

1 Introduction
The cutthroat eel family Synaphobranchidae is a group of anguilliform fishes found worldwide in tropical and temperate oceans [1]. Adults are usually benthic, found in deep water or deep-sea habitats, while larvae are pelagic, mainly at depths of 0–2.820 m, but most frequently at depths of 450–600 m [1,2]. Synaphobranchids are arranged in the three subfamilies Simenchelyinae, Ilyophinae, and Synaphobranchinae; with 27 valid ilyophine species (compared to 12 synaphobranchine species and one simenchelyine; [3]), the Ilyophinae is the most speciose subfamily, judging from the great variety of leptocephali that have been collected; most of these larvae cannot be referred to a known adult form. They are of little or no importance to fisheries, although they are sometimes taken in deep trawls [4,5].
The genus Dysomma was originally described by Alcock [6], based on Dysomma bucephalus Alcock 1889 as the type species. The genus is mainly characterised by an origin of the dorsal fin well in advance of the adpressed pectoral fin, the presence of enlarged, compound teeth on the dentary and the vomer, a mouth gap extending to below the rear edge of the eye or beyond, a very short trunk, and an entirely naked body [4]. The genus was revised by Robins & Robins [7] who recognized six species. Subsequently, the following species were described: D. fuscoventralis Karrer & Klausewitz [8], D. dolichosomatum and D. polycatodon Karrer [9], D. tridens Robins, Böhlke & Robins in Robins & Robins [10], D. opisthoproctus Chen & Mok [11], D. longirostrum Chen & Mok [12], and D. taiwanensis Ho et al. [13].
During fieldwork aiming at the collection of deep-water fishes of the Gulf of Aqaba, Red Sea, a specimen of the genus Dysomma was collected with a trammel net from a depth of 350 m off Eilat. The specimen is described in the present paper, bringing the total number of species of Dysomma to 14, and the number of congeners occurring in the Red Sea to two.
2 Methods and materials
Descriptive methods follow Ho et al. [13]. The classification follows Eschmeyer et al. [14], unless otherwise noted; reference citations follow Fricke [15]. The key is based on Robins & Robins [10] and Ho et al. [13], but modified according to subsequent findings and expanded to a worldwide scope. The museum abbreviations follow Fricke & Eschmeyer [16].
2.1 Comparative material
Dysomma anguillare: CAS 235128 (1, 557 mm TL), Philippines, Luzon, 146–203 m in depth; CAS 235465 (5, 525–540 mm TL), Philippines, Luzon, 146–203 m in depth; CAS 235129 (2, 517–575 mm TL), Philippines, Luzon, 240–262 m in depth; CAS 235465 (2, 525–540 mm TL), Philippines, Luzon, 355–368 m in depth.
Dysomma brevirostre: BPBM 21068 (1, 123 mm TL), central Pacific Ocean, Hawaiian Islands, north of Maui; HUJ 15679 (1, 145 mm SL), Mediterranean Sea, Cyprus; HUJ 15683 (1, 113 mm SL), Mediterranean Sea, Israel, Rubin-Tel Aviv; HUJ 15722 (1, ca. 100 mm SL), Mediterranean Sea, Israel; HUJ 17694 (1, 164 mm SL), Mediterranean Sea, Israel, Hadera-Atlit, 320 m in depth.
Dysomma dolichosomatum: MNHN 1979–0004 (holotype, 325 mm SL), southwestern Indian Ocean, off southwestern Madagascar; MNHN 1978–0770 (1 paratype, 268 mm SL), western Pacific Ocean, Philippines, southwest of Luzon.
Dysomma fuscoventralis: HUJ 17142 (1, 231 mm TL), Red Sea, Egypt, Strait of Tiran, 1450 m in depth; HUJ 17541 (1, 264 mm TL), Red Sea, Israel, off Eilat, 172 m in depth; SMF 15660 (holotype, 174 mm TL), central Red Sea.
Dysomma goslinei: ANSP 133805 (holotype, 119 mm TL), northeastern Indian Ocean, Myanmar, off the Gulf of Martaban.
Dysomma longirostrum: MNHN 2003–1515 (1, 285 mm SL), southwestern Pacific, Norfolk Ridge.
Dysomma melanurum: USNM 398473 (1, 165 mm SL), western Pacific, Taiwan; USNM 398474 (1, 178 mm SL), western Pacific, Taiwan.
Dysomma muciparus: BPBM 22879 (2, 164–16 mm TL), central Pacific, northeast of Hawaiian Islands.
Dysomma polycatodon: MNHN 1979–0003 (holotype, 523 mm SL), southwestern Indian Ocean, off northwestern Madagascar.
Dysomma taiwanensis: USNM 427172 (2 paratypes, 192–248 mm TL), southwestern Taiwan.
3 Results
3.1 Dysomma alticorpus new species (Figs. 1–2)
Pale cutthroat eel

Dysomma alticorpus new species, HUJ 17054 (holotype, 323.4 mm TL), Red Sea, Gulf of Aqaba, Israel, off Eilat; A lateral view, B lateral view, detail of head, C trunk, ventral view; colouration in preservative.

Dysomma alticorpus new species, HUJ 17054 (holotype, 323.4 mm TL), Red Sea, Gulf of Aqaba, Israel, off Eilat; A distribution of head pores, B dentition, upper jaw, C dentition, lower jaw. The scale equals 5 mm.
3.1.1 Type material
Holotype: HUJ 17054 (323.4 mm TL), Red Sea, Gulf of Aqaba, Israel, off Eilat, ca. 29°30’45”N 34°56’30”E, 350 m in depth, trammel net, A. Baranes, 1 April 1993.
3.1.2 Diagnosis
Pectoral fin present; origin of the dorsal fin well anterior to the base of the pectoral fin, predorsal length 13.8% TL; anus anterior, slightly behind the level of the tip of the pectoral fin, preanal length 22.8% TL; trunk very short, 4.1% TL; body relatively high, depth at the gill opening 7.6% TL, depth at the anus 7.8% TL; two intermaxillary teeth; three compound teeth on the vomer; four rows of teeth on the upper jaw; single row of eight large compound teeth on the lower jaw; head pores: IO 4, SO 3; M 6; POP 0; AD 1, F 0, ST 0; lateral-line pores: predorsal 4, prepectoral 8, preanal 14, total 57–58, the last at the posterior two-thirds of the total length; MVF 7–16–115, total vertebrae 115; peritoneum whitish; dorsal and anal fin pale.
3.1.3 Description
Measurements, proportions and counts of the holotype are provided in Table 1.
Measurements, proportions and counts of the holotype of the new species Dysomma alticorpus; values are given in mm, and as percentage of the total length (TL) or head length (HL), respectively.
| Holotype | ||
| Catalogue no. | HUJ 17054 | |
| Measurements | mm | % of TL |
| Total length | 323.4 | – |
| Caudal-fin length | 7.7 | 2.4 |
| Head length | 51.0 | 15.7 |
| Predorsal length | 48.1 | 13.8 |
| Preanal length | 73.6 | 22.8 |
| Preanus length | 66.2 | 23.6 |
| Trunk length | 13.1 | 4.1 |
| Tail length | 254.0 | 78.5 |
| Depth at gill opening | 24.6 | 7.6 |
| Depth at anus | 25.3 | 7.8 |
| Width at anus | 15.1 | 4.7 |
| % of HL | ||
| Snout length | 9.0 | 17.6 |
| Eye diameter | 3.0 | 5.9 |
| Interorbital width | 7.6 | 14.9 |
| Upper-jaw length | 21.4 | 42.0 |
| Gill-opening diameter | 4.1 | 8.0 |
| Predorsal vertebrae | 7 | |
| Preanal vertebrae | 16 | |
| Total vertebrae | 115 | |
| Preanal lateral-line pores | 9 | |
| Total lateral-line pores | 57–58 | |
| Supraorbital pores | 3 | |
| Infraorbital pores | 5 | |
| Adnasal pores | 1 | |
| Mandibular pores | 6 | |
| Preopercular pores | 0 | |
| Supratemporal pores | 0 | |
| Frontal pores | 0 | |
| Vomerine teeth | 3 | |
| Intermaxillary teeth | 2 |
Body slender but relatively high, head and trunk profile oval, becoming gradually compressed to the rear end; trunk very short, 4.1% TL (Fig. 1C); body width at the anus 4.7% TL; body depth greatest near the anus, depth at the anus 7.8% TL, narrowing gradually to the caudal fin; diameter of the gill opening 1.3% TL. Dorsal and anal fins low and fleshy, continuous with a small caudal fin. Pectoral fin well developed, situated near the upper corner of the gill opening.
Head slender but relatively high in profile; snout pointed anteriorly and rounded dorsally, covered by a few short papillae, snout length 17.6% HL; tip of the snout projecting considerably beyond lower jaw; eye small, covered by a thick and semitransparent membrane; eye diameter 5.9% HL; interorbital space broad, elevated, its width 14.9% HL; postorbital space a little wider than the interorbital one. Anterior nostrils tubular, directed ventrally. Posterior nostril rounded, below the anterior margin of the eye, opening directed posteroventrally. Lower jaw shorter than the upper, its tip below the anterior nostril. Mouth gape far behind eye, upper jaw length 42.0% HL. Tongue well attached to the mouth floor. Gill opening an oval slit.
Head and lateral-line pores large (Fig. 2A); supraorbital pores 3, all restricted to the anterior portion of the snout; infraorbital pores 5, 2 pores between the nostrils and 3 below the eye; mandibular pores 6, all under mouth gape; preopercular pores 0; adnasal 1; supratemporal commissure 0; frontal 0. Lateral line incomplete, extending along the anterior two-thirds of body, predorsal 4, prepectoral 8, preanal 14 and total 58 (right)/57 (left).
Teeth (Fig. 2B, C) small and pointed. Intermaxillary teeth 2, side-by-side, followed by a row of 22 small teeth, plus a second irregular outer row of five teeth besides the posterior section of the inner row. Lower jaw with single row of eight large compound teeth.
Mean vertebral formula 7–16–115; predorsal vertebrae 7, preanal vertebrae 16 and total vertebrae 115.
Colouration. Live colouration unknown. When preserved (Figs. 1, 2), uniformly pale whitish; fins pale whitish; caudal fin yellowish. Peritoneum pale. Mouth cavity white.
3.1.4 Distribution
This species is only known from the holotype, which was collected at a depth of 350 m off Eilat, Israel, Gulf of Aqaba, Red Sea (Fig. 3).
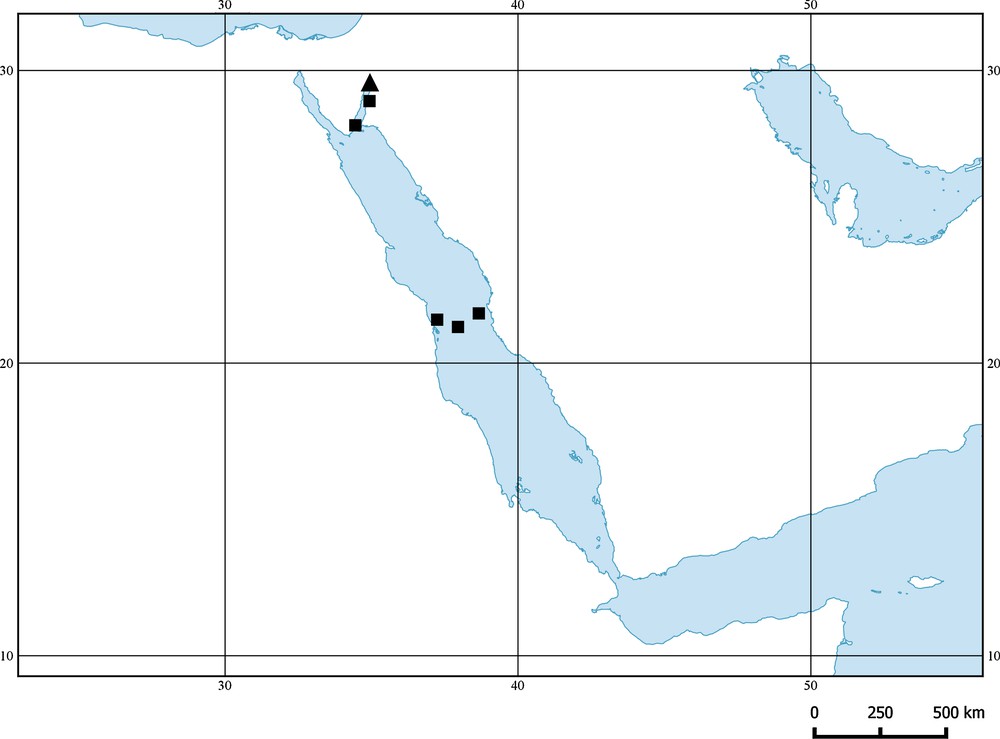
Distribution of the genus Dysomma in the Red Sea; Triangle Dysomma alticorpus new species, Square D. fuscoventralis.
3.1.5 Etymology
Altus (Latin) = high; corpus (Latin) = body. The name of the new species refers to the relatively high body of this new species. It is used as a noun in apposition.
3.1.6 Comparisons
Dysomma alticorpus n. sp. is distinguished from the similar species D. anguillare, D. fuscoventralis, and D. taiwanensis in possessing a well-developed pectoral fin, the presence of intermaxillary teeth, a uniserial row of 7–15 large compound teeth in the lower jaw (which may be followed by a few smaller teeth), and an anteriorly situated anus with the trunk shorter than the head length. It is further compared with these three similar species in Table 2. Besides several proportional differences, D. alticorpus n. sp. differs from D. anguillare in having 3 vomerine teeth (versus 4 in D. anguillare), a body relatively high, a depth at the gill opening 7.6% TL, a depth at the anus 7.8% TL (versus slenderer in D. anguillare, a depth at the gill opening 3.1–5.8% TL, a depth at the anus 2.4–6.0% TL), and the dorsal and anal fins pale, without black pigment (versus dorsal and anal fin black, with a white margin in D. anguillare). It differs from both D. fuscoventralis and D. taiwanensis in having a pale peritoneum (versus peritoneum dark), its 9 preanal lateral-line pores (versus 12–14 in D. fuscoventralis; not reaching the level of the anus in D. taiwanensis) and its 3 vomerine teeth (versus 4). The new species is further distinguished from D. fuscoventralis by its pale body (versus lower half of body dark in D. fuscoventralis), and from D. taiwanensis by its 57–58 total lateral line pores (versus 9–11 in D. taiwanensis), and its 5 infraorbital pores (versus 4 in D. taiwanensis).
Comparison of the morphometric values of four similar species of Dysomma.
| D. alticorpus n. sp. | D. anguillare Barnard 1923 | D. fuscoventralis Karrer & Klausewitz 1982 | D. taiwanensis Ho, Smith & Tighe 2015 | |
| % of TL | ||||
| Head length | 15.7 | 11.0–15.1 | 16.1 | 10.5–12.8 |
| Predorsal length | 13.8 | 10.7–13.1 | 15.5 | 11.3–12.1 |
| Preanal length | 22.8 | 15.2–18.9 | 21.1 | 13.3–16.7 |
| Trunk length | 4.1 | 2.8–5.5 | 4.8 | 2.8–3.8 |
| Tail length | 78.5 | 81.1–84.1 | 82.1 | 83.3–85.4 |
| Depth at gill opening | 7.6 | 3.1–5.8 | 5.6 | 3.4–4.7 |
| Depth at anus | 7.8 | 2.4–6.0 | 5.6 | 2.5–5.3 |
| Width at anus | 4.7 | 1.7–4.6 | 4.4 | 2.2–3.5 |
| % of HL | ||||
| Snout length | 17.6 | 17.8–23.5 | 22.5 | 22.2–26.7 |
| Eye diameter | 5.9 | 5.3–9.4 | 7.5 | 4.7–7.5 |
| Interorbital width | 14.9 | 11.5–20.4 | 15.4 | 13.9–15.1 |
| Upper-jaw length | 42.0 | 31.9–39.5 | 38.9 | 41.1–43.1 |
| Counts | ||||
| Predorsal vertebrae | 7 | 7–9 | ca. 11 | 9–10 |
| Preanal vertebrae | 16 | 12–16 | ca. 18 | 13–17 |
| Total vertebrae | 115 | 119–128 | 119–124 | 137–139 |
| Preanal lateral-line pores | 9 | 8–12 | 12–14 | Not reaching anus |
| Total lateral-line pores | 57–58 | 57–75 | 57–63 | 9–11 |
| Supraorbital pores | 3 | 3 | 3 | 3 |
| Infraorbital pores | 5 | 4–5 | 4–5 | 4 |
| Adnasal pores | 1 | 1 | 1 | 1 |
| Mandibular pores | 6 | 6 | 6 | 6 |
| Preopercular pores | 0 | 0 | 0 | 0 |
| Supratemporal pores | 0 | 0 | 0 | 0 |
| Frontal pores | 0 | 0 | 0 | 0 |
| Vomerine teeth | 3 | 4 | 4 | 4 |
| Intermaxillary teeth | 2 | 2 | 2 | 2 |
From a complex of eastern Indian Ocean/western Pacific Ocean species (D. goslinei, D. longirostrum, D. melanurum), the new species is mainly distinguished by the presence of intermaxillary teeth, and the dentary with uniserial teeth (versus intermaxillary teeth absent, and dentary with multiserial teeth); it is furthermore distinguished from these species by its short trunk, (trunk length ca. 4% TL, versus 17–19% TL in D. goslinei, 9.7–10.3 in D. longirostrum), 115 total vertebrae (versus 130–131 total vertebrae in D. goslinei, 130 in D. longirostrum, 131–138 in D. melanurum), and 57–58 total lateral-line pores (versus 27 total lateral-line pores in D. goslinei, 77–96 in D. longirostrum, 107–117 in D. melanurum).
From another complex of species (D. brevirostre, D. dolichosomatum, D. muciparus, D. tridens), the new species differs by the presence of a pectoral fin (versus pectoral absent in those species); it is further distinguished from those species by its 115 total vertebrae (versus 195–205 in D. brevirostre, 146–156 in D. dolichosomatum, 157 in D. muciparus, and 175 in D. tridens).
The new species differs from the Indo-West Pacific species D. polycatodon by its lower jaw, which is bearing a single row of 8 large compound teeth (versus 2 large compound teeth followed by row of 22–31 smaller ones in D. polycatodon), 115 total vertebrae (versus 128–133 total vertebrae in D. polycatodon), and 57–58 total lateral-line pores (versus 65–78 in D. polycatodon); it is distinguished from D. bucephalus from India (following the redescription of the holotype by Robins & Robins [10]) by its single row of 8 teeth in the lower jaw (versus 35–40 small teeth in D. bucephalus), 115 total vertebrae (versus 107 in D. bucephalus), an eye diameter 5.9% of TL (versus 1.9% of TL in D. bucephalus), and a head length 15.7% of TL (versus 10.5% of TL in D. bucephalus).
From the Taiwanese species D. opisthoproctus, the new species is distinguished by its anus situated anteriorly, and the trunk length shorter than the head length, 4.1% of TL (versus an anus situated posteriorly, a trunk length much longer than the head length, 20.2% of TL in D. opisthoproctus), preanal lateral-line pores 14 (versus 24 in D. opisthoproctus), preanal vertebrae 16 (versus 35 in D. opisthoproctus), and anal and caudal fins pale (versus distally black in D. opisthoproctus).
In order to distinguish the new species within the genus, a key to the species of Dysomma and Dysommina is presented below. Also presented below is a checklist of the species of Dysomma.
3.2 Checklist of the species of the genus Dysomma
Dysomma alticorpus new species
Present paper.
Distribution. Red Sea, Gulf of Aqaba. Depth: 350 m.
Dysomma anguillare Barnard 1923
Dysomma anguillaris Barnard 1923: Barnard [17]: 443 [off Tugela River mouth, KwaZulu-Natal, South Africa, southwestern Indian Ocean, depth 63 fathoms (115 m); holotype: SAM 12774].
Sinomyrus angustus Lin 1933: Lin [18]: 94, fig. 1 (Hoitow, Hainan, China; holotype: FESC 405, now at PRFRI collection.).
Dysomma japonicus Matsubara 1936: Matsubara [19]: 961, fig. [Kumano-Nada, southeast of the Kii Peninsula, Japan, depth 150 fathoms (274 m); holotype: FAKU 1941].
Dysomma zanzibarensis Norman 1939: Norman [20]: 44, fig. 17 (Zanzibar area, Tanzania, western Indian Ocean, 5°38’54”S, 39°15’42” to 39°17’36”E, depth 183–194 m; holotype: BMNH 1939.5.24.651).
Dysomma aphododera Ginsburg 1951: Ginsburg [21]: 452, fig. 7 [off Padre Island, Texas, U.S.A., 26°30’N, 96°26’W, depth 50 fathoms (91 m); holotype: USNM 154992].
Distribution. Circumglobal in tropical waters. Surface to 600 m in depth.
Remarks. This may be a complex of several closely related species.
Dysomma brevirostre (Facciolà 1887)
Nettastoma brevirostre Facciolà 1887: Facciolà [22]: 166, pl. 3 (fig. 3) (Sicily, Italy, Mediterranean Sea; holotype: lost).
Leptocephalus telescopius Schmidt 1913: Schmidt [23]: 11, pl. 1 (fig. 8) (Tyrrhenian Sea, Mediterranean Sea; holotype: ZMUC, probably lost).
Distribution. Mediterranean Sea, eastern and western Atlantic; Hawaiian Islands. Depths of 320–650 m.
Dysomma bucephalus Alcock 1889
Dysomma bucephalus Alcock 1889: Alcock [6]: 459 (35) [Bay of Bengal, eastern Indian Ocean, 20°17’30”N, 88°51’E, depth 193 fathoms (353 m); holotype: ZSI F11675]. Alcock [24]: pl. 6 (fig. 1).
Distribution. Indian Ocean, India. Depth of 353 m.
Dysomma dolichosomatum Karrer 1983
Dysomma dolichosomatum Karrer 1983: Karrer [9]: 93, figs. 28 A–B (Madagascar, 22°25'S, 43°04.5’E, depth 550–555 m; holotype: MNHN 1979-0004).
Distribution. Indo-West Pacific, Madagascar, Philippines, Taiwan, Solomon Islands. Depths of 105–897 m.
Dysomma fuscoventralis Karrer & Klausewitz 1982
Dysomma fuscoventralis Karrer & Klausewitz 1982: Karrer & Klausewitz [8]: 199, figs. 1–3 (central Red Sea, 21°26.50’N, 38°38.30’E, depth 799–801 m; holotype: SMF 15660).
Distribution. Red Sea. Depths of 172–1425 m.
Dysomma goslinei Robins & Robins 1976
Dysomma goslinei Robins & Robins 1976: Robins & Robins [10]: 261, figs. 3, 6, 7c, 8 (Indian Ocean, 14°52’N, 96°39’E; holotype: ANSP 133805).
Distribution. Indian Ocean, Andaman Sea; a record from Taiwan is questionable.
Dysomma longirostrum Chen & Mok 2001
Dysomma longirostrum Chen & Mok 2001: Chen & Mok [12]: 79, figs. 1A, 2A–C (Northeastern coast of Taiwan, Nan-fang-ao, depth 100–150 m, holotype: NSYU 2732).
Distribution. Western Pacific: Taiwan; Norfolk Ridge (New Caledonia). Depths of 100–540 m.
Dysomma melanurum Chen & Weng 1967
Dysomma melanurum Chen & Weng 1967: Chen & Weng [25]: 84, fig. 63 (Tungkang, southwestern Taiwan. Lectotype: NMMBP 5284).
Distribution. Western Pacific: Taiwan.
Dysomma muciparus (Alcock 1891)
Dysommopsis muciparus Alcock 1891: Alcock [26,27]: 137 [52 km southeast of Machilipatnam, Andhra Pradesh, India, Bay of Bengal, 15°56’50”N, 81°33’30”E, Investigator station 120, depth 240–276 fathoms (439–505 m); lectotype: ZSI F13107; lectotype selected by Robins & Robins 1976: 260]. Alcock [28]: pl. 34 (figs. 1, 1a).
Distribution. Indo-Pacific: Bay of Bengal, India; Hawaiian Islands. Depths of 439–505 m.
Dysomma opisthoproctus Chen & Mok 1995
Dysomma opisthoproctus Chen & Mok 1995: Chen & Mok [11]: 927, fig. 1 (off the coast of Nan-fang-ao, 24°51’24”N, 121°58’30”E, northeastern coast of Taiwan, depth 200 m; holotype: NSYSU 2701, whereabouts unknown).
Distribution. Western Pacific: Taiwan. Depth of 200 m.
Dysomma polycatodon Karrer 1983
Dysomma polycatodon Karrer 1983: Karrer [9]: 89, figs. 27 A–B (northwestern Madagascar, 15°21'S, 46°11’E, depth 170–175 m; holotype: MNHN 1979–0003).
Distribution. Indo-West Pacific: Madagascar; Philippines to Taiwan. Depths of 170–175 m.
Dysomma taiwanensis Ho, Smith & Tighe 2015
Dysomma taiwanensis Ho et al., 2015; Ho et al. [13]: 87, figs. 1–2 (Daxi, Yilan, northeast Taiwan, depth ca. 200–400 m: holotype: NMMB-P11115).
Distribution. Western Pacific: Taiwan. Depths of 200–400 m.
Dysomma tridens Robins, Böhlke & Robins in Robins & Robins 1989
Dysomma tridens Robins, Böhlke & Robins in Robins & Robins 1989: Robins & Robins [7]: 250, figs. 237–240 [off Belize, Caribbean Sea, 16°44’N, 87°55’W, depth 190 fathoms (347 m); holotype: USNM 193563].
Distribution. Western Atlantic: Caribbean Sea.
3.3 Key to the species of the genera Dysomma and Dysommina
| 1a. | Pectoral fin absent; about 2 rows of teeth on the upper jaw | 2 |
| 1b. | Pectoral fin present; 4 or more rows of teeth on upper jaw | 5 |
| 2a. | Upper jaw with three characteristic large, ventrally protruding premaxillary teeth | Dysomma tridens Robins, Böhlke & Robins 1989 |
| 2b. | Upper jaw not as described in 2a | 3 |
| 3a. | Tip of snout pointed; dorsal-fin origin about 0.8–1 head length behind the gill opening; total vertebrae 146–156 | 4 |
| 3b. | Tip of snout bulbous; dorsal-fin origin slightly behind the level of gill openings; total vertebrae 190–205 | Dysomma brevirostre (Facciolà 1887) |
| 4a. | Lateral-line pores 9–11; infraorbital pores 4 | Dysomma dolichosomatum Karrer 1983 |
| 4b. | Lateral-line pores 8; infraorbital pores 5 | Dysomma muciparus Alcock 1891 |
| 5a. | Intermaxillary teeth present; uniserial teeth on the dentary | 6 |
| 5b. | Intermaxillary teeth absent; multiserial teeth on the dentary | 12 |
| 6a. | Anus anterior, trunk shorter than the head length | 7 |
| 6b. | Anus posterior, trunk much longer than the head length | Dysomma opisthoproctus Chen & Mok 1995 |
| 7a. | Lower jaw with a series of 35–40 small teeth | Dysomma bucephalus Alcock 1889 |
| 7b. | Lower jaw with a series of 7–32 teeth in varying sizes | 8 |
| 8a. | Two large compound teeth followed by row of 22–31 smaller ones on the lower jaw | Dysomma polycatodon Karrer 1983 |
| 8b. | Single row of 7 to 15 large compound teeth on the lower jaw, maybe followed by a few smaller teeth | 9 |
| 9a. | Peritoneum dark | 10 |
| 9b. | Peritoneum whitish | 11 |
| 10a. | Total vertebrae 137–139; total lateral-line pore 40–48 | Dysomma taiwanensis Ho, Smith & Tighe 2015 |
| 10b. | Total vertebrae 119–124; total lateral-line pores 57–63 | Dysomma fuscoventralis Karrer & Klausewitz 1982 |
| 11a. | Vomerine teeth 4; body slender, depth at the gill opening 3.1–5.8% TL, depth at the anus 2.4–6.0% TL; dorsal and anal fin black, with a white margin | Dysomma anguillare Barnard 1923 |
| 11b. | Vomerine teeth 3; body relatively high, depth at the gill opening 7.6% TL, depth at the anus 7.8% TL; dorsal and anal fins pale | Dysomma alticorpus new species |
| 12a. | Anus anterior, just below the pectoral fin | 13 |
| 12b. | Anus posterior, well behind pectoral fin | 14 |
| 13a. | Lower jaw projects beyond snout, not appressed to the upper jaw when the mouth is fully closed; the lateral line extends to 1/2 head length before the base of the caudal fin | Dysomma melanurum Chen & Weng 1967 |
| 13b. | Lower jaw included, not projecting beyond snout, appressed to upper jaw when mouth fully closed; the lateral line extends anteriorly to about 23% of body | Dysomma goslinei Robins & Robins 1976 |
| 14a. | Five compound teeth on the vomer; lateral-line pores present; head pores present on frontal, infraorbital posterior to the eye and the preopercular | Dysomma longirostrum Chen & Mok 2001 |
| 14b. | Four compound teeth on vomer; pores absent on the lateral line, frontal, infraorbital posterior to the eye and the preopercular | Dysommina rugosa Ginsburg 1951 |
4 Discussion
It is notable that the new species Dysomma alticorpus co-occurs in the Gulf of Aqaba off Eilat with D. fuscoventralis, which was collected at a depth of 172 m (HUJ 17541). The latter record documents that D. fuscoventralis is not restricted to the main body of the Red Sea, as previously supposed by Karrer & Klausewitz [8] and later authors, but also occurs in the Gulf of Aqaba. Another specimen of D. fuscoventralis was collected in the Strait of Tiran at a depth of 1,450 m (HUJ 17142). The two specimens from the northern Red Sea also represent the shallowest (172 m) and the deepest records (1,450 m) for this species; the species was previously known from depths of 750–1,425 m (Karrer & Klausewitz [8]).
Like several other fish species, the new species D. alticorpus may be endemic to the Gulf of Aqaba. Other endemics include (Golani & Bogorodsky [29], Bogorodsky et al. [30], Russell [31], Fricke & Golani [32,33], Fricke et al. [34–36], Hibino et al. [37], Kovačić & Bogorodsky [38], Randall & Victor [39], Greenfield & Randall [40]; this study): Heteronarce bentuviai (Baranes & Randall 1989) (Narkidae), Gorgasia sillneri Klausewitz 1962 (Congridae), Mixomyrophis longidorsalis Hibino, Kimura & Golani 2014 and Suculentophichthus nasus Fricke, Golani & Appelbaum-Golani 2013 (Ophichthidae), Saurida golanii Russell 2011 (Synodontidae), Vinciguerria mabahiss Johnson & Feltes 1984 (Phosichthyidae), Ostichthys sufensis Golani 1984 (Holocentridae), Syngnathus safina Paulus 1992 (Syngnathidae), Scorpaenodes steinitzi Klausewitz & Frøiland 1970 (Scorpaenidae), Pterygotrigla spirai Golani & Baranes 1997 (Triglidae), Pseudanthias heemstrai Schuhmacher, Krupp & Randall 1989 (Serranidae), Pectinochromis lubbocki (Edwards & Randall 1983) and Pseudochromis fridmani Klausewitz 1968 (Pseudochromidae), Symphysanodon disii Khalaf & Krupp 2008 (Symphysanodontidae), Gymnapogon melanogaster Gon & Golani 2002 (Apogonidae), Epigonus marisrubri Krupp, Zajonz & Khalaf 2009 (Epigonidae), Atrobucca geniae Ben-Tuvia & Trewavas 1987 (Sciaenidae), Pempheris tilman Randall & Victor 2015 (Pempheridae), Upeneus davidaromi Golani 2001 (Mullidae), Chromis pelloura Randall & Allen 1982 (Pomacentridae), Stalix davidsheni Klausewitz 1985 (Opistognathidae), Trichonotus nikii Clark & von Schmidt 1966 (Trichonotidae), Limnichthys marisrubri Fricke & Golani 2012 (Creediidae), Ecsenius dentex Springer 1988 (Blenniidae), Callionymus profundus Fricke & Golani 2013 (Callionymidae), Amblyeleotris neglecta Jaafar & Randall 2009, Hazeus elati (Goren 1984), Hetereleotris psammophila Kovačić & Bogorodsky 2014, Priolepis goldshmidtae Goren & Baranes 1995, Suevita pyrios Greenfield & Randall 2017, Tomiyamichthys dorsostigma Bogorodsky, Kovačić & Randall 2011, Trimma filamentosus Winterbottom 1995, T. fishelsoni Goren 1985, and Vanderhorstia opercularis Randall 2007 (Gobiidae), Evoxymetopon moricheni Fricke, Golani & Appelbaum-Golani 2014 (Trichiuridae), Thyrsitoides jordanus Ajiad, Jafari & Mahasneh 1987 (Gempylidae), Soleichthys dori Randall & Munroe 2008 (Soleidae), Cynoglossus crepida Fricke, Golani & Appelbaum-Golani 2017 (Cynoglossidae), Canthigaster pygmaea Allen & Randall 1977 (Tetraodontidae). The total number of endemics of the Gulf of Aqaba is now as high as 38 species.
Disclosure of interest
The authors declare that they have no competing interest.
Acknowledgments
We would like to thank Albert Baranes (HUJ) who collected the holotype of the new species. We are grateful to I. Aizenberg, The Veterinary Teaching Hospital, Koret School of Veterinary Medicine, HUJ, Jerusalem for providing X-ray photographs of the holotype of Dysomma alticorpus n. sp., to David Darom (HUJ, Jerusalem, Israel) for photographs of the specimen, to Dominique Didier (ANSP), Patrice Pruvost (MNHN, Paris), and Arnold Suzumotu (BPBM) for giving access to materials under their care, and to Clea Fricke (München, Germany) for translating the abstract into French.