

1 Introduction
Family Euscorpidae Laurie, 1896 is globally distributed and is represented by ten genera and ninety-nine species [1]. Of these, three genera, viz. Euscorpiops Vachon, 1980, Scorpiops Peters, 1892 and Neoscorpiops Vachon, 1980, have been reported from India. In India, Euscorpiops and Scorpiops are primarily distributed across the western Himalayas and in northeast India (a single Scorpiops species has been reported from central India), while Neoscorpiops is restricted to the northern Western Ghats [2,3]. Among the Indian Euscorpids, the genus Scorpiops is most well represented, with at least nine species viz. Scorpiops braunwalderi Kovařík 2000, S. dastychi Kovařík 2000, S. demise Kovařík 2005, S. feti Kovařík 2000, S. hardwickei Gervais 1843, S. leptochirus Pocock 1893, S. pachmarhicus Bastawade 1992, S. petersii Pocock 1893 and S. rohtangensis Mani 1959 currently reported from India [3–5]. A majority of these species typically occur at high-elevations (< 1000 m) with some species occurring at elevations greater than 3700 m.
Recently, individuals from one such high-elevation (4000–4200 m) Scorpiops population from Spiti Valley in Himachal Pradesh were collected. Morphologically, these individuals show affinity to S. petersii. Detailed comparisons of the collected individuals with the lectotype of S. petersii however indicate significant differences between the two, which warrant the individuals from Spiti Valley to be considered a distinct taxon. In this paper, we present a detailed description of the collected types and provide diagnostic characters and notes on its natural history.
2 Methods
Specimens of the new species are stored in 70% ethanol in the collection of the Bombay Natural History Society (BNHS, Mumbai). Measurements were taken with a Mitutoyo™ digital calliper with an error of 0.1, and morphological details were recorded using an Olympus™ SZ40. Measurements were taken following Stahnke [6] and are provided in millimetres (mm). Trichobothrial nomenclature follows Vachon [7] and morphological terminologies follow Hjelle [8]. Acronyms used in the manuscript are as follows: BNHS – Bombay Natural History Society, Mumbai; NHM – Natural History Museum, London.
In addition to the classical diagnostic characters, we propose a new method to quantify cuticular granulation that is an important diagnostic character in scorpion taxonomy. Granulation has until now been presented largely descriptively, e.g., dense, sparse, moderately dense, etc. This method, though informative, is prone to personal subjectivity and bias. Moreover, it fails to provide an absolute measure of granulation, making comparisons between closely related species complicated, especially when types cannot be accessed. To counter this issue, we propose a novel yet highly simplistic technique that allows quantitative assessment of granulation. To quantify granulation, photographs (with scale) of the types of the new species and the closely related S. petersii were taken and resized to 300 dpi. Grids of 10 pixel2 were overlaid on the image in Adobe Photoshop CS3 and percent area coverage of granules was calculated by counting the total number of grids required to cover the focal body part and the number grids among them that showed the presence of granules.
3 Taxonomic treatment
Family EUSCORPIIDAE Laurie, 1896
Subfamily SCORPIOPINAE Kraepelin, 1905
Scorpiops spitiensis sp. nov. (Figs. 1–4)

(Colour online.) Scorpiops spitiensis sp. nov. holotype male Bombay Natural History Society Sc-67, (a) dorsal view, (b) ventral view. Scale bar: 10 mm.

(Colour online.) Scorpiops spitiensis sp. nov. holotype male Bombay Natural History Society Sc-67 (a–c), (a) carapace, (b) mesosoma, (c) telson. Scorpiops petersii lectotype make British Museum of Natural History (BMNH) (c–e), (c) carapace, (d) mesosoma, (e) telson.

(Colour online.) Scorpiops spitiensis sp. nov. holotype male Bombay Natural History Society Sc-67 manus (a) dorsal view, (b) external view, (c) ventral view, patella (d) dorsal view, (e) external view, (f) ventral view.
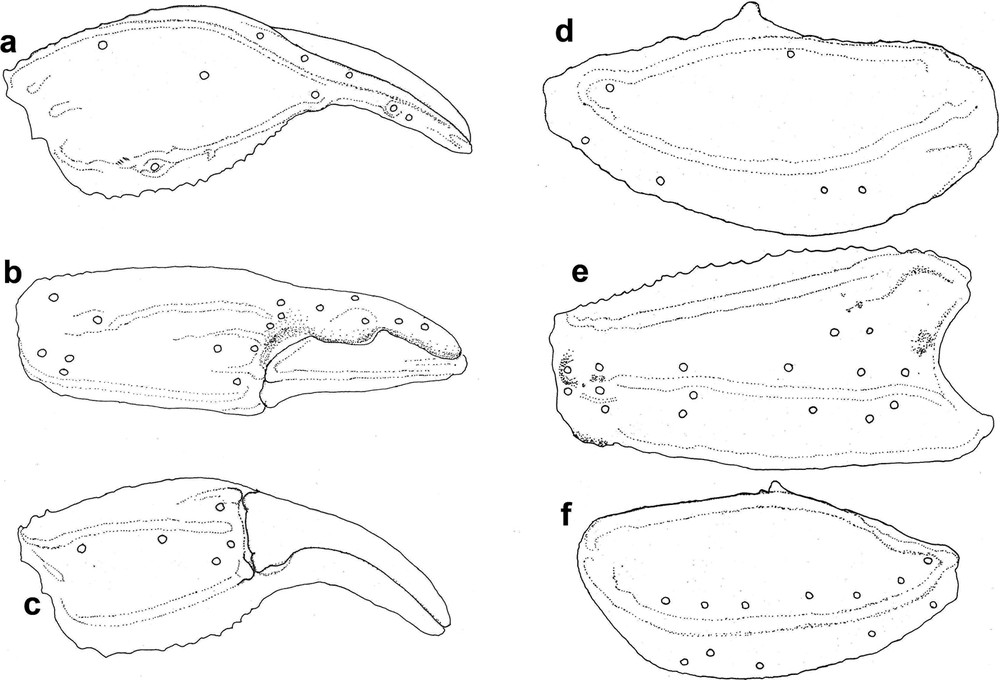
Scorpiops spitiensis sp. nov. holotype male Bombay Natural History Society Sc-67 showing trichobothrial pattern on manus (a) dorsal view, (b) external view, (c) ventral view, patella (d) dorsal view, (e) external view, (f) ventral view.
urn:lsid:zoobank.org:act:32A6146C-6336-4024-869E-598F05ED37BA
Holotype. BNHS Sc-67, adult male collected by Mayank Kohli and Ashwin Viswanathan from Kibber village, Spiti Valley, Himachal Pradesh, India (32.3309N, 78.0072E, altitude 4200 m).
Paratypes. One female BNHS Sc-68 and one male BNHS Sc-69 by Kulbhushansingh Suryawanshi; 1 female BNHS Sc-70 by Mayank Kohli and Ashwin Viswanathan. Collection locality same as the holotype.
Etymology. The specific epithet refers to the type locality “Spiti” where the types were collected.
3.1 Diagnosis
A medium-sized species in relation to members of the genus Scorpiops (39.51 mm maximum). Carapace anterior margin raised upwards. Median eyes situated anteriorly in the ratio 1:3. Patella with 7 ventral trichobothria and 16 external trichobothria, single em present. Manus flexed in males. Mesosomal tergites I–VI with median carinae, posterior and laterally portion of tergites with depressed granulation. Telson moderately bulbous. Pectines 5 in number.
3.2 Description of holotype BNHS Sc-67
Colouration (Fig. 1a & 1b, Table 1). Overall in a shade of brownish black with the chelicerae being yellowish brown with dark brown reticulations. Manus and telson with a slight reddish ting. Pectines and genital operculum pale yellowish. Sternites dark brown.
Morphometric details of the type series of Scorpiops spitiensis sp. nov.
| Characters | BNHS Sc-67 Holotype | BNHS Sc-68 Paratype | BNHS Sc-69 Paratype | BNHS Sc-70 Paratype | Scorpiops petersii Lectotype |
| Total length | 39.51 | 33.13 | 31.08 | 26.56 | 69.3 |
| Carapace length | 6.91 | 5.59 | 5.55 | 4.67 | 8.8 |
| Carapace anterior width | 3.99 | 4.11 | 3.2 | 2.62 | NA |
| Carapace posterior width | 6.56 | 5.68 | 5.75 | 4.38 | 8 |
| Mesosomal length | 15.24 | 10.52 | 9.68 | 10.63 | NA |
| Metasoma total length | 17.36 | 17.02 | 15.9 | 11.26 | 34.7 |
| Metasomal seg I length | 1.84 | 2.26 | 2.16 | 1.35 | 3.5 |
| Metasomal seg I width | 2.81 | 2.54 | 1.91 | 1.72 | 3.5 |
| Metasomal seg II length | 2.58 | 3.34 | 2.8 | 1.89 | 4.1 |
| Metasomal seg II width | 2.41 | 2.36 | 1.33 | 1.35 | 3 |
| Metasomal seg III length | 3.38 | 2.87 | 2.56 | 2.06 | 4.4 |
| Metasomal seg III width | 2.3 | 2.15 | 1.71 | 1.37 | 2.8 |
| Metasomal seg IV length | 3.7 | 3.25 | 3.33 | 2.47 | 5 |
| Metasomal seg IV width | 2.28 | 2.04 | 1.75 | 1.4 | 2.6 |
| Metasomal seg V length | 5.86 | 5.3 | 5.05 | 3.49 | 8.2 |
| Metasomal seg V width | 2.13 | 1.9 | 1.57 | 1.36 | 2.3 |
| Telson length | 6.29 | 4.74 | 4.12 | 3.81 | 8.7 |
| Femur length | 4.65 | 4.54 | 4.06 | 3.29 | 7.2 |
| Femur width | 2.13 | 1.96 | 1.82 | 1.31 | 3.3 |
| Patella length | 5.47 | 4.34 | 4.07 | 3.36 | 7.2 |
| Patella width | 2.27 | 1.84 | 1.8 | 1.49 | 3.4 |
| Chela length | 10.94 | 9.02 | 8.45 | 7.36 | 15.1 |
| Chela width | 4.34 | 3.09 | 2.92 | 2.1 | 5.8 |
| Movable finger length | 6.06 | 5.12 | 4.42 | 4.04 | 7.5 |
| Pectines L/R | 5/5 | 6/6 | 6/6 | 5/5 | 5/5 |
Morphology. Carapace anterior margin with a moderately deep ‘U’ shaped indentation with its edges along the lateral eyes and anterior margin raised upwards. Surface with coarse granulation throughout. Carapacial carinae indistinct. Median eyes situated anteriorly in the ratio of 1:3. Median and lateral ocular tubercle glossy black (Fig. 2a). Three pairs of lateral eyes. Mesosoma (Figs. 1a, 1b & 2b) – Tergites coarsely granular throughout with large depressed granules concentrated on the lateral and posterior region of each segment. Median carinae fairly distinct on segments I–VI, lateral carine indistinct, merely represented by concentrated granules. Pectines 5 in number with fulcra present. All sternites smooth and glossy lacking carinae except sternite VII, which bears two pairs of lateral carinae. Metasoma – First metasomal segment with 10 keels (dorso-median vestigial, merely represented by a few granules), II–IV segment with 8 keels (latero-median absent), segment V with 7 keels. Keels on segment V serrated. Vesicle sparsely granular with a clear smooth band on its lateral aspect (Fig. 2c). Pedipalp (Figs. 3a–f & 4a–f) – Trichobothrial pattern C, neobothriotaxic [7]. Patella with 16 trichobothria on its external (5 eb, 2 esb, 1 em, 4 est, 4 et; Figs. 3e, 4e) and 7 on its ventral aspect (Figs. 3f, 4f). Patella with a large tooth and a few small ones on its internal aspect. Movable finger with a single row of granules. Manus length to width ratio 1:2.5. Chelicerae – Fixed finger with one large tooth at distal end and a small bifid tooth. Movable finger with two rows of teeth, from 4–6.
Distribution and natural history notes. The type locality, Spiti Valley, is located in western Himachal Pradesh in northern India (Fig. 5). Spiti Valley is a cold, high-altitude desert characterized by a mosaic of rolling hills, rocky cliffs and outcrops with altitudes varying from 3000 to 5000 m. Mean summer temperature is around 25 °C, but however can drop to −35 °C during winters. Precipitation is in the form of snow during months of December to March. The vegetation of the area is classified as dry alpine steppe and is dominated by several shrub species [9]. The types were found while ground was being dug for the construction of houses at 4000–4200 m (Fig. 6), but however might occur at higher elevation, but dedicated surveys need to be conducted to ascertain the higher limits of distribution of the new species. Local residents report this species to be common while digging during summer months indicated fossorial habits. Given that the type locality remains covered with snow for more than 4 months, it is likely that this species hibernates during the cold months of the year and surfaces only during the hotter months of the year.
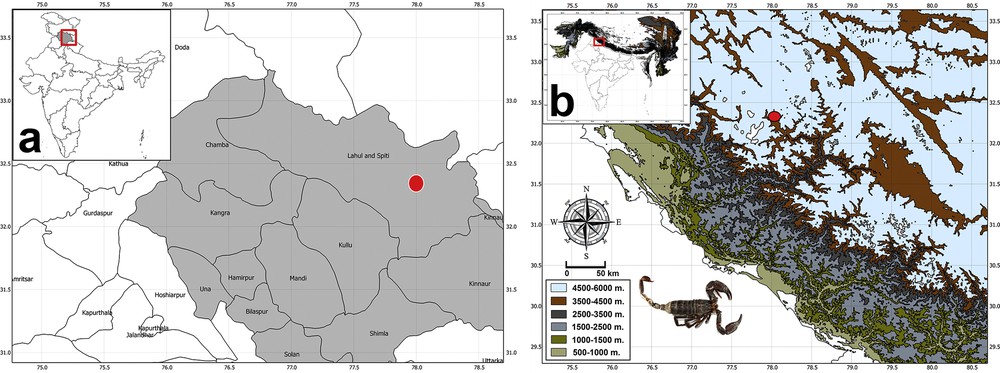
(Colour online.) Map showing the type locality of Scorpiops spitiensis sp. nov. (a) type locality indicated by a solid red circle in the Indian state of Himachal Pradesh in a political map, (b) type locality highlighted by a solid red circle in a map showing elevational gradient in the Himalayas. The inset map shows the location of Himachal Pradesh in India in a shade of grey and in a red box.

(Colour online.) Images depicting the type locality of Scorpiops spitiensis sp. nov. in two different seasons (a) winter, (b) summer. Types were collected from under such houses while digging to renovate the houses.
Photo by Kulbhushansingh Suryawanshi.
3.3 Relationship
The new species closely resembles S. petersii, but however differs from it as follows:
- • maximum length 39.51 mm (average length 69.3 mm in S. petersii);
- • 16 trichobothria on external aspect of patella (17 in S. petersii);
- • carapace anterior margin with a moderately deep ‘U’ shaped indentation with its edges along the lateral eyes and anterior margin raised upwards (‘U’ shaped indentation deep with anterior margin not raised in S. petersii, Fig. 2);
- • anterior eyes situated medially in the ratio of 1:3 (1:2.6 in S. petersii, Fig. 2);
- • mesosomal tergites coarsely granular throughout with large depressed granules concentrated on the lateral and posterior region of each segment. Percent granulation on mesosomal segment IV: 70%, V: 87% and VII: 92% (granulation sparse and restricted to the posterior portion. Percent granulation on mesosomal segment IV: 50%, V: 55% and VII: 45% in S. petersii, Fig. 2);
- • telson moderately bulbous, vesicle with 51% granulation 2.5 times that of the aculeus (telson bulbous, vesicle with 25% granulation 3 times that of the aculesus in S. petersii, Fig. 2).
Variation: Paratypes resemble the holotype in all aspects except for the females, which do not have a flexed manus.
4 Discussion
The occurrence of the new species at 4200 m represents the highest-elevation record of any scorpion in India and the second highest to the best of our knowledge in the whole of Asia. The highest record for a scorpion species reported from Asia is ∼ 4600 m by Tibetiomachus himalayensis Lourenço and Qi 2006 [10], followed by Scorpiops rohtangensis Mani 1959 [3], the holotype of which is missing from the collection of the Zoological Survey of India, Kolkata. Few other species are known to inhabit elevations higher than S. spitiensis sp. nov. with the exception of Orobothriurus crassimanus Maury, 1975 from the Andes, which have been reported from altitudes of up to 5500 m [11].
S. spitiensis sp. nov. shows close affinities to S. petersii, which is likely a species complex comprised of multiple cryptic species. S. spitiensis is possibly one of the cryptic species of this complex, has only 16 trichobothria on the external surface of the patella – one of strongest diagnostic character that separates it from S. petersii. The members of the genus Scorpiops show large variation in the number of trichobothria (17–21) on the external aspect of the patella. The presence of only 16 trichobothria in the new species sets it apart from all known Scorpiops species, with it being the only species exhibiting this unique character. Whether this character alone warrants the erection of a new genus to accommodate the new species as yet remains unclear. Though similar differences have been used in the past to separate Euscorpid genera, we prefer to be conservative in our approach, acknowledging a potential need to do so if more species exhibiting the unique character are revealed in the future.
Acknowledgments
ZM & RS were able to visit the Natural History Museum, London through a travel grant from the Newby Trust Limited to ZM. Nature Conservation Foundation (NCF, Bangalore) for help with logistics. Special thanks are addressed to Kulbhushansingh Suryawanshi (NCF, Bangalore), Mayank Kohli and Ashwin Viswanathan for providing specimens, images and data on the species as well as its habitat; without them, this publication would not have been possible. Rahul Khot helped with registration of the types at the Bombay Natural History Society, Mumbai, and Janet Beccaloni granted access to specimens at Natural History Museum, London. For logistic support and lab facilities we acknowledge support of Krushnamegh Kunte (NCBS, Bangalore). Special thanks go to an anonymous reviewer for his constructive comments, from which the manuscript benefited.