

1 Introduction
As outlined in recent papers [1–3], the taxonomy of the subfamily Scorpiopinae can be considered as complex. Scorpiopinae were first proposed by Kraepelin [4] as Scorpiopsinae, a subfamily of Vaejovidae. Lourenço [5] confirmed the previous decision by Stockwell [6] to raise Scorpiopinae to the family level. Subsequently, Soleglad and Sissom [7] downgraded Scorpiopidae to a subfamily of Euscorpiidae, and grouped the Asian genera into the tribe Scorpiopini, also including in this subfamily the North American genus Troglocormus (tribe Troglocormini). According to morphological assessments, the subfamily Scorpiopinae currently forms a monophyletic group within Euscorpiidae and does not share any synapomorphies with North American Vaejovidae [7]. The tribe Scorpiopini includes six Asian genera, mainly from the southern and southeastern regions of the continent. The composition of these genera is mainly due to Vachon [8], who revised Scorpiops and described three new subgenera – Alloscorpiops, Euscorpiops and Neoscorpiops – in addition to the nominotypical subgenus, Scorpiops. These four subgenera were later elevated to the generic rank by Lourenço [5], who added the monotypic genera Parascorpiops Banks 1928 and Dasyscorpiops Vachon, 1974 to the tribe, bringing the total number of genera to six. Soleglad and Sissom [7] then supported the validity of Euscorpiops based on the position of chelal trichobothrium Eb3 and the presence of an annular ring on the telson. Recently, Lourenço [1] divided the genus Alloscorpiops into two subgenera, with the creation of the subgenus Laoscorpiops. Since most historical aspects concerning these decisions have been concisely synthesized by Lourenço [1,2] and Lourenço and Pham [3], they will not be further discussed here. In the present note, a new species belonging to the genus Euscorpiops is described from caves in Central Vietnam. This new scorpion taxon is the second species of Scorpiopinae to be discovered in a cave system and may be yet another endemic element in the fauna of this country.
2 Cave scorpions
Cave or, more precisely, true troglobitic scorpions are globally rare. It was not until the late 1960s that the first truly troglobitic scorpions were described from Mexican caves: Typhlochactas rhodesi Mitchell, 1968 and T. reddelli Mitchell, 1968 [9]. Since these initial discoveries, the available data on cave scorpions have been synthesized by certain authors, in particular Lourenço and Francke [10] and Volschenk and Prendini [11]. The true troglobitic scorpions discovered to date belong to the families Akravidae Levy, 2007, Chaerilidae Pocock, 1893, Diplocentridae Karsch, 1880, Euscorpiidae Laurie, 1896, Hormuridae Laurie 1896, Pseudochactidae Gromov 1998, Typhlochactidae Mitchell, 1971, Troglotayosicidae Lourenço, 1998, Urodacidae Pocock, 1893 and Vaejovidae Thorell, 1876. These troglobitic species have been discovered mainly in North and South America, the Middle East, Madagascar, Christmas Island (Indian Ocean), Australia and Southeast Asia. For a more detailed synopsis, see Lourenço and Pham [3].
Until recently, no species of the subfamily Scorpiopinae had been reported inhabiting cave environments. The first case concerned Euscorpiops cavernicola Lourenço and Pham, 2013 from Vietnam [3]. The new species described here represents the second known species of cave-dwelling scorpion belonging to the subfamily Scorpiopinae. Some of its morphological features indicate a certain degree of adaptation to cave life.
3 Methods
Scorpions were found on the walls of the cave, approximately 60–70 m from the main cave entrance. Illustrations and measurements were produced using a Wild M5 stereo-microscope with a drawing tube and an ocular micrometer. Measurements follow Stahnke [12] and are given in millimeters. Trichobothrial notations follow Vachon [13] and morphological terminology mostly follows Vachon [14] and Hjelle [15].
4 Taxonomic treatment
Family Euscorpiidae Laurie, 1896
Subfamily Scorpiopinae Kraepelin, 1905
Genus Euscorpiops Vachon, 1980
Euscorpiops dakrong sp. n. (Figs. 1–3)

(Color online). Euscorpiops dakrong sp. n. A–D. Male holotype and female paratype. Habitus, dorsal and ventral aspects (scale bars = 1 cm).
Euscorpiops dakrong sp. n. Male holotype (A, B, D, F) and female paratype (C, E). A. Carapace. B–C. Metasomal segment V and telson, lateral aspect. D–E. Ventral aspect showing sternum, genital operculum and pectines. F. Cutting edge of movable chelal finger.

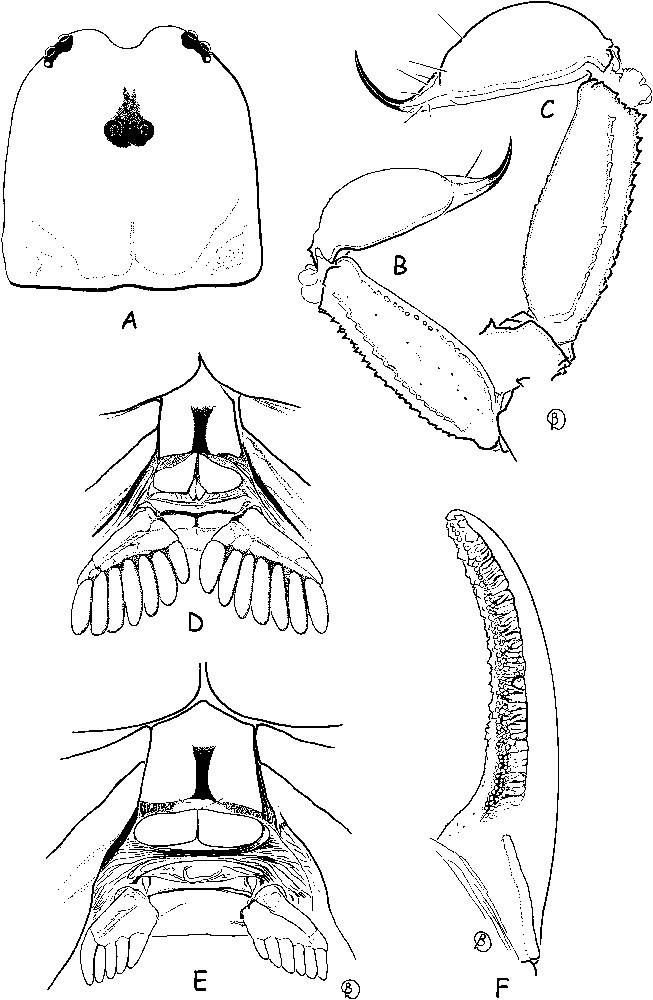
Euscorpiops dakrong sp. n. Male holotype (G–M, P) and female paratype (N, O). Trichobothrial pattern. G–J. Chela, dorso-external, external, ventral and internal aspects. K–O. Patella, dorsal, ventral and external aspects. P. Femur, dorsal aspect.
Vietnam, Quang Tri Province, Dakrong district, Dakrong Nature Reserve cave system (precise cave not identified), 60–70 m from entrance, 3/XII/1958 (B. Dejenböl). Male holotype and female paratype deposited in the Muséum national d’histoire naturelle, Paris.
Etymology: the specific name is a noun in apposition to the generic name and refers to the region where the new species was found.
Diagnosis: The new species exhibits the general characteristics of the genus Euscorpiops [7,8]. Total body length medium to small relative to other species of the genus. Total length for adult males and female 27.3 and 25.7 mm, respectively. General coloration yellow to reddish–yellow. Body and pedipalps moderately slender. Internal aspect of patella with two small spinoid tubercles of similar size. Trichobothrial pattern with three trichobothria on femur: dorsal, internal and external. Patella with 2 dorsal, 1 internal, 9 ventral and 17 external trichobothria in both sexes. Chelal manus with 4 ventral, 2 dorsal (Dt, Db), 2 internal (ib, it), 1 Est, 5 Et, 1 Esb and 3 trichobothria in the Eb series. Trichobothrium Eb3 is distal in relation to Eb2 [8,13].
Description. Coloration yellow to reddish-yellow. Carapace reddish-yellow with paler zones on anterior edge and on furrows. Tergites reddish-yellow. Metasomal segments reddish-yellow; telson yellow; base of aculeus yellow and tip reddish. Chelicerae yellow with inconspicuous variegated spots, these better marked at the base of the teeth. Pedipalps reddish-brown; fingers darker than chela manus. Legs yellow. Venter globally yellow, without infuscations.
Morphology. Carapace moderately granular, granulation better marked in female; furrows moderately deep. Median eyes anterior to middle of carapace; three pairs of lateral eyes, the third pair only slightly smaller than the first two. Sternum pentagonal, wider than long. Tergites moderately granulated; VII with five carinae, moderately marked. Pectines very large in male, reduced in female; pectinal tooth count 7–7 in male, 6–5 in female; fulcra absent in both sexes. Sternites almost smooth and punctate; sternite VII with four weakly marked carinae and little granulation. Metasomal segment I wider than long; segments II to V longer than wide; 10–8–8–8–7 carinae present on segments I–V; dorsal carinae on segments II–IV with a single, strong, posterior spinoid granule; metasomal tegument weakly granulated; ventral carina on segment V with moderately marked spinoid granules. Telson vesicle smooth and without granulation. Setation weak on metasomal segments and telson. Pedipalps: femur with dorsal internal, dorsal external, ventral internal and ventral external carinae moderately marked; tegument moderately granular, better marked in the female. Patella with dorsal internal, dorsal external, ventral internal, ventral external and external carinae moderately marked; two moderately marked spinoid granules of similar size present on the internal aspect; tegument moderately granular. Chela with dorsal marginal, external secondary, ventral internal and ventral carinae moderately marked; other carinae moderately to weakly marked; tegument granulated dorsally and ventrally. Chelal fingers with two longitudinal series of granules, almost fused, a few inner and several outer accessory granules. Cheliceral dentition as defined for the family [16]; 4–5 teeth on the ventro-internal face of the movable finger. Trichobothriotaxy type C, as shown in Fig. 3 (G-p) [13]: see diagnosis for additional details.
Morphometric values (in mm) of the male holotype/female paratype. Total length (including telson) 27.3/25.7. Carapace: length 4.1/3.8; anterior width 2.6/2.4; posterior width 4.0/4.2. Mesosoma length 8.3/9.1. Metasomal segment I: length 1.6/1.2, width 1.8/1.7; II: length 1.7/1.5, width 1.6/1.4; III: length 2.0/1.7, width 1.6/1.4; IV: length 2.3/1.9, width 1.5/1.3; V: length 3.8/3.3, width 1.4/1.2, depth 1.2/1.1. Telson length 3.5/3.2. Vesicle: width 1.6/1.2, depth 1.2/1.0. Pedipalp: femur length 4.9/3.3, width 1.4/1.3; patella length 5.2/3.6, width 1.5/1.5; chela length 8.8/6.8, width 2.2/2.5, depth 1.9/2.1; movable finger length 3.3/3.2.
Relationships: Only few species of Euscorpiops have previously been described from Vietnam, namely E. kaftani (Kovařík, 1993) and E. sejnai (Kovařík, 2000). Originally described in Scorpiops, these species were subsequently transferred to the genus Euscorpiops [7,17]. Their status remains obscure and comparison is hampered by their incomplete descriptions, without adequate illustrations of most characters. In addition, the type material is deposited in a private collection and is not available for study. The new species is geographically close to E. sejnai (Fig. 4), but it can be distinguished from the latter by several features:
- • a markedly smaller adult size and quite distinct morphometric values;
- • chela of pedipalp proportionally narrower, particularly in males;
- • paler coloration pattern;
- • less marked carinae and granulations;
- • only 17 trichobothria on the external aspect of the patella.

Map of Vietnam showing the type localities of Euscorpiops dakrong sp. n (black star), and related species, E. sejnai (black square), E. kaftani (black circle) and Scorpiops oligotrichus (black triangle).
5 Taxonomic remarks on Scorpiops oligotrichus Fage, 1933
Scorpiops oligotrichus was described by Fage [18] as a subspecies of Scorpiops montanus Karsch, 1879. The description of the new subspecies was extremely limited and based only on the number of patellar trichobothria. Fage [18] refers to three specimens collected in Annam: one female of length 25 mm, without any further precision of locality, one juvenile of length 16 mm, collected at Bana, at 300 m in altitude, and one juvenile of length 12 mm, collected at Djirinh, at an altitude of 1200 m on Pandarang Mountain. Later, Fage [19] promoted S. oligotrichus to the rank of species, based on the study of several new specimens, including adults reaching 32 to 33 mm in total length. Some confusion concerning the status of type material of this species existed in the collections of the Muséum in Paris, since Fage included the material listed in 1946 as part of the type material. However, only the three specimens originally listed by Fage [18] can be considered as types. We here designate the 25-mm-long pre-adult female as the lectotype and the two juveniles as paralectotypes.
The generic position of S. oligotrichus may, however, require reconsideration. The position of chelal trichobothrium Eb3 seems instead to associate this species with the genus Euscorpiops, but the presence of an annular ring on the telson does not appear to be clearly marked. Perhaps this is due to the fact that only juvenile and pre-adult specimens have been examined. A final decision on the generic position of this species must await the availability of fresh adult specimens.
6 Dakrong Nature Reserve
Dakrong Nature Reserve is located in the South of Dakrong district, Quang Tri Province (Fig. 4). In the North it is adjacent to Cam Lo district, in the South to A Luoi district (Thua Thien Hue Province), in the East to Phong Dien district (Thua Thien Hue Province), and in the West to Dakrong river and to the National Road 14B (Ho Chi Minh highway).
The Reserve has a total area of 37,640 ha, with a strict protection zone of 23,590 ha, an ecological rehabilitation zone of 13,409 ha, and an administration zone of 641 ha. The landscape is dominated by low mountains belonging to the Eastern Truong Son range. Its southeastern boundary is shared with Phong Dien NR and continues along the borders of the provinces of Quang Tri and Thua Thien Hue. The highest mountain peaks are Coc Muen Cave (1410 m) and Cacut Cave (1405 m). Here originate several streams of the Dakrong river watershed area. The altitude of the territory declines gradually along the Dakrong valley and towards the Ba Long delta. It is rich in mountains and valleys due to its history of tectonic and mountain-forming processes. The Dakrong NR area is noted for its cave systems (Figs. 5 and 6).

(Color online). A typical cave of the Dakrong Nature Reserve cave system. External view.

(Color online). A typical cave of the Dakrong Nature Reserve cave system. Interior view.
The continuous forest block that covers Phong Dien and Dakrong Nature Reserves, in Thua Thien Hue and Quang Tri Provinces, respectively, hold some of the region's rarest and most unusual species.
Dakrong NR is covered with primary and secondary vegetation. It belongs to the transition zone between the northern and southern regions of Central Truong Son. The typical vegetation is evergreen lowland rain forest (Fig. 7) with high biodiversity values in a special landscape. The flora is very diverse, with high numbers of species, including many gymnosperms. Its forests are important for the protection and maintenance of the watershed area, providing irrigation reservoirs for agriculture [20].

(Color online). Typical evergreen lowland rain forest vegetation of Dakrong Nature Reserve.
Disclosure of interest
The authors declare that they have no conflicts of interest concerning this article.
Acknowledgements
We are most grateful to Mark Judson (Muséum, Paris) for revising an earlier version of the text and to Élise-Anne Leguin (Muséum, Paris) for her contribution to the preparation of the photos and plates.