

1 Introduction
In one of his contributions to the Arachnida of France, Simon [1] dedicated a large section to the order Scorpiones. In this section, he dealt with all the species known or reported from France and also described a few new ones. Among these, he described a new genus Belisarius and a new species Belisarius xambeui (Fig. 1) discovered in the southeastern region of France, more precisely in the Pyrenées-Orientales department. Curiously, he placed this new genus and species as well as the genus Euscorpius Thorell, 1876 in the family Ischnuridae.

(Colour online.) Belisarius xambeui, female from French Oriental Pyreneans with second-instar juveniles.
Photo courtesy E. Ythier.
The description of the new genus and species [1] was quite vague about the number and the sex of the described specimens. In fact, he referred to a male, as the only known sex. Besides, in several characters, in particular the few morphometric values presented, seemed to be associated with a single specimen. Simon [1] did not provide any type of illustration along with his description, which was a common feature of his publications. Subsequently, Borelli [2] clarified the characteristics of both sexes in B. xambeui and clearly stated that the specimens used by Simon [1] for his description were females.
In a more accurate redescription of B. xambeui, Vachon [3] suggested that Borelli [2] examined the material used by Simon [1] for the description of the new species; however, this is not certain. In this same publication, Vachon [3] stated that most previous publications about B. xambeui were merely new citations or new records, and none of these, including the one by Borelli [2], presented illustrations for the species. This contribution by Vachon [3] brought a supplement of confusion about the type material studied by Simon [1]. Vachon (1945) literally lists 11 females and two male juveniles as the type material used by Simon [1]. This probably influenced some future publications that totally misinterpreted these data. In fact, in his list of material, Vachon [3] took into consideration all the specimens of B. xambeui, which were available in the collections of the Museum in Paris by that date. Besides, he referred to several specimens collected in localities quite different from the one cited by Simon [1] as the type locality, and several specimens cited in his list have been collected many years after the date of the original description. In their Catalog of the Scorpions of the World, Fet and Sissom [4] compiled the list proposed by Vachon [3], and once again cited one female holotype as lost, and two male juveniles and ten adult females as paratypes.
Recently, I started an intensive investigation on both the collections of the Natural History Museum in Paris, and also on the private notes of Simon. These led to a number of clarifications. According to the personal notes of Simon, only the specimen registered under the number “Simon No. 2675” was considered as the type of B. xambeui. Vachon [3] listed No. 2675 as containing two adult females, and this was precisely what I found in the collections of the Museum in Paris. Of the two females registered under No. 2675, only one corresponds precisely to the specimen described by Simon [1], in particular to the morphometric values he provided in the original description. This specimen is now considered as the holotype of B. xambeui. It is difficult to affirm that the second female contained in the vial labelled No. 2675 is also part of the original material described by Simon [1], since this author used to mixed specimens of a given species after the original publication. Nevertheless, I will assume, a priori, that the second female may be considered as a paratype.
While investigating on the Belisarius material, I also located another interesting specimen registered as RS-7748 and collected by J. Malhomme. This collector did intensive collections in Morocco during the 1940/1950s, and this lead Vachon (determined from personal notes) to believe that the specimen was collected in this country. However, more accurate unpublished notes by Vachon do confirm that J. Malhomme also collected in Spain, and that the Belisarius specimen was in fact collected in the “Sierra de las Nieves” in the south of Spain. This new record for the genus Belisarius changes the known range of distribution (see Biogeographic section) and attests to the existence of a new geographical vicariant species, which is described here.
2 Methods
Illustrations and measurements were made with the aid of a Wild M5 stereo-microscope equipped with a drawing tube (camera lucida) and an ocular micrometre. Measurements follow Stahnke [5] and are given in mm. Trichobothrial notations follow Vachon [6] and morphological terminology mostly follows Hjelle [7].
3 Taxonomic treatment
Family TROGLOTAYOSICIDAE Lourenço, 1998
Subfamily BELISARIINAE Lourenço, 1998
Genus Belisarius Simon, 1879
Belisarius xambeui Simon, 1879
One adult female holotype (Fig. 2), one adult female paratype, France, Pyrénées-Orientales, Territoire de Conat, Vallée de Queillan, under stones, collected in April (year unknown) by Monsieur Xambeu.

(Colour online.) Belisarius xambeui, female holotype, habitus. A–B. Dorsal and ventral aspects.
Revised diagnosis based on the female holotype and female paratype. Moderate in size with the female holotype being 26.5 mm in total length. Coloration (the specimens are partially faded in alcohol) reddish to reddish–yellow, except for the venter and legs that are yellow. Pedipalps reddish; chela darker than patella and femur; granulations on the cutting edge of fingers blackish. Body and appendages weakly granulated or smooth; carapace and tergites with minute punctations. Anterior margin of carapace with a weakly marked concavity; median eyes absent; lateral eyes represented by a single macula, without lens (Fig. 3). Pectines with 4–4 teeth. Metasomal segment V with weakly marked spinoid granulations on ventral aspect; vesicle smooth, globular and flattened dorso-ventrally; aculeus short and strongly curved; annular-ring is moderately marked (Fig. 3). Dentate margins on movable and fixed fingers with a median denticle row composed of 5–6 groups of granules. Chelicerae with the dentition characteristic of the genus Belisarius and of the family Troglotayosicidae [3,8,9]; movable finger with the presence of a serrula [9]. Trichobothrial pattern type C orthobothriotaxic. Chela with four ventral trichobothria; patella with three ventral and 13 external trichobothria.
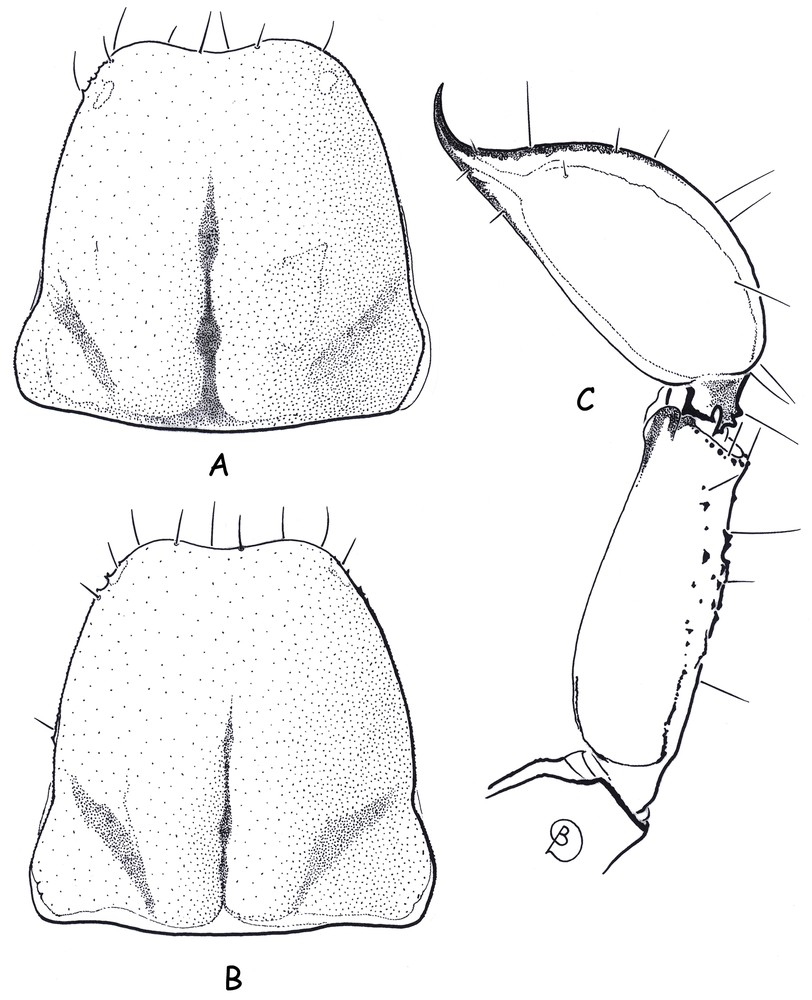
Belisarius xambeui, female holotype (A–C) and female topotype (B). A–B. Carapace, dorsal aspect. C. Metasomal segment V and telson, lateral aspect.
B. ibericus sp. n. (Figs. 4 and 5)

Belisarius ibericus sp. n. Female holotype. A–B. Carapace, dorsal aspect and detail of lateral eye. C. Metasomal segment V and telson, lateral aspect. D. Cutting edge of movable finger with rows of granules. E–H. Chelicera. E. Ventral aspect. F–G. Movable finger in detail, showing the serrula. H. Dorsal aspect.
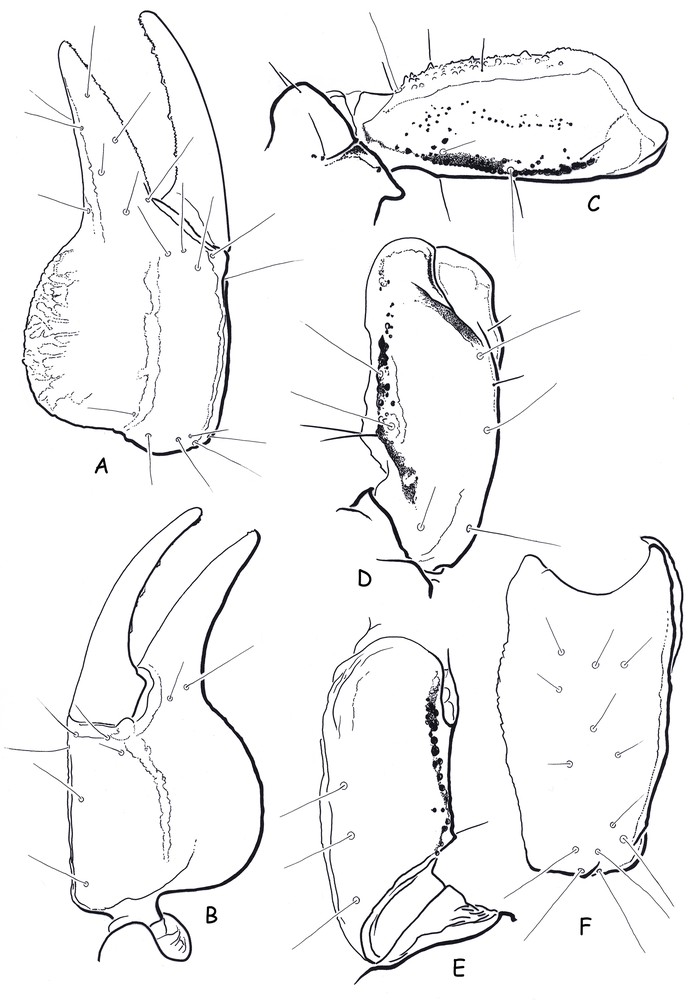
Belisarius ibericus sp. n. Female holotype. Trichobothrial pattern. A–B. Chela, dorso-external and ventro-internal aspects. C. Femur, dorsal aspect. D–F. Patella, dorsal, ventral and external aspects.
Type material: Spain, Province of Malaga, Sierra de las Nieves, south of Ronda, 800–860 m, under rocks near to a cave entry, V/1952 (J. Malhomme). Holotype female deposited in the Muséum national d’histoire naturelle, Paris (RS-7748).
Etymology: The specific name refers to Iberia, the geographical region were the new species was collected.
Diagnosis. Species large in size when compared to the average size observed for adult females of B. xambeui. The female holotype reaches 37.8 mm, while the biggest examined female topotype of B. xambeui reached only 26.7 mm. Coloration reddish–yellow to dark reddish with dark to blackish carinae; only the venter and legs are yellow; pedipalps are reddish with dark to blackish carinae. Body smooth; appendages very weakly granulated or smooth; carapace and tergites with minute punctation. Anterior margin of carapace with a moderately marked concavity (Fig. 4). Median eyes absent; lateral eyes represented by one vestigial lens (Fig. 4). Pectines with 4–4 teeth in the female holotype. Trichobothrial pattern type C orthobothriotaxic. Chela with four ventral trichobothria; patella with three ventral and 13 external trichobothria. The external trichobothrium on the femur is partially displaced over the dorsal surface (Fig. 5).
Relationships. The new species can be distinguished from B. xambeui Simon 1879 (the only other species previously known to this genus) by the following features: (i) a globally bigger size with distinct morphometric values – see measurement after the description –, (ii) anterior margin of carapace with a better marked concavity, (iii) lateral eyes less vestigial than those observed for B. xambeui, with at least one marked lens, (iv) carinae on pedipalps and metasomal segments generally better marked, (v) telson more globular, flattened and with a better marked annular-ring, (vi) trichobothrium e on the femur partially displaced over the dorsal surface.
Description based on holotype.
Coloration. Generally reddish–yellow to dark reddish (the specimen is partially faded in alcohol). Prosoma: carapace reddish. Tergites reddish–yellow, paler than the carapace. Metasomal segments reddish–yellow, with blackish zones over a number of carinae; vesicle yellow with the aculeus dark red. Chelicerae yellow; fingers yellow with reddish teeth. Pedipalps dark reddish to reddish-brown; chela darker than patella and femur; some carinae and granulations on the cutting edge of fingers blackish. Legs yellow. Venter and sternites yellowish; pectines and genital operculum pale yellow.
Morphology. Anterior margin of carapace with a moderate concavity; lustrous and acarinate, with minute punctations; furrows shallow. Median eyes absent; lateral eyes represented by one vestigial lens (Fig. 4). Sternum pentagonal, slightly wider than long. Tergites acarinate, smooth and shiny, with punctations. Pectinal tooth count 4–4, fulcra absent. Sternites smooth and shiny without punctations, VII acarinate; spiracles small in size and oval in shape. Metasomal segments with a lustrous tegument; dorsal carinae weak to moderate on all segments; latero-dorsal carinae strongly marked on segments I to IV; other carinae absent; segment V with spinoid granulations on ventral aspect; vesicle smooth, globular and flattened dorso-ventrally; aculeus short and strongly curved; an annular-ring is well marked (Fig. 4). Pedipalps: femur with dorsal internal, dorsal external and ventral internal carinae moderately to strongly marked; ventral external carina absent; dorsal and ventral faces with minute granulations; internal face with spinoid granules. Patella almost smooth and lustrous; dorsal internal and ventral internal carinae moderate; ventral external carinae weak to vestigial; other carinae absent. Chela lustrous; ventral median and dorsal internal carinae moderate; other carinae weak to vestigial; internal face with a few minute granules, other faces smooth. Dentate margins on movable and fixed fingers with a median denticle row composed of five to six groups of granules (Fig. 4). Chelicerae with the dentition characteristic of the genus Belisarius and of the family Troglotayosicidae [3,8,9]; movable finger with the presence of a serrula [9]; an intense setation is present on the ventral aspect (Fig. 4). Trichobothriotaxy type C; orthobothriotaxic [6]; chela with four ventral trichobothria; patella with three ventral and 13 external trichobothria; trichobothrium e on femur partially displaced over the dorsal surface (Fig. 5). Ventral surface of tarsus in legs III and IV with a few median small spines and 4–5 external and internal setae.
Comparative morphometric values (in mm) of female holotype and a female topotype of B. xambeui and female holotype of B. ibericus sp. n.
Total length (including telson) 26.5/26.6/37.8. Carapace: length 3.6/3.7/5.3; anterior width 2.1/2.2/3.2; posterior width 3.7/3.8/5.5. Mesosoma length 8.6/9.4/13.2. Metasomal segment I: length 1.5/1.3/1.8, width 1.9/1.8/2.8; II: length 1.6/1.5/2.2, width 1.7/1.7/2.3; III: length 1.8/1.7/2.4, width 1.6/1.6/2.2; IV: length 2.2/2.0/2.9, width 1.4/1.4/1.9; V: length 3.2/3.3/4.8, width 1.3/1.3/1.8, depth 1.2/1.2/1.6. Telson length 4.0/3.7/5.2. Vesicle: width 1.8/1.7/2.5, depth 1.5/1.3/2.0. Pedipalp length 12.9/12.7/17.5: femur length 3.1/3.1/4.2, width 1.2/1.2/1.8; patella length 3.2/3.2/4.4, width 1.2/1.3/2.0; chela length 6.6/6.4/8.9, width 2.2/2.1/3.1, depth 3.0/2.9/4.4; movable finger length 3.4/3.3/5.2.
4 Ecology of the Natural Park of Sierra de la Nieves
The Natural Park of Sierra de las Nieves, declared by the UNESCO as a Biosphere's Reserve, is located inside the natural region of the Serrania de Ronda, in the province of Malaga (Fig. 6). This mountain region presents some deep ravines and spectacular gorges. The majority of the region is associated to a limestone formation. The highest altitude achieved is the Torrecila Mountain (1919 m), at the Sierra Blanca of Tolox.
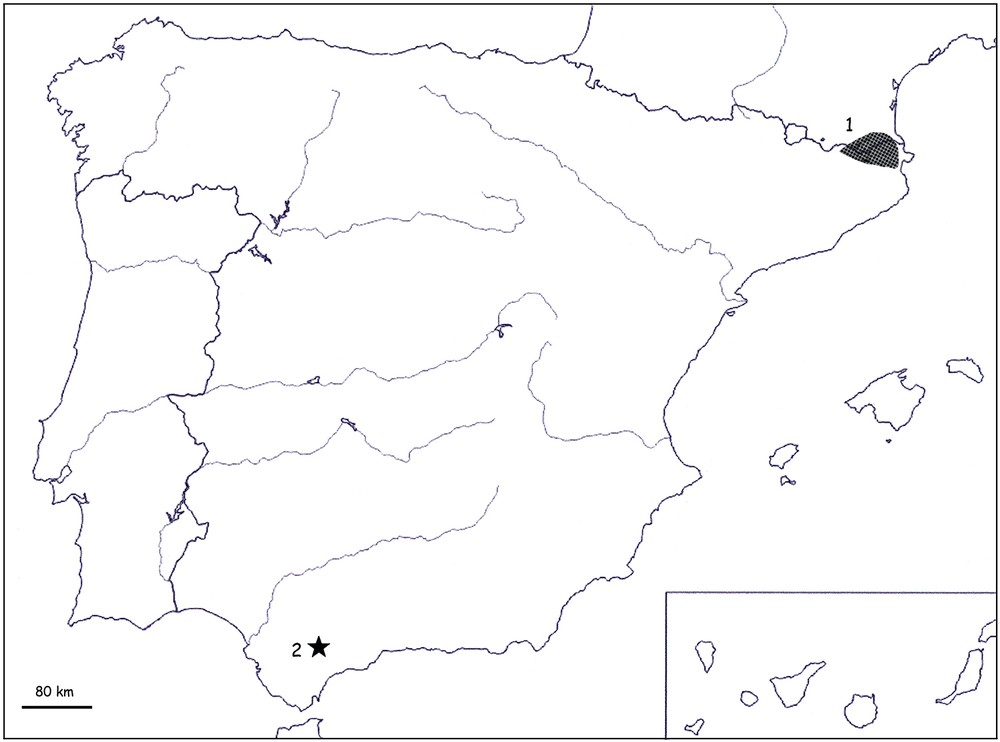
Map of Spain and South of France, showing the known area of distribution of B. xambeui (1) and the type locality of Belisarius ibericus sp. n. (black star, 2).
The Park comprises a total surface of 20,163 ha. Inside the park there are no main roads, only forest ways, which largely helps the preservation of the ecological system. In account to its particular location between the Atlantic and the Mediterranean systems, the park presents an original and quite diverse flora [10]. In the highest zones/altitudes of the mountain system, there is a relatively high level of rainfall, and there is a good coverage of vegetation (trees and bushes), predominately the “Spanish Fir”, a species of tree mainly distributed in the south of the Iberian Peninsula that cover more than 3000 ha in the park. Areas near small brooks and springs are ideal for the ash (or fresno in Spanish, Fraxinus). The chestnut tree or castaño (Castanea sativat) prefers the moist areas and silica soil at altitudes between 400 and 1000 m. In the drier areas, pine trees can be found. Pinus pinaster aiton is the most common, and since it is highly resistant to cold, it can be found even at 1700 m. The P. halepensis prefers the lower slopes and is found up to 1200 m. The third pine found in the park is Pino pinonero. Its presence is entirely due to reforestation. The pine forest constitutes 2000 ha of the park (Fig. 7). The carob tree or algarrobo (Ceratonia siliqua) mixes with the pines, and grows spontaneously all over the park. C. siliqua can withstand high temperatures and is found between 600 and 1000 m. The common Quercus found all over the Mediterranean is equally found in the park, in the form of the gall oak or quejigo (Q. faqinea) and the evergreen oak or encina (Q. ilex). The wild olive tree or acebuche (Olea europea) is one of the species best adapted to the Mediterranean climate. Cork oak is also found in the park, mainly in the west around Mount Alcornoque.

(Colour online.) View of Alpine region in the Natural Park Sierra de las Nieves, with the typical pine vegetation.
Photo courtesy, SINATUR.
In conclusion, in the region defined by the park, environmental conditions are clearly more mesic than those found in the lower nearby regions close to the Mediterranean Sea and the Atlantic Ocean.
5 Biogeographic remarks
The general patterns of distribution observed for a given zoological (or scorpion) group are largely dependent on the historical factors that took place in more or less recent periods of time. The supposed climatic “stability” has been interrupted by periods of climatic change during several dry/wet/dry episodes of the Pleistocene and Holocene epochs.
This historical reduction of alpine vegetation to patch refugia is supported by the existing biogeographical patterns of distribution and differentiation of several taxa, and by palynological and geomorphological evidence from many regions of the world. Several authors also postulate that the reduction of forest to small patches stimulated many changes in plant and animal populations in the refugia. These ranged from extinction to an increased rate of speciation [11].
For the present climate in the Iberian Peninsula, Schütt [12] stated as follows: “Iberian Peninsula can be divided into two large climatic regions: the permanently humid ‘zona verde’ at the northern periphery and dry-summer ‘zona parda’ of central and southern Iberia. According to the degree of dependence on the form and intensity of maritime influence, the dry-summer zone of Iberia is subdivided into the Atlantic-influenced western periphery, the Mediterranean-influenced southern and eastern periphery, and the central regions which are influenced by a continental climate”. As for the palaeoclimatic conditions in the Iberian peninsula, this author also stated the following: “Since the Last Glacial Maximum, the climate of the Iberian Peninsula has changed from a subarctic climate in large parts of central and northern Iberia into the typically Mediterranean climate presently prevailing in the area between the Betic Cordillera in the south and the Pyrenees and Cantabrian Range in the north. The Holocene climate underwent a zonal change in which large-scale topographic conditions were repeatedly modified by local topography. By contrast, the impact of maritime and continental climates was limited to a few locations and single periods. Already since the Atlantic period, a strongly Mediterranean climate has prevailed in large parts of inland and southern Iberian Peninsula, only interrupted by short phases of slightly higher humidity in the Sub-Boreal and during the climatic cooling of the Little Ice Age. Since antiquity at the latest, land degradation and desertification processes have increasingly modified the natural environment”.
The present patterns of distribution observed for the two Belisarius species can be tentatively explained as the result of a past continuous distribution over more mesic formations, which have changed subsequently during recent Quaternary vicissitudes in a vicariant process [13].
Disclosure of interest
The author declares that he has no conflicts of interest concerning this article.
Acknowledgements
I am most grateful to Bernard Duhem and Élise-Anne Leguin, Muséum, Paris for respectively the preparation of the drawings and photographs, and to Michael M. Webber, University of Nevada, Las Vegas for her review of an earlier version of the manuscript.