river corridors, while riverbanks close to the sea host very few endemic species [18,54];
- • there is limited experimental and observational evidence that endemic species are endangered by invasions of alien plants [64].
Endemic loss and extinction in the Mediterranean basin are more likely to be a result of human-induced habitat loss [65] than that of the spread of alien plants, which have been persistently linked to human disturbance in riparian ecosystems [18]. In such lowland systems, the high probability of occurrence and diversity of aliens in contrast with the almost total lack of endemics may indicate that on riverbanks with N-rich soils (human activity, diffuse pollution, etc.), only strongly competitive species, such as aggressive native (e.g. Phragmites, Typha, etc.) and alien plants can survive and maintain good populations. On the contrary, endemics, which were documented in lowland areas until the early 1900s [38], cannot survive. The rivers are so degraded and transformed that native vegetation with high biodiversity could only be recovered by laborious, costly, intensive and extensive restoration.
In contrast, endemic species, such as Hypericum hircinum subsp. hircinum, Carex microcarpa and Salix arrigonii [22,66], closely linked to the most natural shrubby and woody riparian habitats, showed the highest occurrence probability and took refuge in the upper parts of rivers, indicating woody/shrubby riparian habitats with only limited experience of human disturbance and transformation, conditions which are further confirmed by the very low presence of aliens. Maintaining these habitats serves as a significant barrier to the spread of invasive species [67], as well as improving many other ecosystem functions, such as the resistance of soils to erosion during floods, frequent in the area in autumn and spring [22]. Moreover, the fact that the endemic species did not seem to be threatened in the upper part of these rivers may also be related to the buffer effect of native vegetation of the typical shrubby maquis, with its high resilience and resistance to invasion [68–70]. To conserve ecological integrity here, it may be sufficient simply to maintain the natural woody riparian vegetation, which is presumably capable of protecting natural areas from the impact of alien species [71]. Helichrysum microphyllum subsp. tyrrhenicum, Polygonum scoparium and Leucanthemum flosculosum or Delphinium pictum, endemic species that like other Mediterranean endemics [72] are largely stress-tolerant and perfectly adapted to harsh areas, can widely colonize the alluvial terraces and incoherent soils of the middle parts of rivers, where, however, many aliens had their highest occurrence probability. These included woody species linked to reforested areas and “unintentionally introduced” species [73] associated with cultivations, such as Avena sativa and Eucalyptus camaldulensis, which are only rarely a danger to native vegetation.
The coexistence of endemic and alien species underlined the presence of areas important for riparian flora conservation in the middle stretches of the river, which are also affected by human disturbance in the river corridor and modifications to the surrounding landscape. This coexistence may also be related to morphological transitions that influence landforms (incised valleys/level land) [74]. Endemic species closely linked to riparian habitats in the middle sections (such as Carex microcarpa, Eupatorium cannabinum subsp. corsicum, Euphorbia meuselii, Hypericum hircinum subsp. hircinum, Mentha suaveolens subsp. insularis and Salix arrigonii), which seemed unable to colonize other more xeric habitats in the upper sections, or more nitrified habitats towards the lowlands, should be considered sensitive indicators. This is why what currently seems a situation of equilibrium and coexistence between endemic and alien species actually requires careful control, especially because many studies have demonstrated the ability of some alien species to displace native flora in rivers through monospecific spread on river margins and in aquatic ecosystems [10]. These are the most fragile areas, where the challenge of aliens towards endemics, and more generally between natural and human-transformed habitats, is occurring now and will occur to a greater extent in the future. The upper sections of the rivers are in protected areas (SCI) and their riparian vegetation structure is still largely determined by natural dynamics [22,32] and cannot be considered a source of alien species. The fastest and easiest way to avoid this danger is therefore to reduce human pressure, maintaining and restoring natural habitats and vegetation cover, and at the same time, reducing the probability of alien invasion in areas not yet under conservation order but where endemics are nonetheless present. Since riparian ecosystems are important for life at all levels, our results stress that comprehensive management of alien invasive species and protection of endemic species should include monitoring of the quality and ecological integrity of terrestrial riparian habitats and adjacent freshwater ecosystems [32], and surveillance of known endemic populations.
This study highlights the utility of our analysis using alien and endemic plant species distribution models as a starting point for obtaining information about the more critical sectors of Mediterranean-type rivers, especially in the islands where the percentage of endemic species is high, even in riparian habitats. Like many other rapid assessment tools of plant species diversity and distribution, this approach, however, simplifies nature by representing only the main (longitudinal) gradient, ignoring other important ones, such as geomorphological gradients. Moreover, protection of specific riparian areas along rivers may have limited impact on conservation because many alien species come from outside the protected area. In any case, our approach can be regarded as a preliminary, rapid, cost-effective tool for conservation planning at landscape scale. The easily drawn zones with different protection needs can facilitate and accelerate effective conservation and management planning in riparian zones.
Disclosure of interest
The authors declare that they have no conflicts of interest concerning this article.
Acknowledgments
We are particularly grateful to Tiziana Carai, Pasqualina Casula, Daniela Fanti, Martino Orrù and Cristiano Pontecorvo for their help in field work, and to Lina Podda and Francesco Mascia for their help with plant determinations.