

1 Introduction
The determination of species boundaries, one of the main objectives of taxonomy, is important to evolutionary ecology and conservation ecology, because species remain the fundamental units and operational entities in most disciplines in these fields. Species misidentification and species confusion could lead to overestimating genetic diversity, biasing estimates of genetic differentiation between populations, overestimating densities, underestimating risks of local extinction, or producing meaningless estimates of demographic parameters. This in turn may misguide management actions. A common problem is that of cryptic species, undetected using traditional taxonomic approaches.
Cryptic species are defined as distinct evolutionary lineages with a substantial amount of genetic distinctiveness and no apparent morphological differences [1–3]. Highly divergent mitochondrial clades within a nominal species, where within-clade diversity is several times lower than divergence between clades might be caused by either secondary contact, or introgression following interspecific hybridization, or the occurrence of hitherto-unrecognized, “cryptic” species. The barcoding literature shows several examples of deep divergence at the mitochondrial cytochrome-oxidase 1 (CO1) locus within fish species, which have been ascribed to cryptic species (e.g., [4–12]). These examples thus illustrate the potential of mitochondrial sequences to flag putative new species in marine fishes.
The low level of morphometric variability and the poor phylogenetic information borne by the morpho-anatomical characters used so far in the systematics of the grey mullets (Actinopterygian fish family Mugilidae) have led to contradictory hence unreliable morphology-based phylogenies (reviewed in [13]). This emphasizes the need for molecular systematics in this family. Molecular phylogenetics has demonstrated the occurrence of distinct, deep, sometimes paraphyletic mitochondrial lineages in a proportion of species in the Mugilidae, pointing to the possible occurrence of cryptic species [13–15]. As a consequence, the species richness of the family Mugilidae is currently underestimated and possibly largely so. The species concept on which the present revision is based is the unified species concept of de Queiroz [16], which views species as separately evolving metapopulation lineages. Reciprocal monophyly and reproductive isolation are two of the relevant properties of species [16] one expects to observe or infer from molecular population genetic data. These two properties of species will be the focus of the present taxonomic review of the Mugilidae.
Based on the only comprehensive, mitochondrial phylogeny of species in the family Mugilidae available to date [13], the objectives of the present paper are:
- • to identify deeply divergent mitochondrial lineages that correspond to putative cryptic species in several mugilid genera;
- • to revise the current nomenclature of species by proposing new, provisional names to these lineages;
- • to provide molecular diagnoses to species and putative cryptic species.
Addressing these objectives is a necessary step to clarify the nomenclature of species in the Mugilidae, in a taxonomic context where genetic markers are replacing traditional morphological characters.
2 Materials and methods
2.1 Rationale of the present systematic revision
Durand et al.’s [13] mitochondrial phylogeny of the Mugilidae has uncovered a number of deeply divergent lineages within nominal species. Several of the lineages were paraphyletic with other species; other lineages represented reproductively isolated sympatric species, as demonstrated by genotypic frequencies at nuclear loci or inferred from karyotypes. Last, in some instances, deeply divergent sister-lineages characterized geographically separate populations within a species. Thus, there was substantial evidence for cryptic species in Mugilidae, based on the tree topology, on independent data from nuclear markers, and on the geographic distribution of sister lineages. We used W.N. Eschmeyer's fish database [17] as the reference for the current nomenclature. The current nomenclature was maintained for a lineage when its geographic distribution was compatible with the type locality of the species. By analogy with these cases where specific status was documented, we considered other deep lineages in Mugilidae, i.e. lineages whose distance to its nearest neighbour exceeded the gap between infra-specific and inter-specific pairwise distances (see section 2.5), to potentially represent additional cryptic species. We maintained the current nomenclature to designate those lineages that unambiguously correspond to the type material, based on the type locality, and we arbitrarily assigned capital letters to the other lineages. The other lineages were thus provisionally denominated “sp. A”, “sp. B”, etc.
We emphasize that our approach is not one of DNA barcoding, but one of molecular taxonomy, where molecular diagnoses of species and putative cryptic species are provided. We use gaps in the distributions of pairwise genetic distances as a means to distinguish deep lineages, which is where one may find analogy with barcoding. Nevertheless, the utility of CO1 barcoding for identifying species in the family Mugilidae will be the topic of a separate paper.
2.2 Genus nomenclature
In this paper, genus nomenclature accords with our recent revision [18], where the following changes have been made, relative to the previous nomenclature: Moolgarda seheli and Valamugil buchanani have been placed together with Crenimugil crenilabis under Crenimugil, and Moolgarda cunnesius, Moolgarda engeli, Moolgarda perusii, and Valamugil robustus have been placed under the resurrected genus Osteomugil; likewise, Liza aurata, Liza bandialensis, Liza dumerili, Liza ramada, Liza richardsonii, Liza saliens, and Liza tricuspidens have been placed together with Chelon labrosus under Chelon; likewise, Chelon macrolepis, Chelon melinopterus, Chelon subviridis, Liza abu, Liza affinis, Liza alata, and Liza haematocheila have been placed under the resurrected genus Planiliza; C. planiceps has since then been synonymized with Liza tade [17] and placed under Planiliza; also, Sicamugil cascasia, Agonostomus monticola, Liza argentea, Rhinomugil nasutus, and Oedalechilus labiosus have been placed, respectively, under the resurrected genera Minimugil, Dajaus, Gracilimugil, Squalomugil, and Plicomugil whereas Xenomugil thoburni has been placed under Mugil; the genus names Liza, Moolgarda, Valamugil and Xenomugil have been dismissed; three new genera have been erected: Neochelon (for Liza falcipinnis), Parachelon (for Liza grandisquamis), and Pseudomyxus (for Myxus capensis).
Durand et al. [18] have also synonymized the genus Paramugil [19] with Planiliza. We must acknowledge that this was an error as explained in the following. We erroneously used as reference specimen for Planiliza parmatus individual MNHN-IC-2011-0212, numbered 118 in [18], which had been collected in south Java by S. Kleinertz. On the basis of photographs that he kindly agreed to examine, H. Senou identified this specimen as a Planiliza (“Chelon”), and not a Paramugil. This specimen was subsequently examined by J. Ghasemzadeh who also rejected our identification as Paramugil and identified it as Planiliza (“Liza”) melinoptera based on its external morphological features.
2.3 Choice of a reference database
Durand et al.’s comprehensive mitochondrial phylogeny of the Mugilidae [13,18], which is based on the concatenated partial 16S rRNA, COI and cytochrome b gene sequences (3885 bp long in total) of 257 reference specimens (including 120 vouchers deposited in museum collections), was used for the present investigation. Zooms on regions of interest in this phylogeny are presented in Figs. 1 and 2.
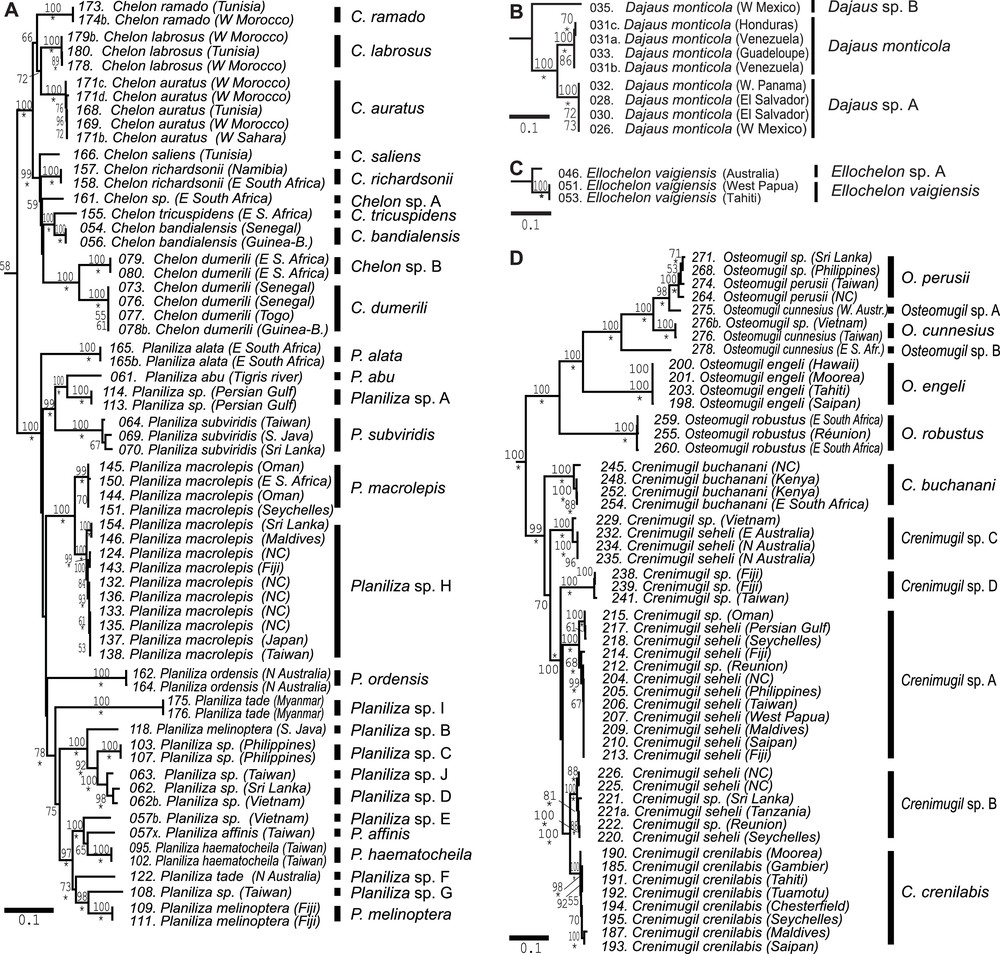
Phylogenetic trees depicting relationships among mugilid species, constructed from partitioned maximum-likelihood (ML) analysis of 3885 aligned nucleotides from 16S rRNA, COI and cytb gene sequences [13]. Vertical bars on the right of the tree indicate well-supported lineages that potentially represent distinct species. NC: New Caledonia. A. Among species within genera Chelon and Planiliza as redefined by [18]. B. Within genus Dajaus as redefined by [18]. C. Within genus Ellochelon. C. Among species within genera Osteomugil and Crenimugil as redefined by [18].
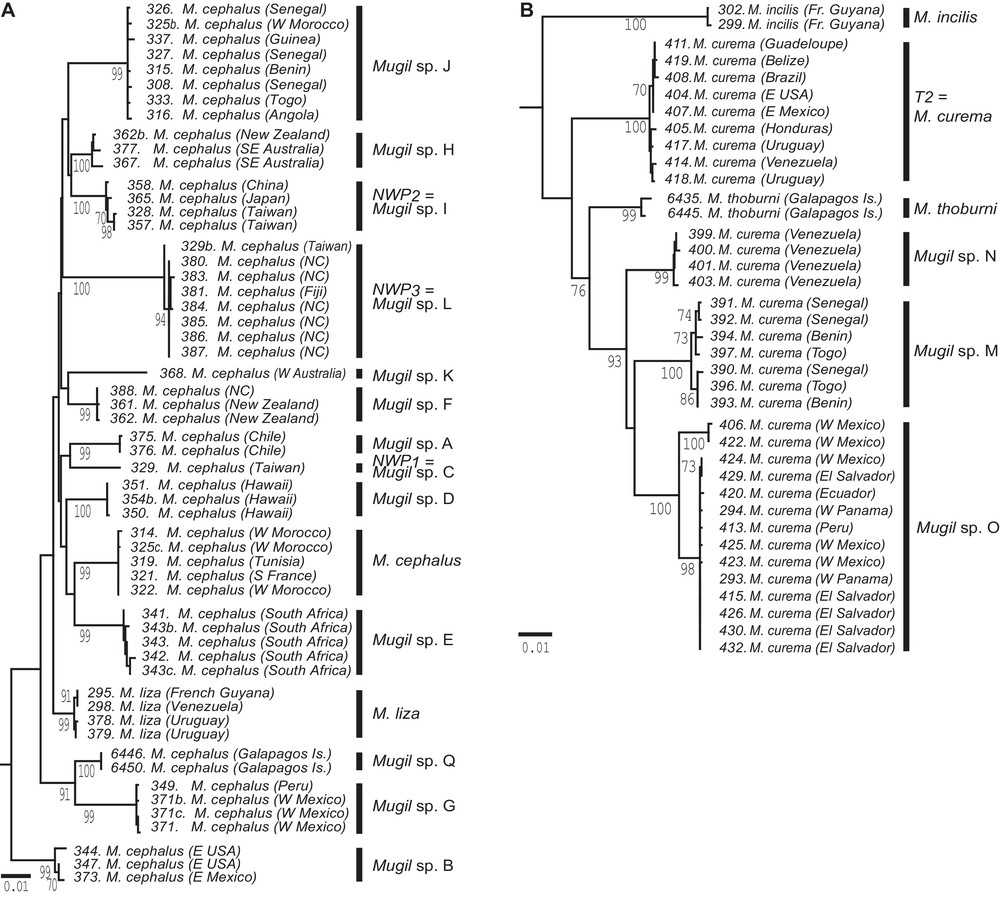
Phylogenetic trees of species in the genus Mugil, constructed from partitioned maximum-likelihood (ML) analysis of 3885 aligned nucleotides from 16S rRNA, COI and cytb gene sequences [13]. Bootstrap scores >50% are indicated. Vertical bars on the right of the tree indicate statistically supported lineages that potentially represent distinct species [13,18]. A. Tree of individual haplotypes in the Mugil cephalus species complex, including M. cephalus and Mugil liza. NWP1, NWP2, NWP3: lineages characteristic of three respective cryptic species (Mugil sp. C, Mugil sp. I, Mugil sp. L) sampled in the East China Sea [14]; the other lineages were assigned species names Mugil spp. A, B, D–J, and Q; the lineage sampled in the northeastern Atlantic and in the Mediterranean is the actual M. cephalus. NC: New Caledonia. B. Tree of individual haplotypes in the Mugil curema species complex.
2.4 Identification of within-genus gaps in nucleotide distance
Pairwise nucleotide distances between haplotypes sampled within each of seven mugilid genera (Chelon, Crenimugil, Dajaus, Ellochelon, Mugil, Osteomugil, Planiliza) were estimated under Mega5 [20] from the concatenated haplotype sequences at loci 16S rRNA, COI and cytb, which have been published previously [13,18]. Nucleotide distance was estimated according to the model of molecular evolution that, among the list of models proposed by Mega5, ranked as the most likely after the GTR-related model used to construct the phylogeny of [13], because the GTR model is not proposed by Mega5 for estimating nucleotide distances. The model thus chosen was the Tamura-Nei (TN93; [21]) with gamma distribution and invariable sites (+G + I) model. Nucleotide distances between lineages estimated according to the Kimura-2 parameter (K2P; [22]) model of molecular evolution were also presented. For each of the seven genera or species complexes we focused on, the resulting phylogenetic tree was examined together with the matrix of pairwise nucleotide distances between haplotypes. Our objective was first to determine the threshold below which distances all were infra-specific and above which they were all inter-specific. We then used this value as a yardstick to determine deep lineages that may represent cryptic species.
Further, alternative analysis of the dataset was done using the automatic gap determination algorithm proposed by N. Puillandre and co-authors to detect putative species from barcode datasets (ABGD; http://wwwabi.snv.jussieu.fr/public/abgd/; [23]). The analysis was run on each of the seven sequence datasets representing genera or species complexes, using the default settings of the program. This algorithm detects the gap in the distribution of pairwise nucleotide distances as the first significant gap beyond infra-specific distances and uses it to partition the dataset. Inference of the limit and gap detection are then recursively applied to previously obtained groups to get finer partitions until there is no further partitioning [23].
3 Results and discussion
3.1 Evidence of nucleotide-distance gaps within mugilid genera
Pairwise distributions of nucleotide distances among individuals within each of the seven genera focused on in the present paper are presented Fig. 3. Detailed examination of the distribution in the genus Mugil (Fig. 3E) revealed a gap after 1%: it is therefore sensible to consider the values ≤1% separately from the rest of the distribution and to ascribe them to genetic variation at the infra-specific level. This 1% threshold value also precisely coincided with the right boundary of the first mode of pairwise nucleotide distances in the genus Chelon (Fig. 3A), and it encompassed the homologous first modes in Dajaus (Fig. 3C) and Ellochelon (Fig. 3D). Detailed examination of intraspecific distances within C. crenilabis and its morphologically distinct sister-species Crenimugil sp. B, the two most closely related lineages in the genus Crenimugil (Fig. 1D), showed no infra-specific distance greater than 1.5%. Within the other Crenimugil lineages (Fig. 1D) the highest pairwise distance was 2.2% while the lowest inter-lineage distance was 3.4%. Similarly, in the genus Osteomugil a gap in pairwise distances occurred between 2.1% and 4.3%. In Planiliza, a similar, although narrower gap was observed between 2.1%, the highest distance found within P. subviridis, and 2.6%. Thus, placing a threshold at 1% allowed the delineation of the first mode of the distribution of pairwise nucleotide distances in four (Chelon, Dajaus, Ellochelon, and Mugil) (Fig. 3A, C–E) of the seven mugilid genera tested. In the three remaining genera (Crenimugil, Osteomugil, and Planiliza) (Fig. 3B, F, G), the threshold should be placed at 2.5% based on the gap in the distribution of pairwise nucleotide distances.
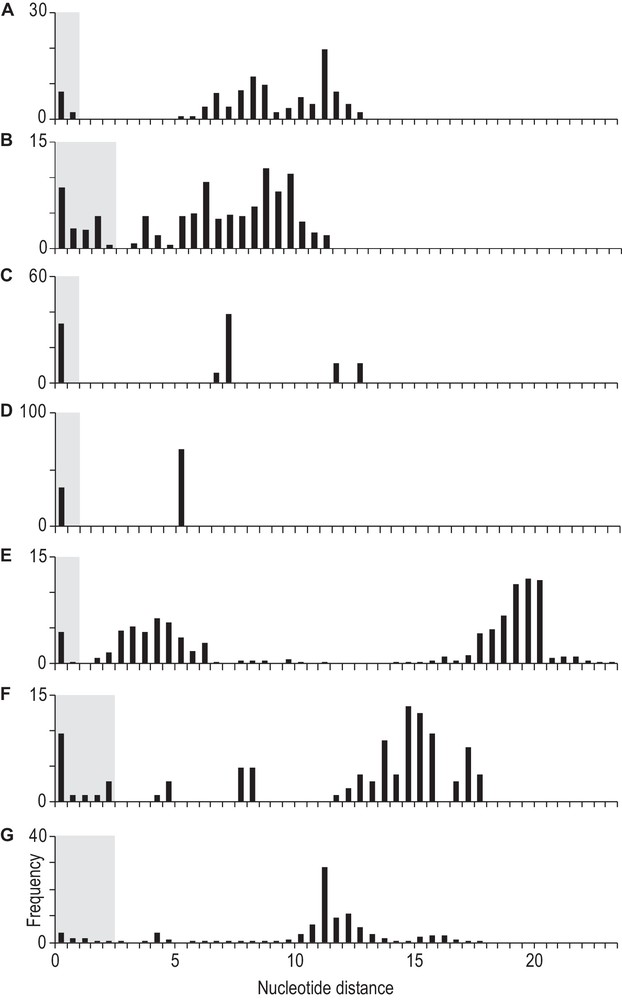
Frequency distribution of pairwise nucleotide distance estimates (TN93 + G + I model; Mega5 [20]) among individuals within each of 7 mugilid genera. Shaded rectangles highlight pairwise nucleotide distances ≤ 1% (in Chelon, Dajaus, Ellochelon, and Mugil) or ≤2.5% (in Crenimugil, Osteomugil, and Planiliza) within a deep lineage. A. Chelon. B. Crenimugil. C. Dajaus. D. Ellochelon. E. Mugil. F. Osteomugil. G. Planiliza.
3.2 Mitochondrial lineages that characterize cryptic species
In this section, we review all cases where lineages separated from the closest neighbouring lineage by a nucleotide distance larger than the threshold defined in the preceding section correspond to distinct species. In the mitochondrial phylogeny of the Mugilidae published by [13], Dajaus monticola consisted of three deeply rooted lineages (Fig. 1B), the two most recently diverged of which were geminate lineages distributed on either side of Central America, separated by 7.9% net nucleotide divergence under the K2P model and 7.0% under the TN93 + G + I model, all three markers combined. These two lineages have likely been geographically isolated from one the other for millions of years, hence are likely to represent separate species. The third lineage, from the eastern Pacific, branches externally to the two latter and is likely to represent another species. The type-locality of D. monticola is Jamaica [17]. Therefore, we here maintain the epithet monticola for the Atlantic lineage and provisionally designate the two other lineages, both from the eastern Pacific, as Dajaus sp. A and sp. B. The geographic distribution of the different lineages within D. monticola is presented in Fig. 4B. A subsequent study [15] estimated the divergence between the two geminate lineages D. monticola and Dajaus sp. A to be 31.8–11.8 million years old; it also evoked morphological differences between the two lineages from the eastern Pacific, confirming their status as separate species. The same study reported a fourth lineage currently under D. monticola from the Mexican rivers of the Gulf of Mexico, that is, geographically separated from what we here consider to be the true D. monticola [15].
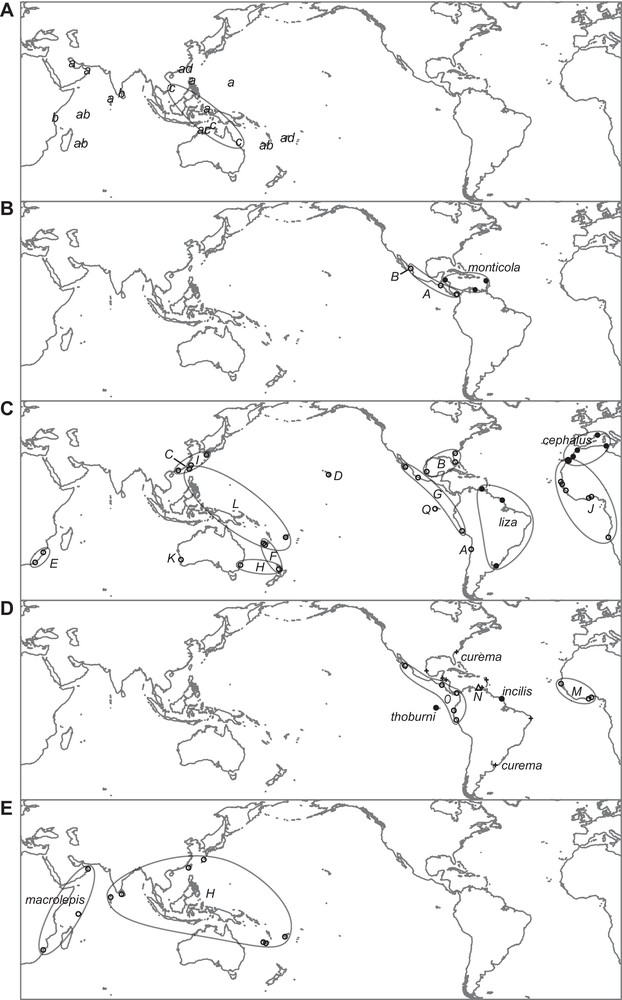
Geographic distribution of mitochondrial lineages (putative species) in grey mullets, based on the sampling of [13,18] (Figs. 1 and 2). Background map of the Indo-West Pacific was obtained from Digital Vector Maps, San Diego (http://digital-vector-maps.com/). A. Lineages of the Crenimugil seheli species complex. a–c, putative cryptic species Crenimugil spp. A to C, respectively; doted ellipse encompasses all known locations where Crenimugil sp. C occurs; d, Crenimugil sp. D. B. Lineages of Dajaus monticola. A, B: putative cryptic species Dajaus spp. A and B, respectively; all three samples from the Pacific coast of Central America included Dajaus sp. A. C. Lineages of the Mugil cephalus species complex. A-L, Q: putative cryptic species Mugil spp. A to L and Q, respectively. D. Lineages of the Mugil curema species complex. Crosses (+): M. curema; M–O: putative cryptic species Mugil spp. M to O, respectively. E. Lineages of Planiliza macrolepis. H, putative cryptic species Planiliza sp. H.
The mitochondrial phylogeny of the M. cephalus/M. liza species complex (Fig. 2A) revealed 15 separate lineages, each with deep (>1%) rooting and shallow (<1%) within-lineage diversity. Three of these lineages, which occur in sympatry in Taiwan, belong to genetically distinct forms reproductively isolated from one another as demonstrated by their distinct composition at nuclear loci and by the quasi-absence of hybrids [14]. These lineages, coined NWP1-3 by [14], show 3.2%–4.8% nucleotide divergence under the K2P model, all three markers combined, and 3.3%–5.1% divergence under the TN93 + G + I model [20]. The three lineages are here assigned arbitrary species names sp. C, sp. I and sp. L., respectively (Fig. 2A). Mugil curema similarly consisted of a complex of species, where deeply rooted lineages were paraphyletic with another species, Mugil thoburni [18] (Fig. 2B). The type locality of M. curema is Bahia, Brazil [24] where only one lineage, also characterized by a chromosome complement number of 2n = 28, is present [13]. This is the “Type 2” (T2) karyological form of [25]. The topology of the tree (Fig. 2B) shows lineage T2 to root externally to the sub-clade consisting of M. thoburni and the other M. curema lineages. We here keep the name M. curema exclusively for lineage T2 and we designate the other lineages as Mugil sp. M to sp. O. The case of the three Mugil sp. M to sp. O lineages will be discussed in the following section.
3.3 Recognizing deeply divergent lineages as putative new species
A number of deeply divergent lineages potentially represent additional cryptic species in the Mugilidae. These cases are examined genus by genus in the following, where each lineage either was assigned a capital letter, or conserved its current name.
In the genus Chelon (Fig. 1A), Durand et al. [13] had sampled an unidentified Chelon sp. lineage from southeastern Africa (their specimen no. 161), which is here provisionally designated as Chelon sp. A. In the same genus, the haplogroup corresponding to Chelon dumerili actually comprised two distinct lineages separated by a net nucleotide distance, all three markers combined, of 7.5% (under the K2P model) or 8.1% (under the TN93 + G + I model). One lineage was exclusively sampled in western Africa, including Saint-Louis, Senegal [13], which is the type locality of the species [26]. We maintain epithet dumerili for this lineage. The other lineage was sampled exclusively in southeastern Africa and is here provisionally referred to as Chelon sp. B. We consider Chelon sp. B to be putatively a species distinct from C. dumerili based on the disjunction in geographic distributions and the level of nucleotide distance between the two lineages.
In the genus Crenimugil (Fig. 1D), three distinct lineages were observed within C. seheli under its current definition. These three lineages, which occur sympatrically in the Indo-West Pacific, are separated by a net nucleotide divergence, all three markers combined, of 4.5%–7.8% under the K2P model and 4.8%–8.6% under the TN93 + G + I model, whereas intraspecific nucleotide diversity under both models was ≤2.2%. The three lineages were paraphyletic with C. crenilabis and with an undescribed Crenimugil species sampled from Taiwan and Fiji, represented by individuals Nos. 238, 239 and 241 of [13]. Therefore, we consider them to characterize putative, distinct species, here designated as Crenimugil spp. A-C. The undescribed Crenimugil sp. species from Taiwan and Fiji is here designated as Crenimugil sp. D. Fig. 4A presents the geographic distribution of all four deep lineages within the C. seheli/C. crenilabis species complex.
In the genus Ellochelon (Fig. 1C), two separate lineages were observed, which diverged by 4.8% net nucleotide distance under the K2P model and 5.1% under the TN93 + G + I model, all three markers combined. One lineage included specimens from Waigeo, the type-locality [27] and French Polynesia, and another lineage was represented by a specimen from an unknown location in Australia. Epithet vaigiensis is here provisionally retained for the Ellochelon lineage sampled in Waigeo while the other lineage is provisionally assigned putative species name Ellochelon sp. A.
In the genus Mugil, the 13 distinct lineages originally uncovered within M. cephalus belonged to the same sub-clade as M. liza [13]. The average ± SD net nucleotide distance between lineages, from which M. liza was excluded was, all three markers combined, 3.6% ± 1% under the K2P model and 3.8% ± 1% under the TN93 + G + I model. Subsequently, a fourteenth lineage comprising haplotypes sampled from the Galapagos Islands was reported [18] (Fig. 2A). Three of the lineages currently within M. cephalus (i.e., sp. C, sp. I and sp. L), which occur in sympatry, belong to genetically distinct forms reproductively isolated from one another (see preceding section). Hence, basing our analogy on similarity in ratios of inter- to intra-lineage nucleotide distance, and also taking into account the current taxonomic standards that designate M. liza as a species separate from M. cephalus, we consider all other 11 lineages within M. cephalus under its current definition to be putative, distinct species. One notes that these lineages apparently have allopatric or parapatric distributions (Fig. 4C). The original description of M. cephalus [28] geographically refers to a species which “habitat Oceano Europeo”. Accordingly, we can designate without ambiguity the only lineage sampled in the Mediterranean Sea [13,29] as characterizing the actual M. cephalus. The 10 remaining lineages are here provisionally assigned putative species names Mugil sp. A, sp. B, spp. D-H, sp. J, sp. K and sp. Q. Fig. 4C presents the geographic distribution of the 15 deeply rooted lineages within the M. cephalus species complex (i.e., M. cephalus, M. liza, cryptic species Mugil spp. C, I, L, and putative cryptic species Mugil spp. A, B, D-H, J, K, Q). Three other Mugil spp. lineages within the species initially designated as M. curema were uncovered (see preceding section). Mugil sp. N and Mugil sp. O differ from M. curema by their karyotypes (respectively, 2n = 24 and 48) indicating that they are likely reproductively isolated from the latter as well as from one the other [13]. Mugil sp. M, from the Pacific Ocean, is paraphyletic with Mugil sp. N and Mugil sp. O, both from the Atlantic Ocean (Fig. 2B). These lineages differ by 3.2%–5.4% net nucleotide distance under the K2P model and 3.4–5.8% under the T93 + G + I model, all three markers combined. In comparison, nucleotide diversities within a lineage ranged from 0.2% to 0.5% (under both models). Hence, we consider the three lineages to represent distinct species. Still in Mugil, a lineage originally assigned to Mugil hospes by [13] was represented exclusively by haplotypes sampled from the Gulf of Mexico. Because the type-locality of M. hospes is Mazatlan in the eastern Pacific [30], it is sensible to provisionally assign the lineage from the Gulf of Mexico to a yet undetermined Mugil species, here designated as Mugil sp. R. Last, Mugil rubrioculus comprises two distinct lineages, one sampled from Venezuela, the type locality of the species [31], and the other one from the eastern Pacific [13]. The lineage from Venezuela retains epithet rubrioculus while the eastern Pacific lineage is here provisionally designated Mugil sp. P. This lineage may represent the same species as M. aff. rubrioculus previously mentioned from the eastern Pacific [31]. Fig. 4D presents the geographic distribution of the 7 deep lineages (i.e., M. curema, Mugil incilis, M. thoburni, and putative cryptic species Mugil spp. M-P) within the M. curema species complex.
In the genus Osteomugil (Fig. 1D), three distinct lineages representing Osteomugil cunnesius under its current definition have been found to be paraphyletic with Osteomugil perusii [13]. The type locality of O. cunnesius is the Moluccas [24]. Hence, the lineage sampled in Taiwan and in Vietnam by [13], geographically closest to the Moluccas, is provisionally retained as the actual O. cunnesius while the two other lineages, one from eastern Western Australia, the other one from South Africa, are here assigned provisional names Osteomugil sp. A and Osteomugil sp. B, respectively.
In the genus Planiliza (Fig. 1A), both Planiliza melinoptera and Planiliza tade were found to be polyphyletic ([13]; present work). Individual 118 of [13], which was initially, erroneously identified as Planiliza parmata is now recognized as a cryptic lineage of Planiliza melinoptera (see section 2.2). This lineage was separated from the P. melinoptera sampled in Fiji (individuals 109 and 111 of [13]) by 10.3% nucleotide distance (under the K2P model) and 11.4% (under the TN93 + G + I model), all three markers combined. We kept the Fiji specimens under P. melinoptera, because of the geographic proximity of Fiji with Vanikoro, the type locality [24] and we assigned provisional species name Planiliza sp. B to the lineage sampled in South Java. One of the two P. tade lineages concerned specimens sampled in Myanmar; the other lineage was sampled in northern Australia. These two regions being remote from the Red Sea, the type-locality of P. tade [32], we here followed a cautious line by designating as Planiliza sp. F the lineage from northern Australia, and Planiliza sp. I the lineage from Myanmar. Two sister-lineages were observed within P. macrolepis, separated by 3.5% (under the K2P model) or 3.7% (under the T93 + G + I model) net nucleotide divergence, all three markers combined: one lineage was exclusive to the western Indian Ocean west of the Seychelles Islands, including the Seychelles Islands and including South Africa, the type-locality [33] while the other lineage had a wide geographic distribution, as it consisted of all haplotypes sampled east of the Seychelles Islands, from the Maldives Islands to Fiji [13]. The latter is here provisionally designated as Planiliza sp. H, while its sister lineage retains the epithet macrolepis. Fig. 4E presents the geographic distribution of the two deep lineages within P. macrolepis (i.e., the actual P. macrolepis, and Planiliza sp. H). A lineage represented by a single individual from Taiwan (no. 108, “Planiliza sp.” of [18]) separated from its sister-lineage, P. melinoptera, by 7.2% nucleotide distance (under the K2P model) and 7.8% (under the TN93 + G + I model), all three markers combined, is here provisionally designated as Planiliza sp. G. Five other undetermined Planiliza species were here assigned provisional species names Planiliza sp. A (including individuals nos. 113 and 114 of [13]), Planiliza sp. C (nos. 103, 107), Planiliza sp. D (nos. 062, 062b), Planiliza sp. E (no. 057b), and Planiliza sp. J (no. 063). Our distinguishing Planiliza sp. J from its sister-lineage Planiliza sp. D is justified by the distance between the two lineages (2.6–2.8%, above the 2.5% threshold set for the genus).
Through automatic gap determination using the ABGD algorithm of [23], the present sequence dataset was found to conceal 10 separate lineages in genus Chelon, 6 in Crenimugil, 3 in Dajaus, 15 in the M. cephalus species complex, 6 in the M. curema species complex, 6 in Osteomugil, and 17 in Planiliza. The lineages designated by ABGD were all identical to those reported on Figs. 1 and 2, except for one lineage in Planiliza (Planiliza sp. J) that escaped detection using the default settings of ABGD.
3.4 Molecular diagnoses of species
The present results lead us to propose molecular diagnoses for a number of mugilid species currently considered as valid [17]. We aligned the partial sequences of the specimens characterized at the three loci (i.e., 16S, CO1 and cytb) used as phylogenetic markers [13,18] in each of 7 cases treated in the present study (i.e., Chelon spp., Crenimugil spp., Dajaus spp., species in the Mugil cephalus species complex, species in the M. curema species complex, Osteomugil spp. and Planiliza spp.). Ellochelon spp. was excluded because it consisted of two main lineages only, one of which was represented by a single individual in our dataset, thus insufficient for a comparison of inter-lineage vs intra-lineage variation. Variable nucleotide sites in each alignment were highlighted (Supplementary material, Tables S1–S21). Nucleotide sites diagnostic of species were determined. This information is summarized in Table 1. For example, M. cephalus L. is here diagnosed relative to the other species in the M. cephalus species complex by triplets (T234, T435, C693) at locus CO1 and (T183, T483, G510) at locus cytb, where nucleotide sites are numbered from the start of the gene, using the mitochondrial DNA sequence of Mugil sp. C (GenBank no. AP002930) as reference. Anonymous lineages designated by alphabetical letters were similarly diagnosed (Table 1). No molecular diagnosis was proposed for those species for which a single specimen was available: Chelon saliens, C. tricuspidens, Osteomugil cunnesius, and Planiliza abu. Similarly, no diagnosis was proposed for lineages Chelon sp. A, Dajaus sp. B, Mugil spp. C, K, Osteomugil sp. A, and Planiliza spp. E-G.
Nucleotide diagnostic of a lineage in the genera Chelon, Crenimugil, Dajaus, Osteomugil and Planiliza, and in the Mugil cephalus and M. curema species complexes. Based on the individual sequence data presented in Supplementary material, Tables S1–S7 (locus 16S), Tables S8–S14 (locus CO1), and Tables S15–S21 (locus cytb).
| Genus/species complex, | Locus | ||
| Lineage | 16S | CO1 | cytb |
| Chelon (JF911706) | |||
| C. auratus | T1099 C1228 C1299 | G129 T244 A247 T249 T267 A579 G591 G675 | G240 T279 G387 T540 T580 A585 G699 C783 |
| C. bandialensis | – | T393 C582 T666 | T264 G303 T321 C381 T399 C426 T630 G714 C774 |
| C. dumerili | A988 T1141 G1176 A1191 G1193 T1196 C1217 G1223 C1246 C1275 [T] G1296 [A] A1297 C1306 C1323 T1348 T1396 | T114 G237 G324 C414 C609 | C136 G221 A472 G498 G573 G576 T621 T625 T710 T717 T753 G771 |
| C. labrosus | T1096 T1237 | C132 T264 A276 T327 C552 G690 | T232 T396 T561 T585 T597 C603 T669 G681 G690 |
| C. ramado | – | T282 T336 C337 G396 T444 T450 T468 T630 G666 T699 | T102 A108 C153 T237 C243 T303 T345 G471 T573 T624 G645 G753 A795 |
| C. richardsonii | T1098 T1138 G1218 | A231 A279 G360 G477 A624 | T444 T476 A550 G609 G635 G641 T698 G795 |
| Chelon sp. B | G1021 C1066 C1095 G1181 T1183 C1192 [ATC] T1203 C1234 G1270 G1290 [C] | C114 G228 A258 T273 T279 C285 C366 C390 G456 G672 T687 | T129 A136 T138 G213 T216 C219 C245 A447 A453 C498 A501 T543 T612 T660 T715 T798 |
| Crenimugil (JF911707) | |||
| C. buchanani | T1174 T1199 T1236 | T123 C270 C337 C381 A399 T447 C453 G465 G471 C507 G534 C555 T570 C609 T618 T642 | C135 C261 A306 G339 A363 T420 G471 C507 T564 T567 T573 C580 T582 A583 T654 T655 C690 T693 C696 C714 G753 C789 T862 T873 |
| C. crenilabis | T1340 | C159 A348 G423 C486 G498 C654 | A177 T315 T324 A483 T597 A657 |
| Crenimugil sp. A | C1115 G1132 T1215 | G129 C267 C279 T303 C366 G408 C525 T699 | T273 C306 G645 C747 |
| Crenimugil sp. B | T1135 | C246 T264 T270 G543 | C369 T612 C804 |
| Crenimugil sp. C | C1203 C1298 T1364 | T174 A177 A246 T345 T384 C411 T414 T438 T454 A456 G678 | C138 G159 C171 T180 T216 A258 T267 A324 C345 A429 C438 T456 C477 T522 C528 A627 A630 T678 A693 T705 T765 G771 G789 C867 |
| Crenimugil sp. D | T988 C992 A1010 C1043 C1063 T1066 G1069 G1175 G1205 T1207 C1221 T1226 [CA] A1298 T1315 [A] A1359 T1369 A1480 | T111 T121 T234 T288 C372 G411 C433 T465 C531 T576 G585 A603 A612 C669 T700 | T225 T378 C390 C429 C453 T462 A472 A558 T784 T858 |
| Dajaus (JF911702) | |||
| D. monticola | C993 T994 G1138 G1139 T1300 T1319 G1331 T1503 | C141 C162 C219 T252 C264 C282 C297 C306 T310 C315 T336 C351 G378 A408 G435 T447 C450 G516 A534 G558 C565 C660 C687 G690 G696 | T105 C108 T120 G144 T165 A178 C183 T204 T225 T243 C288 T303 T322 C345 G351 T384 T429 T459 G489 T522 T537 T555 G576 T621 T627 T669 A702 A718 A750 A783 T784 A798 T810 T834 C837 T846 T852 |
| Dajaus sp. A | T990 G1129 G1165 T1228 A1300 | G129 G213 A222 A247 T249 C336 T351 T381 T435 T441 C447 C483 T528 A582 C591 T615 C636 T643 C654 G669 | T102 G108 T117 A123 T138 A144 A171 T288 C303 T326 G343 T378 C426 G457 G471 T492 A498 C522 C528 C594 C597 T672 T715 C730 C741 T774 G783 G789 T795 C798 C804 G807 T840 T843 |
| Mugil cephalus species complex (AP002930) | |||
| M. cephalus | – | T234 T435 C693 | T183 T483 G510 |
| M. liza | – | - | G580 T657 C843 |
| Mugil sp. A | – | G309 C312 A603 | G582 A627 C653 A711 |
| Mugil sp. B | T1223 | C219 G357 G462 T611 T660 | C384 C537 T708 A789 G831 |
| Mugil sp. D | – | G267 C270 | T318 T627 C719 C792 G849 |
| Mugil sp. E | – | A390 G579 | T516 T612 A637 |
| Mugil sp. F | – | – | T492 C876 |
| Mugil sp. G | T1208 T1244 | A438 | G219 C426 T444 T465 T507 T717 G810 G874 |
| Mugil sp. H | G1216 A1218 | – | C468 |
| Mugil sp. I | T1216 | A611 | C300 T357 T777 |
| Mugil sp. J | C1240 C1314 | C342 | G246 T312 T322 T636 |
| Mugil sp. L | G1106 T1189 T1205 A1238 A1294 T1311 | C210 C294 A330 T337 A348 G366 G393 T399 G400 A480 C510 T663 | T237 C288 G333 T342 G522 T540 T600 T756 T765 T768 |
| Mugil sp. Q | – | T552 C555 G597 | T346 G390 A426 C738 |
| M. curema species complex (JF911710) | |||
| M. curema | G1180 C1232 C1279 | C117 T153 A216 C270 T394 T411 C475 | T121 T150 G219 C279 C318 T324 T357 G387 C426 T585 T678 C685 T713 C852 |
| M. incilis | A979 T985 A993 T1029 C1056 G1082 G1139 A1191 T1194 T1200 C1202 T1206 G1209 A1214 A1215 C1218 G1224 T1226 C1233 T1234 [TATTTT] T1297 G1298 T1312 | C108 T135 G144 G246 G339 T390 A429 C483 C504 C525 A534 C555 G606 T609 C642 A690 C700 | C111 C153 C207 T216 G231 C286 G318 G331 A369 T399 G411 C429 T477 G501 T504 C537 T564 C567 G570 A585 A591 G609 T648 G681 C709 T724 T750 C810 C837 T862 |
| Mugil sp. M | C1138 T1361 | G105 G264 A390 T492 | T225 T684 T687 T765 T846 |
| Mugil sp. N | – | C142 T313 A420 T441 C552 | T183 G234 T336 C396 A522 C555 T627 T819 |
| Mugil sp. O | T1198 A1211 T1320 | T183 G477 C579 G591 T604 | T138 G159 T201 C204 T285 T840 |
| M. thoburni | A1174 | C174 A228 C429 G639 T654 | G127 T129 T258 C561 G634 T663 T675 C747 |
| Osteomugil (JF911717) | |||
| O. cunnesius | G1213 | G168 G339 T360 C366 T591 C606 G615 T663 G672 G684 | G117 C137 C291 T306 C369 G447 T498 A519 T576 G579 T630 T684 A810 G831 T855 |
| O. engeli | T998 C1069 T1086 T1100 T1119 T1142 C1143 G1198 C1203 T1206 T1220 A1232 A1270 G1272 C1277 A1306 A1326 C1342 T1352 | C105 T117 G120 G123 C141 A177 C246 C312 T337 C393 C435 T441 T498 T555 G558 A564 A567 C621 G633 A666 T675 | G114 A123 A147 G168 C198 T201 A228 G274 G373 C374 A387 C429 G441 G477 C580 T645 T648 T685 C705 T708 A723 T724 A753 T770 A864 T873 |
| O. perusii | T1307 | T126 A285 T318 G408 T447 C525 G540 G630 T678 | T363 T364 T445 A483 G582 C700 |
| O. robustus | C984 T990 C993 C1074 T1087 C1120 A1141 T1143 C1182 G1189 T1204 G1220 C1227 T1233 C1274 T1291 C1310 G1345 C1351 [A] T1430 A1438 | A123 C135 A136 T150 G291 A333 C360 A369 G378 T387 G435 C453 T462 A468 C480 T543 C549 G639 C654 T690 | C136 C147 T162 C168 A174 T178 G180 A213 A234 C288 T326 G387 C390 C399 A417 T429 C458 G531 A574 A583 T588 G589 C591 T607 C635 T636 A655 T660 A688 T697 T698 T712 T713 T719 C723 C724 A736 T742 T747 C789 T841 |
| Planiliza (JF911709) | |||
| P. affinis | – | A315 G585 | G114 A475 T546 G714 G729 T777 |
| P. alata | G1397 T1336 | T595 | T115 C137 T144 A178 C265 G300 A355 C367 C458 T526 C528 T543 C562 G592 C596 T606 C693 G700 G709 T719 A738 A751 G778 A846 |
| P. haematocheila | – | T394 | T276 G426 T612 T873 |
| P. macrolepis | – | G243 T363 | G234 T823 C879 |
| P. melinoptera | – | T147 A450 A594 | C357 T420 C867 |
| P. ordensis | A1198 G1263 C1265 | C108 G132 A249 A312 A543 G576 T630 G672 | C136 A168 C207 A240 A258 C438 G489 T510 G577 T578 C595 A712 T858 |
| Planiliza sp. A | A1068 | G372 T477 A513 A612 C672 | T102 T303 A462 T570 A579 T624 |
| Planiliza sp. C | T1041 C1042 C1092 G1213 G1398 A1473 | C138 G169 G300 C354 | G156 T384 G627 T660 C729 G774 G777 |
| Planiliza sp. D | – | – | T582 G681 |
| Planiliza sp. H | – | T267 | – |
| Planiliza sp. I | C978 A984 C988 T992 G1018 C1040 A1063 G1069 C1078 T1096 C1115 T1138 C1183 C1186 C1187 C1188 [CAA] C1195 G1196 A1199 C1201 [TCAT] T1219 C1223 C1237 T1252 A1258 A1261 C1263 G1271 T1275 G1296 [TAC] A1305 A1308 A1309 G1333 | T168 T189 T228 A330 A349 C350 A381 A390 T396 G438 A465 C474 T531 | T182 A288 G318 G405 A429 T603 T684 T706 A721 G732 A771 T822 T855 T864 |
| P. subviridis | T1028 T1136 T1197 A1371 | T142 G177 T360 G459 G462 C480 T525 G630 G690 | T114 T121 C574 T770 |
We are aware that future additional samples may lead to restricting the number of diagnostic sites for any given species relative to the other species in a genus. This is most likely to occur if additional cryptic species are sampled. However, the information in Table 1 may still provide the basis to future identification keys.
4 Conclusion
The morphological features that delineate species in the family Mugilidae [34] are insufficient to describe its actual species diversity. This was documented in Dajaus monticola, where two sister lineages are geographically isolated from one the other by a continent [13,15], in Mugil cephalus from the South and East China Seas where the three lineages present characterize reproductively isolated species [14], and in M. curema where distinct lineages are characterized by distinct karyotypes [13]. The mitochondrial phylogeny of [13,18] reveals an additional proportion of deeply rooted lineages that by analogy with the foregoing, flag as many additional putative cryptic species.
Future population genetic investigations based on nuclear markers are expected to provide clues to the degree of reproductive isolation between the populations harbouring separate mitochondrial lineages, in the cases where populations are sympatric or parapatric. In the case of allopatric lineages, reproductive isolation cannot be tested directly, hence additional lines of evidence would be necessary to distinguish species (e.g., [10,35]). Pending possible confirmation that the deeply divergent lineages listed in this paper are cryptic species, we anticipate changes to the current species nomenclature of the Mugilidae. Although new species descriptions might eventually be necessary in some cases, it will be first necessary to evaluate the validity of available names formerly assigned to a proportion of the lineages and subsequently considered junior synonyms. Epithets to be considered a priori for possible resurrection should be based on geography, i.e. by ensuring that the type-locality lies within the geographic range of the lineage, and on chronological priority [36].
Molecular genetic surveys of species in the Mugilidae may help uncover additional deep lineages. DNA-barcoding surveys potentially represent such opportunities [37]. For this purpose, it will be first necessary to evaluate the ability of the COI fragment used as barcode, to identify deep lineages that represent species or potential cryptic species in the Mugilidae.
Disclosure of interest
The authors declare that they have no conflicts of interest concerning this article.
Acknowledgments
We are grateful to H. Senou and J. Ghasemzadeh for kindly accepting to examine specimen MNHN-IC-2011-0212 (or photographs of it) and concluding in the rejection of our previous identification of it as Paramugil parmatus. We therefore acknowledge our error in two previous papers [13,18]. Many references from the taxonomic literature have been compiled by R. Fricke (http://research.calacademy.org/research/ichthyology/catalog/fishcatmain.asp). Pre-twentieth century books and articles were consulted online from the Biodiversity Heritage Library website (http://www.biodiversitylibrary.org/). Our background map of the Indo-West Pacific was edited from images downloaded from Digital Vector Maps, San Diego (http://digital-vector-maps.com/). PB, JDD designed the study; PB, JDD analysed and interpreted the data; PB wrote the paper.