

1 Introduction
The scorpion fauna of Mexico is among the highest worldwide, especially in the arid environments of the Baja California Peninsula [1], but also in Central Mexico [2,3]. Recently, several new species have been discovered due to the extensive collecting trips in many unexplored areas as part of several scorpion projects in the country (i.e. Revsys: Systematics of the scorpion family Vaejovidae http://www.vaejovidae.com/; Aracnidos y Aracnidismo en México http://www.aracnidos.unam.mx). The scorpion genus Diplocentrus Peters, 1861 presently comprises 61 species distributed from the southwestern USA to northern Honduras [4], but its highest diversity and endemism are found in southern Mexico [3,5]. Despite recent efforts to summarize its diversity in Guatemala [6], Mexico [3], and Honduras [7], and a preliminary phylogenetic analysis [4], the systematics of the genus remains incomplete.
The present contribution describes a new species of Diplocentrus, endemic to the Valley of Zapotitlán de Salinas, in the biogeographic province of the Valley of Tehuacán–Cuicatlán ([8]; Figs. 1 and 2 in this article), an area shown to be highly diverse [3]. Diplocentrus duende n. sp. is the fifth species to be described possessing punctate pedipalp surfaces, besides Diplocentrus bereai Armas & Martín-Frías, 2004, Diplocentrus gertschi Sissom & Walker, 1992, Diplocentrus melici Armas, Martín-Frías & Berea, 2004 and Diplocentrus motagua Armas & Trujillo, 2009. We hypothesized that D. duende n. sp. belongs to the mexicanus group defined by Santibañez-López et al. [4] by the presence of that character. Although punctate pedipalp surfaces are a homoplastic character at the generic level [9], as it has evolved independently several times within Diplocentridae (e.g., in Bioculus Stahnke, 1968; Didymocentrus Kraepelin, 1905; Kolotl Santibáñez-López et al., 2014), it appears to be a suitable character to group some species within the mexicanus group of Diplocentrus.
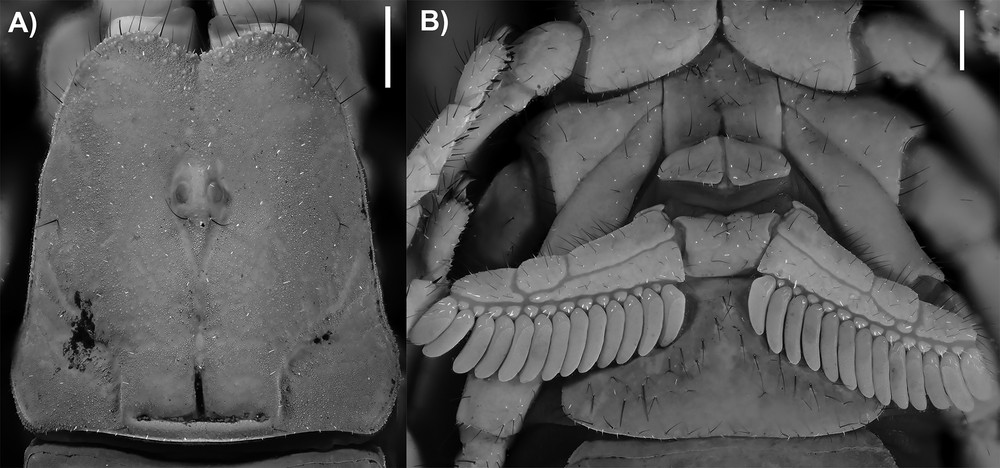
Diplocentrus duende n. sp., paratype ♂. A. Carapace, dorsal aspect. B. Sternum, genital operculum and pectines, ventral aspect. Scale bars = 1 mm.

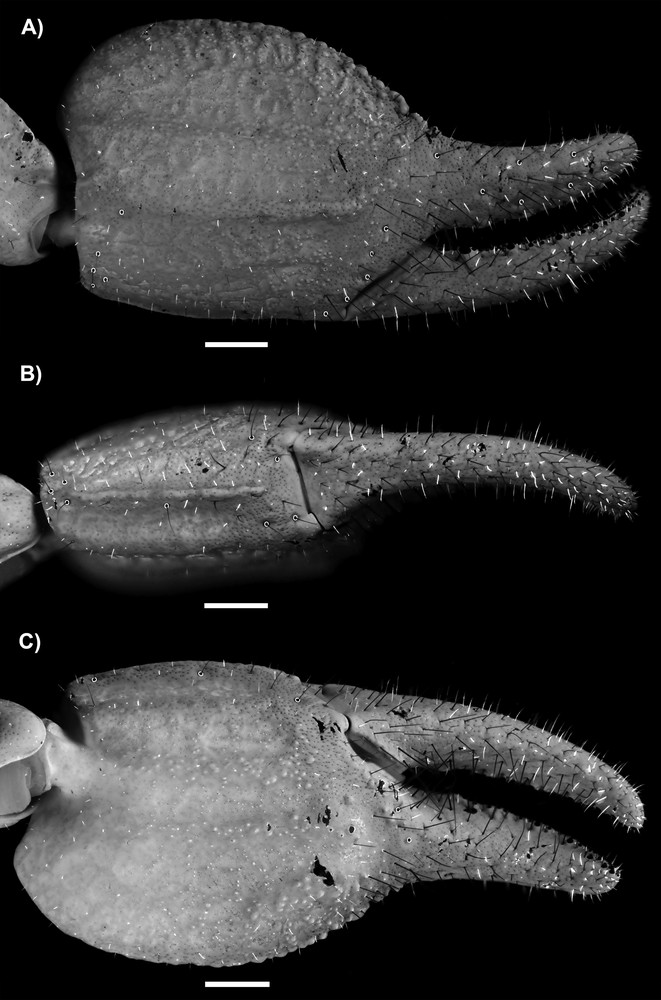
Diplocentrus duende n. sp., paratype ♂. A. Dextral pedipalp femur, dorsal aspect. B. Dextral pedipalp patella, dorsal aspect. C. Dextral pedipalp patella, retrolateral aspect. D. Dextral pedipalp patella, ventral aspect. Scale bars = 1 mm.
2 Material and methods
Scorpions were collected at night with ultraviolet (UV) light detection [10]. Geographical coordinates of collection localities were recorded in the field with a GARMIN eTREX H GPS device.
The material has been deposited in the following collections: American Museum of Natural History, New York (AMNH), with tissue samples stored in the Ambrose Monell Cryocollection (AMCC); Colección Nacional de Arácnidos, Instituto de Biología, Universidad Nacional Autónoma de México, Mexico City (CNAN).
Observations were made using Nikon SMZ-800 and SMZ-1500 stereomicroscopes. Measurements, given in millimeters, were obtained with an ocular micrometer calibrated at 10× magnification. Hemispermatophores were dissected following Vachon [11], and cleared with pancreatin. Digital images were taken under visible and UV light with a Microptics ML-1000 digital imaging system, equipped with a Nikon DS80 camera, or a Nikon SMZ-800 with Nikon Coolpix S10 VR camera attachment. The focal planes of image stacks were fused with CombineZM [12] and composite images edited with Adobe Photoshop CSS.
The scorpion higher classification follows Sharma et al. [13]. Nomenclature and mensuration follows Stahnke [14], except for hemispermatophore San Martín [15], trichobothrial pattern Vachon [16], Francke [17] for metasomal carination, Prendini [18] for pedipalpal carination, carapace surfaces Prendini et al., [19], and for tarsal setation counts Santibáñez-López, [20] and Santibáñez-López et al., [4,5,21].
Systematics
Family Diplocentridae Karsch, 1880
Genus Diplocentrus Peters, 1861
“mexicanus” species group
Diplocentrus duende n. sp.
Diplocentrus duende n. sp., paratype ♂, dextral pedipalp chela. A. Dorsal, retrolateral aspect. B. Ventral aspect. C. Prolateral aspect. Scale bars = 1 mm.

Diplocentrus duende n. sp., paratype ♂, habitus. A. Dorsal. B. Ventral.

Live habitus of Diplocentrus duende n. sp., paratype ♂.
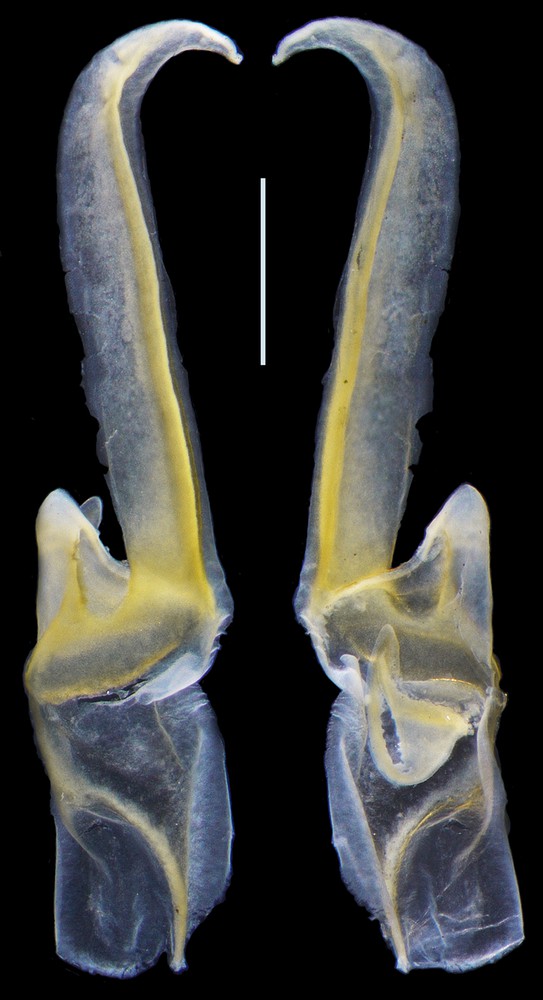
Diplocentrus duende n. sp., holotype ♂ (CNAN) sinistral hemispermatophore. Scale bar = 1 mm.
Measurements (mm) of type specimens of Diplocentrus duende n. sp. deposited in the following collections: American Museum of Natural History, New York (AMNH), and Colección Nacional de Arácnidos, Instituto de Biología, Universidad Nacional Autónoma de México, Mexico City (CNAN).
| CNAN | AMNH | AMNH | CNAN | CNAN | CNAN | CNAN | |
| ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | |
| Holotype | Paratype | Paratype | Paratype | Paratype | Paratype | Paratype | |
| Chelicera length | 2.8 | 2.5 | 2.5 | 2.7 | 2.4 | 2.8 | 2.5 |
| Width | 1.1 | 1.1 | 1.3 | 1.4 | 1.2 | 1.3 | 1.2 |
| Movable finger length | 1.6 | 1.5 | 1.7 | 1.6 | 1.6 | 1.6 | 1.4 |
| Fixed finger length | 1.0 | 1.0 | 0.9 | 1.0 | 1.0 | 1.0 | 1.0 |
| Carapace length | 5.4 | 5.0 | 5.4 | 5.5 | 5.2 | 5.3 | 5.0 |
| Anterior width | 3.3 | 3.1 | 3.3 | 3.5 | 3.3 | 3.4 | 3.0 |
| Posterior width | 5.0 | 4.9 | 5.4 | 5.6 | 4.9 | 4.9 | 4.9 |
| Pedipalp: femur length | 4.4 | 4.2 | 4.9 | 4.8 | 4.5 | 4.9 | 4.1 |
| Femur width | 2.0 | 1.7 | 1.8 | 1.8 | 1.6 | 1.8 | 1.5 |
| Femur height | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 0.9 |
| Patella length | 4.6 | 4.1 | 5.0 | 4.9 | 4.4 | 5.0 | 4.4 |
| Patella width | 1.9 | 1.6 | 1.8 | 1.9 | 1.7 | 1.8 | 1.7 |
| Patella height | 2.0 | 2.0 | 2.3 | 2.1 | 2.0 | 2.4 | 2.1 |
| Chela length | 8.8 | 8.1 | 9.5 | 9.4 | 8.6 | 9.7 | 8.2 |
| Chela width | 3.1 | 2.8 | 3.2 | 3.1 | 3.1 | 3.3 | 3.0 |
| Chela height | 2.7 | 2.4 | 2.7 | 2.6 | 2.5 | 2.6 | 2.5 |
| Manus length | 5.3 | 4.9 | 5.7 | 5.6 | 5.2 | 5.8 | 5.0 |
| Fixed finger length | 3.5 | 3.2 | 3.8 | 3.8 | 3.4 | 3.9 | 3.2 |
| Movable finger length | 5.3 | 4.6 | 5.5 | 5.5 | 5.1 | 5.4 | 4.7 |
| Mesosoma length | 11.4 | 10.1 | 10.4 | 11 | 10 | 10.3 | 9.3 |
| Coxa II length | 2.0 | 2.0 | 2.2 | 2.4 | 2.2 | 2.3 | 2.0 |
| Coxa IV length | 3.3 | 3.2 | 3.7 | 3.7 | 3.5 | 3.7 | 3.3 |
| Sternum length | 1.9 | 1.8 | 2.0 | 2.1 | 2.1 | 2.0 | 1.8 |
| Sternum ant. Width | 1.7 | 1.6 | 1.8 | 1.8 | 1.8 | 1.8 | 1.7 |
| Metasoma length | 17 | 15.7 | 18.8 | 17.4 | 16.8 | 19.1 | 17.2 |
| Segment I length | 2.5 | 2.1 | 2.9 | 2.5 | 2.4 | 2.8 | 2.5 |
| Segment I width | 3.2 | 3.0 | 3.2 | 3.2 | 3.1 | 3.2 | 3.1 |
| Segment I height | 2.3 | 2.0 | 2.3 | 2.3 | 2.2 | 2.3 | 2.0 |
| Segment II length | 2.8 | 2.8 | 3.1 | 2.8 | 2.8 | 3.2 | 2.9 |
| Segment II width | 2.9 | 2.7 | 3 | 3 | 2.8 | 3 | 2.8 |
| Segment II height | 2.3 | 2.0 | 2.3 | 2.3 | 2.1 | 2.3 | 2.0 |
| Segment III length | 3.0 | 2.8 | 3.4 | 3.1 | 3.0 | 3.5 | 3.0 |
| Segment III width | 2.8 | 2.6 | 2.8 | 2.9 | 2.7 | 2.9 | 2.6 |
| Segment III height | 2.2 | 2.0 | 2.2 | 2.2 | 2.1 | 2.2 | 2 |
| Segment IV length | 3.9 | 3.7 | 4.3 | 4 | 3.8 | 4.4 | 3.9 |
| Segment IV width | 2.5 | 2.4 | 2.5 | 2.6 | 2.5 | 2.6 | 2.5 |
| Segment IV height | 2.0 | 1.9 | 2.0 | 2.1 | 2.0 | 2.0 | 2.0 |
| Segment V length | 4.8 | 4.3 | 5.1 | 5 | 4.8 | 5.2 | 4.9 |
| Segment V width | 2.1 | 1.9 | 2.1 | 2.1 | 2 | 2.1 | 1.9 |
| Segment V height | 1.7 | 1.6 | 1.7 | 1.8 | 1.7 | 1.7 | 1.7 |
| Telson length | 4.2 | 4.0 | 4.6 | 4.6 | 4.1 | 4.5 | 4.0 |
| Vesicle length | 3.5 | 3.4 | 3.9 | 3.9 | 3.5 | 3.9 | 3.4 |
| Vesicle width | 2.1 | 1.9 | 2.1 | 2.3 | 2.1 | 2.1 | 2 |
| Vesicle height | 1.6 | 1.3 | 1.6 | 1.5 | 1.6 | 1.5 | 1.3 |
| Aculeus length | 0.7 | 0.6 | 0.7 | 0.7 | 0.6 | 0.6 | 0.6 |
| Total l | 38.0 | 34.8 | 39.2 | 38.5 | 36.1 | 39.2 | 35.5 |
Telotarsal spiniform macrosetal counts (number of macrosetae in pro- and retroventral rows of telotarsi on legs I–IV) in Diplocentrus duende n. sp., given as number of legs observed with corresponding proventral (p) and retroventral (r) setal counts.
| Leg I | Leg II | Leg III | Leg IV | |||||||||||||
| p | r | p | r | p | r | p | r | |||||||||
| 4 | 5 | 4 | 5 | 4 | 5 | 5 | 6 | 5 | 6 | 5 | 6 | 5 | 6 | 5 | 6 | 7 |
| 14 | 2 | 1 | 15 | 1 | 15 | 14 | 2 | 3 | 13 | 1 | 15 | 1 | 13 | 1 | 11 | 3 |
Type material.MEXICO: Puebla: Municipio Zapotitlán de Salinas: Holotype ♂ (CNAN-T0907), Botanical Garden Helia Bravo Holis, on trails and wild land owned by the Botanical Garden, 18.32798°N 97.45437°W, 1440 m. 22.v.2014, C. Santibáñez, U. Hernández, R. Monjaraz and A. Guzmán. Paratypes: 5 ♂ (CNAN-T0908), 1 ♂ and 1 subad. ♂ paratypes (AMNH) same data as holotype.
Etymology. “Duende” is the Spanish word for Goblin, nocturnal creatures of the common folklore conceived as small and mischievous. It is used as a noun in apposition and it is considered masculine in gender.
Diagnosis. The following character combination is diagnostic for D. duende, n. sp. Total length (♂ adults only), 35–40 mm. Base coloration (adult), yellowish brown to light reddish. Carapace anteromedian notch weakly to moderately deep, V-shaped (Fig. 1A). Pedipalp femur width greater than height, dorsal intercarinal surface slightly convex (Fig. 2A). Pedipalp patella, dorsal median carina weakly developed to obsolete, smooth; dorsal retrolateral carina weakly developed to obsolete, smooth; retrolateral median carina weakly developed to obsolete, smooth; ventral median carina obsolete. Pedipalp chela manus, dorsal intercarinal surface weakly reticulated, punctate (♂; Fig. 3A); digital carina moderately developed, smooth (♂); dorsal secondary carina weakly developed to obsolete, smooth. Legs I–IV telotarsi, counts of spiniform macrosetae in pro- and retroventral rows, 4/5:5/5:6/6:6/6; basitarsi, spiniform macrosetae pattern, leg I pst, rst; leg II pt, rt, pst, rst, Rm; legs III–IV pt, rt, vt, vst. Pectinal tooth count, 11–14 (♂).
Diplocentrus duende n. sp. can be assigned to the mexicanus group [4], by sharing with D. bereai, D. gertschi, D. melici and D. motagua the presence of punctate pedipalp chela surface, but can be readily differentiated from these species as follows. The body coloration is medium brown in D. duende n. sp. and in D. bereai, it is dark orange brown in D. gertschi, but yellowish brown in D. motagua and dark brown in D. melici. The counts of spiniform macrosetae of legs III and IV are higher in D. duende n. sp., D. gertschi and D. melici (6/6), than in D. bereai and D. motagua (5/5). The pm and rm spiniform macrosetae in basitarsi of leg II are present in D. bereai, D. gertschi, D. melici and D. motagua (except rm), but they are absent in D. duende n. sp.; the vm spiniform macroseta in basitarsi of legs III and IV is present in D. bereai and D. melici, but absent in D. duende n. sp., in D. gertschi and in D. motagua.
Description. Based on holotype ♂ and paratype ♂ (Figs. 4 and 5). Measurements are given in Table 1.
Coloration. Carapace medium to light brown, with moderate fuscosity throughout, uniform around median eyes and variegated elsewhere; venter cream to pale orange. Pedipalps brown to brown orange, carinae darker. Mesosoma medium brown, venter pale brown, tergites light to medium brown, with moderately dense variegated fusco-piceous pattern; sternites medium brown to pale brown. Metasoma medium brown to light brown. Telson orange brownish, uniformly fuscous. Legs light brown, uniformly infuscated.
Carapace. Anterior margin weakly setose, anteromedian notch moderately deep, V-shaped (Fig. 1A). Frontal lobes and interocular surface weakly granular; other surfaces shagreened.
Pedipalps. Orthobothriotaxic, Type C. Femur width greater than height; dorsal intercarinal surface flat, sparsely granular medially, punctated (Fig. 2A); retrolateral intercarinal surface punctated; ventral intercarinal surface flat, punctated; prolateral intercarinal surface coarsely and slightly granular, with medium-size granules; dorsal prolateral carina strongly developed, granular; dorsal retrolateral carina moderately developed, densely granular proximally and smooth distally; ventral retrolateral carina obsolete; ventral prolateral carina strongly developed, granular proximally, becoming obsolete distally. Patella, dorsal, retrolateral and ventral intercarinal surfaces slightly punctated; prolateral intercarinal surface moderately granular distally; proximal tubercle moderately developed; dorsal prolateral carina weakly developed to obsolete, slightly granular; dorsal median carina weakly developed to obsolete, smooth; dorsal retrolateral carina weakly developed to obsolete, smooth (Fig. 2B); retrolateral median carina weakly developed to obsolete, smooth (Fig. 2C); ventral retrolateral carina moderately developed, crenulate to smooth; ventral median carina obsolete; ventral prolateral carina strongly developed, granular, comprising large granules (Fig. 2D). Chela manus, slender, width greater than height (♂), moderately setose; dorsal intercarinal surface weakly reticulated, punctate (♂; Fig. 3); retrolateral intercarinal surface weakly reticulated, punctate; dorsal marginal carina moderately developed, granular; digital carina moderately developed, smooth (♂); dorsal secondary and retrolateral secondary carinae weakly developed to obsolete, smooth (Fig. 3A); ventral retrolateral carina weak to obsolete, smooth distally, becoming obsolete proximally; ventral median carina strongly developed, crenulated proximally, becoming obsolete distally, directed towards midpoint of movable finger articulation; ventral prolateral carina weakly to moderately developed, crenulated (Fig. 3B); prolateral dorsal carina weakly developed, smooth to slightly granular; prolateral median and prolateral ventral weakly developed, smooth; prolateral surface with shallow longitudinal depression where chela rests against patella (Fig. 3C). Chela fixed finger curved; length shorter (♂) than femur length and patella length; dorsal surface smooth and densely setose proximally, retrolateral surface flat; prolateral surface shallowly concave.
Legs. Legs I–IV femora and tibiae, prolateral surfaces shagreened; telotarsi, counts of spiniform macrosetae in pro- and retroventral rows (dextral/sinistral), 4/5 4/5: 5/5 5/6: 6/6 6/6: 6/6 6/6 (holotype; variation in Table 2). Basitarsi, spiniform macrosetae pattern, leg I pst, rst; leg II pt, rt, pst, rst, Rm; legs III–IV pt, rt, vt, vst, (holotype); these setae were constant in all legs, except for two legs I, which presented setae pt and rt.
Pectines: Tooth count: 13-13 (♂; Fig. 1B). Variation: one comb with 11 pectines, four combs with 12 pectines, and 10 combs with 13 pectines.
Mesosoma. Tergites I–VI with pre-tergites smooth, post-tergites sparsely granular, VII granular. Sternites smooth; VII, submedian carinae weakly developed, slightly granular to crenulated proximally, fading distally; lateral carinae weakly to moderately developed, smooth to slightly granular proximally, moderately developed, crenulated distally.
Metasoma. Metasomal segments I–V, dorsal and lateral intercarinal surfaces punctated; ventral intercarinal surfaces smooth on segments I–II, punctated on IV–V. Segments I–IV, dorsal lateral carinae weakly to obsolete, granular, (♂); lateral supramedian carinae moderately developed, granular to slightly crenulated on I, crenulated to smooth on II–IV; lateral inframedian carinae weakly developed, granular to slightly crenulated on I, weak to obsolete, crenulated to smooth on II–IV; ventral lateral carinae moderately developed, crenulated to smooth on I–III, weakly developed, crenulated on IV; ventral submedian carinae moderately developed, crenulated to smooth on I–III, weak to obsolete, smooth on IV. Segment V length:pedipalp femur length ratio, 1.12 (♂); dorsal lateral carinae moderately developed, crenulated; lateral inframedian carinae obsolete; ventral lateral carina moderately developed, crenulated to slightly serrated; ventral median carina moderately developed, granular, with subspiniform granules; ventral transverse carina moderately developed, comprising three subspiniform granules; anal arch semicircular; anal subterminal carina moderately to weakly developed, comprising ten subspiniform granules; anal terminal carina vestigial, weakly granular.
Telson. Telson width:length ratio, 0.5 (♂). Vesicle, lateral surfaces punctated; ventral surface granular anteriorly, punctated posteriorly. Subaculear tubercle stout, subconical. Aculeus length 1.
Hemispermatophore. Lamelliform, weakly sclerotized (Fig. 6); total length 6.5; distal lamella length 3.7 mm; capsular region, width, 1.3; median lobe narrow, margin entire.
Distribution. Diplocentrus duende n. sp. is recorded only from the type locality (Fig. 7).
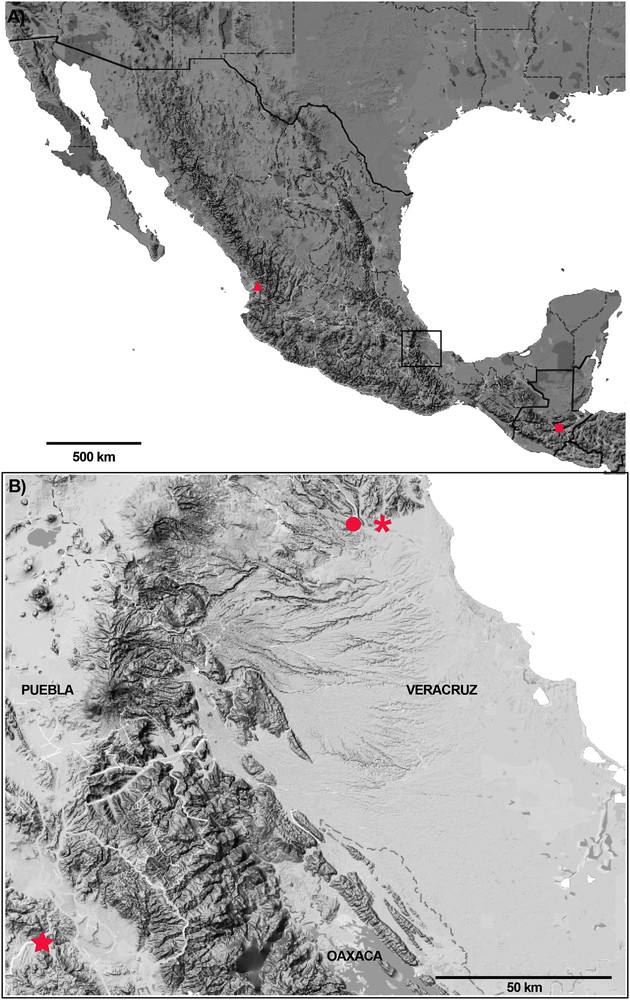
Map plotting known type localities or known records of the species with pedipalp punctated in genus Diplocentrus. A. Diplocentrus gertschi, in Mexico (in triangle); Diplocentrus motagua, in Guatemala (in square). B. Diplocentrus bereai, in Veracruz, Mexico (in circle); Diplocentrus melici, in Veracruz, Mexico (in asterisk); Diplocentrus duende n. sp., in Puebla, Mexico (in star).
Ecology. This species was collected with the aid of UV light walking during night time. The dominant vegetation was thorny deciduous forest and columnar cacti. Centruroides baergi Hoffmann, 1932, Mesomexovis sp., Mesomexovis subcristatus Hoffmannihadrurus aztecus (Pocock, 1902) and Vaejovis solegladi Sissom, 1991 were collected in sympatry. D. duende n. sp., as other diplocentrids, exhibits a fossorial biology [4]. Two species were dug out, the gallery was spiral in shape and had ca. 100 mm in depth.
3 Species of Diplocentrus with punctate pedipalp surfaces
Punctate pedipalp surfaces in diplocentrid scorpions are not rare. This condition was previously used to diagnose species of Didymocentrus Kraepelin, 1905, until genus Bioculus Stahnke, 1968 was described. Sissom & Walker [22] described Diplocentrus gertschi, the first species with punctate pedipalp chela, considering as a “link” to Bioculus due to this condition and similar development of chela carination [4]. Armas & Martín-Frías [23] and Armas et al. [24] described two more species with punctate pedipalp surfaces: D. bereai and D. melici respectively. In both cases, these species were compared to each other, and no arguments on their relationship were made; however, Armas et al. [24] highlighted that, among the main differences between D. melici, D. gertschi and D. bereai, was that the former one had punctate surfaces in all the body, whereas the other two species did not.
Later, Armas & Trujillo [25] described the fourth species: D. motagua, from Guatemala. This species also had punctate surfaces in the entire body (like in D. melici); and the authors commented the differences between these species and the species of Didymocentrus. Recently, in the first phylogenetic analysis of Diplocentrus [4], these species did not form a monophyletic clade (except for D. bereai and D. melici, which were recovered as sister species), suggesting that this condition has evolved independently at least three times in the “mexicanus” group. More recently, the convergence of the punctate pedipalp femur and chela surfaces in Diplocentridae was measured [9]. In this analysis, both characters were informative and evolved not randomly within the family. Furthermore, these characters were strongly correlated to each other, to the development of the digital carina of the female's pedipalp chela, and to the presence/absence of a prolateral concavity on the male's pedipalp chela [9].
In this contribution, a fifth species with punctate pedipalp surfaces was described. According to the previous analyses [4,9], the current explored morphological characters cannot diagnose the three species groups within Diplocentrus. For example, all characters used to separate species of the “keyserlingii” group have appeared at least two times in the “mexicanus” group (D. coylei and D. formosus; see [4]). However, punctate pedipalp surfaces have appeared only in the “mexicanus” group. Therefore, this condition appears to be unique to the “mexicanus” group of Diplocentrus. It is true that a morphological differentiation between the “mexicanus” and “zacatecanus” groups is still missing; however, the four recognized members of the “zacatecanus” group have the following legs III–IV basitarsi spiniform macrosetae formula: pt, rt, vt, rst, vst and vm; and none of the species with punctate pedipalp surfaces have this configuration. Until a more complete morphological characterization of the “zacatecanus” group that differentiates it from the “mexicanus” group is available, all species with punctate pedipalp surfaces should be included in the “mexicanus” group; suggesting punctate surfaces have evolved independently in this group from the rest of the diplocentrids. Therefore, this new species is included in the “mexicanus” group.
Acknowledgements
This research was funded by a postdoctoral grant from CONACyT to EGS and to CESL. Special thanks to Lorenzo Prendini (AMNH) and Fernando Alvarez-Padilla (Facultad de Ciencias, UNAM) for access to microphotography facilities in their respective labs. E. Balcazar, A. Guzmán, U. Hernández, R. Monjaraz and I. Reyes are greatly thanked for their help collecting the type specimens. Fieldwork was conducted under permits issued by the Dirección General de Vida Silvestre (SEMARNAT) to Edmundo González-Santillán (FAUT-0305).