

1 Introduction
Situated at the western side of North Africa and receiving both Mediterranean and Atlantic influences, Morocco is today considered as a zone of high endemicity and biodiversity [1], and it represents one of the Global 200 priority ecoregions for global conservation [2]. For mammals, the last synthesis by Aulagnier et al. [4] confirmed this richness, and some recent molecular works have highlighted important intra- and interspecific rodent diversity in that region (Ndiaye et al. [5,6] for Gerbillus, Nicolas et al. [7] for Gerbillus campestris, Lalis et al. [8] for Meriones shawii).
The order Rodentia is the most diverse group of mammals, as it contains nearly half of the mammalian species. In this group, the Gerbillinae subfamily, among which the genus Gerbillus, represents an important component of the fauna of the arid and semi-arid regions in Africa and Asia [9]. However, the systematics of this subfamily and genus are still a subject of debate [6]. Some of the smallest species of the genus are considered rare or at least difficult to trap and their relationships are far from known. Furthermore, many gaps remain in our understanding of the distribution, ecology, evolution of some lesser known species in this subfamily, especially in Morocco. In this study, we present the most recent findings on two rare dwarf Gerbillinae species: Gerbillus simoni and Gerbillus henleyi.
The lesser short-tailed gerbil or Simon's gerbil (Gerbillus simoni) was first reported and described by Lataste [10], and its type locality is Oued Magra in Algeria. It was often placed into the genus Dipodillus, whom which it is the type species [11–14]. Recent molecular analyses have demonstrated that the genus is no longer valid and it is now included into Gerbillus [6,15–17]. Gerbillus kaiseri, described by Setzer [18] in Egypt and present in the East of Libya and in Egypt, is now considered as synonymous with G. simoni [14]. Another species, Gerbillus zakariai, was described by Cockrum et al. [19] as being endemic to the island of Kerkennah (Tunisia). It was later included within G. simoni by some authors [15,16,20,21], and considered as a separate insular species by others [14,19,22]. Ndiaye et al. [6] sequenced a specimen from mainland Tunisia (Kairouan) and showed that it clusters unambiguously with specimens from Kerkennah Islands. We follow Ndiaye et al. [6] and Happold [16], and consider G. zakariai as a synonym of G. simoni.
Simon's gerbil is endemic to North Africa and has a discontinuous range. The first part of its range goes from the Northeast of Morocco to the Northwest of Libya, through the North of Algeria and central Tunisia. In Algeria, it is found in the northern edge of the High Plateaus, but not in the coastal regions. The second part of the range goes from the northeastern coast of Libya to the west of the Nile Delta in Egypt [3,16]. In Morocco, this species was previously only captured by Schlitter & Setzer [23] in Oriental Morocco near Ain Beni Mathar, which represents the most western limit of this species. Simon's gerbil occurs in many habitats, especially with clay or loam soils. It is not found in sandy habitats, like many other gerbils. It inhabits lowlands and sporadically vegetated littoral desert, Chotts and salt marshes, steppe grassland vegetation (Alfa and Artemisia), steppes replaced by cropland and fallow fields, vegetated slopes and grassy valleys. It also inhabits relatively humid environments [3,4,16,24].
The Pigmy gerbil or Henley's gerbil was first reported and described by de Winton [25], and its type locality is Wadi el Natroun in Egypt. Synonyms of G. henleyi include G. mariae Bonhote, 1909, G. jordani Thomas, 1918, G. makrami Setzer, 1958 and G. syrticus Misonne, 1974, described respectively from Egypt, Algeria, Egypt and Libya [3,16,20,22,26–30].
The Pigmy gerbil lives in the Sahel, from northern Senegal to northern Sudan, and in the southern and northern edges of the Sahara. It is widespread in desert to semi-desert regions in North Africa, from Morocco to Egypt, with an extension in the Middle East and Arabia, from the East of the Sinai desert to Jordan in the North, with dispersed records in western Saudi Arabia, northeastern Qatar, northern Yemen, and Oman [3,4,21,31–33], and possibly Iran [34]. In Morocco, only few capture points have been reported until now: one in the South of Guelmim [35], several in the surroundings of Tantan and Foum el Hassan, in the Southwest [36], and one in the extreme South in Dakhla Peninsula [37]. Aulagnier et al. [4] mentioned its presence in pellets south of Tafilalt and Oued Ad Deheb close to Aousserd. The habitats of Henley's gerbil include coastal sand dunes covered with halophytes, salt marshes, gravel plains, crop fields, Wadi beds with Anabasis and Retama, stony plains and hamadas [3,33].
The most recent phylogenetic studies that were conducted on these two species are those of Abiadh et al. [15] and Ndiaye et al. [6,38]. These authors did not include Moroccan material in their study, and the overall number of specimens was low compared to the relatively large extension of their distributions.
Thanks to our new captures in Morocco, we improve the knowledge of the morphologic and genetic diversity of these poorly known dwarf gerbil species. Finally, we provide additional data on the distribution range of one of these species.
2 Material and methods
2.1 Sampling
Two field sessions in semi-arid and arid regions of the east of Morocco in October/November 2014 and May 2015 allowed us to catch five individuals of small gerbils.
In late October 2014, we have captured a small gerbil (BMT3) in the North of the Ain Beni Mathar village (North-East of Morocco: 34°07’N, 2°03’W), 23 km north of the capture site of Schlitter & Setzer [23] (33°55’N, 2°02’W), in a steppe with low vegetation in an arid region, replaced by irrigated wheat croplands, with a sandy-clay saline soil. This gerbil was initially identified as Gerbillus simoni using its external body features.
In late October and early November 2014, we also have captured three other small gerbils (BMT1, BMT2, BMT4) in the same locality. These gerbils were initially identified as Gerbillus henleyi using its body and skull features. During another field session in mid-May 2015, we have captured another small gerbil (BMT18) 29 km south of the first locality (33°52’N, 2°01’W), in a steppe environment of sandy-clay soil and with low vegetation, in an arid region. This gerbil was also initially identified as G. henleyi based upon its body and skull features. The animals were captured alive using Sherman traps with an effort of 173 trap-nights in 2014 and 266 trap-nights in 2015. The trapping success of G. simoni was 0.23% (1/439 × 100 = 0.23) and for G. henleyi was 0.91% (4/439 × 100 = 0.91). The animals were euthanized by cervical dislocation. This protocol was approved by the Cuvier (‘Muséum national d’histoire naturelle’, Paris) ethics committee. Standard external measurements like weight (WT in g), body length (HB in mm), tail length (T in mm), hind foot length (HF in mm) and ear length (E in mm) were taken. Autopsies allowed us to extract a piece of the liver for the genetic study, and the carcasses were preserved in formaldehyde. The skulls were extracted and prepared for morphometric analysis. Voucher specimens of G. simoni and G. henleyi have been deposited in the collections of the Laboratory ‘Biodiversity, Ecology and Genome’ of the Faculty of Sciences of Rabat. Voucher numbers for the individuals BMT3, BMT1, BMT2, BMT4 and BMT18 are respectively FSR-MAR14-BMT3, FSR-MAR14-BMT1, FSR-MAR14-BMT2, FSR-MAR14-BMT4, and FSR-MAR15-BMT18.
2.2 Morphometric study
For the morphometric identification of our specimens, we used the standard external measurements values, head-body length (HB), tail length (T), hind feet length (HF), ear length (E) and weight (WT), plus the ratio of the tail length to head-body length (%T). Skull measurements (mm) were taken with a Mitutoyo caliper on both dorsal and ventral view of the skull. Abbreviations of these values are as follows: greatest length of skull (GLS), breadth of braincase (BB), least interorbital constriction (IO), length of nasals (LN), length of anterior palatine foramina (LAF), length of upper molar series (M1M3), and diagonal length of tympanic bulla (LTB). Following different authors, we also calculated the proportions of the tympanic bulla length to skull length in form of a ratio (%TB).
2.3 Genetic study
The DNA of the five new specimens was extracted and purified using the QIAGEN Kit (DNeasy Blood & Tissue Kit) following the manufacturer's recommendations. Then the cytochrome b gene (1040 bp) was amplified via polymerase chain reaction (PCR) using the primers L7 (ACC AAT GAC ATG AAA AAT CAT CGT T) and H15915 (TCT CCA TTT CTG GTT TAC AAG AC) [39]. The PCR comprised an initial denaturation step of 3 min at 94 °C, followed by 38 cycles of 30 sec at 94 °C, 40 s at 52 °C, and 90 s at 72 °C, with a final extension step of 5 min at 72 °C. Double-stranded PCR products were purified and sequenced by Eurofins. Chromatograms were checked and sequences were corrected and aligned both manually and using BioEdit [40]. The sequences were then entered into the Basic Local Alignment Search Tool (BLAST, https://blast.ncbi.nlm.nih.gov/Blast.cgi) to obtain the genetic identification.
Evolutionary relationships among sequences were estimated by constructing a phylogenetic tree using the Maximum-Likelihood method (ML) in the software MEGA 7.0.26 [41]. The General time reversible (GTR) + I + G model [42] was determined by the software jModeltest 2.1.10 [43], as the best-fit model of nucleotide substitution, according to the Akaike information criterion [44]. The robustness of the obtained topologies was tested in all the treatments using 1000 bootstrap replicates. All the sequenced specimens of G. simoni and G. henleyi present in the GenBank database were included in the molecular analyses (5 G. simoni and 24 G. henleyi). Representatives of the other species of the genus Gerbillus genus were also included in the phylogenetic tree, as well as the outgroup Taterillus gracilis (Table 1).
Specimens of Gerbillus simoni, Gerbillus henleyi and other Gerbillinae used in our cyt b phylogenetic analyses.
| Species | Field or collection number | Country (locality) | Geographical coordinates | GenBank number | Reference |
| Gerbillus simoni | BMT3 | Morocco (Ain Beni Mathar) | 34°07’N, 2°03’W | MH660910 | This work |
| Gerbillus simoni | 1989024 | Tunisia (Kairouan) | ca. 35°40’N, 10°05’E | LN606694 | [6] |
| Gerbillus simoni | 1987003 | Tunisia (mainland) | – | LN606695 | [6] |
| Gerbillus simoni | 73 K | Tunisia (Kerkennah Islands) | 34°42’N, 11°11’E | GU356577 | [15] |
| Gerbillus simoni | 74 K | Tunisia (Kerkennah Islands) | 34°42’N, 11°11’E | GU356578 | [15] |
| Gerbillus simoni | 75 K | Tunisia (Kerkennah Islands) | 34°42’N, 11°11’E | GU356579 | [15] |
| Gerbillus henleyi | BMT1 | Morocco (Ain Beni Mathar) | 34°07’N, 2°03’W | MH660911 | This work |
| Gerbillus henleyi | BMT2 | Morocco (Ain Beni Mathar) | 34°07’N, 2°03’W | MH660912 | This work |
| Gerbillus henleyi | BMT4 | Morocco (Ain Beni Mathar) | 34°07’N, 2°03’W | MH660913 | This work |
| Gerbillus henleyi | BMT18 | Morocco (Ain Beni Mathar) | 33°52’N, 2°01’W | MH660914 | This work |
| Gerbillus henleyi | AD355H | Mali | – | JF704121 | [46] |
| Gerbillus henleyi | N4292 (2) | Niger (Gangara) | ca. 14°37’N, 8°30’E | JQ753050 | [47] |
| Gerbillus henleyi | M5597 | Burkina Faso (Markoye) | 14°37’N, 0°02’E | KF496220 | [38] |
| Gerbillus henleyi | M4058 | Mali (Makana) | 15°09’N, 9°29’W | KF496221 | [38] |
| Gerbillus henleyi | MAD355 | Mali (Dianbé) | 14°36’N, 5°55’W | KF496222 | [38] |
| Gerbillus henleyi | M4947 | Mali (Tedouft) | 15°55’N, 2°27’E | KF496223 | [38] |
| Gerbillus henleyi | N4291 | Niger (Gangara) | 14°37’N, 8°31’E | KF496224 | [38] |
| Gerbillus henleyi | N4292 | Niger (Gangara) | 14°37’N, 8°31’E | KF496225 | [38] |
| Gerbillus henleyi | N4293 | Niger (Gangara) | 14°37’N, 8°30’E | KF496226 | [38] |
| Gerbillus henleyi | KOR8 | Niger (Gangara) | 14°22’N, 8°18’E | KF496227 | [38] |
| Gerbillus henleyi | KOR10 | Niger (Gangara) | 14°22’N, 8°18’E | KF496228 | [38] |
| Gerbillus henleyi | N3272 | Niger (Tanout) | 14°57’N, 8°53’E | KF496229 | [38] |
| Gerbillus henleyi | AD1066 | Senegal (Dodel) | 16°28’N, 14°27’W | KF496230 | [38] |
| Gerbillus henleyi | AD2030 | Senegal (Dodel) | 16°30’N, 14°27’W | KF496231 | [38] |
| Gerbillus henleyi | KB8153 | Senegal (Dodel) | 16°30’N, 14°26’W | KF496232 | [38] |
| Gerbillus henleyi | AD1054 | Senegal (Dodel) | 16°28’N, 14°27’W | KF496233 | [38] |
| Gerbillus henleyi | AD1064 | Senegal (Dodel) | 16°28’N, 14°27’W | KF496234 | [38] |
| Gerbillus henleyi | AD1078 | Senegal (Dodel) | 16°30’N, 14°27’W | KF496235 | [38] |
| Gerbillus henleyi | AD1079 | Senegal (Dodel) | 16°30’N, 14°27’W | KF496236 | [38] |
| Gerbillus henleyi | H1 | Israel (Makhtesh Ramon) | 30°36’N, 34°50’E | KM236126 | [6] |
| Gerbillus henleyi | H2 | Israel (Makhtesh Ramon) | 30°36’N, 34°50’E | KM236127 | [6] |
| Gerbillus henleyi | H3 | Israel (Makhtesh Ramon) | 30°36’N, 34°50’E | KM236128 | [6] |
| Gerbillus henleyi | H4 | Israel (Makhtesh Ramon) | 30°36’N, 34°50’E | KM236129 | [6] |
| Gerbillus henleyi | 2002487 | Niger (Tanout) | 14°57’N, 8°53’E | LN606682 | [6] |
| Other Gerbillinae | |||||
| Gerbillus amoenus | KM236112 | ||||
| Gerbillus andersoni | LN606673 | ||||
| Gerbillus campestris | LN606674 | ||||
| Gerbillus cheesmani | KM236117 | ||||
| Gerbillus dasyurus | LN606677 | ||||
| Gerbillus floweri | KM236119 | ||||
| Gerbillus gerbillus | AJ851269 | ||||
| Gerbillus hesperinus | JN652803 | ||||
| Gerbillus hoogstrali | JN021414 | ||||
| Gerbillus latastei | GU356550 | ||||
| Gerbillus nancillus | LN606684 | ||||
| Gerbillus nanus | AJ851270 | ||||
| Gerbillus ni Geriae | AJ430555 | ||||
| Gerbillus occiduus | JN652805 | ||||
| Gerbillus perpallidus | JN652806 | ||||
| Gerbillus poecilops | JQ753064 | ||||
| Gerbillus pyramidum | JN652812 | ||||
| Gerbillus rupicola | LN606693 | ||||
| Gerbillus tarabuli | JN652825 | ||||
| Taterillus gracilis | AM409394 |
Pairwise Kimura two-parameter (K2P) genetic distances [45] were computed using MEGA 7.0.26 [41]. The K2P model was chosen for sake of comparison with other studies made on this group. The closest species to G. simoni and G. henleyi in the known phylogeny were used for comparison of the genetic distances. For G. simoni, the species G. dasyurus, G. rupicola and G. campestris of the subgenus Dipodillus were used. For G. henleyi, the species G. amoenus, G. nanus and G. poecilops of the subgenus Hendecapleura were used.
3 Results
3.1 Morphological identification
The specimen BMT3 (an adult male) was initially identified as Gerbillus simoni using the external body features. G. simoni is different from all the other gerbils due to a combination of distinctive characters – the hind feet are naked, the length of the tail is similar to the length of the head and body and the tail does not have a pencil [11,20,23].
The body and cranial measurements of this single specimen, along with the measurements found in literature, are presented in Tables 2 and 3. We can observe that the body measurements of our newly collected specimen fit within the variability of other specimens of G. simoni. Our specimen has a short tail (73 cm with a ratio of tail length to head-body length of 88%), which is a characteristic of the species G. simoni. According to literature data (Table 2), the ratio of tail length to head-body length in G. simoni is between 80 and 108% (and generally below 100%, with an average of 94%). The tail length seems to be higher towards the east (in Kerkennah Islands, Libya, and Egypt), compared to the west (Morocco, Algeria, and mainland Tunisia).
Body measurements of the newly collected specimen and comparison with measurements of Gerbillus simoni according to literature data.
| Reference | Country | Specimen | HB | T | % T | HF | E | WT |
| This work | Morocco | BMT3 | 83 | 73 | 88 | 21 | 13 | 19 |
| [10] | Algeria (specimens of the type locality) | A1 | 78 | 70 | 90 | – | – | – |
| A2 | 75 | 73 | 97 | – | – | – | ||
| [48] | Egypt | E1 | 76 | 64 | 84 | – | – | – |
| E2 | 86 | 81 | 94 | – | – | – | ||
| E3 | 86 | 83 | 97 | – | – | – | ||
| E4 | 79 | 82 | 104 | – | – | – | ||
| E5 | 87 | 85 | 98 | – | – | – | ||
| E6 | 89 | 91 | 102 | – | – | – | ||
| E7 | 82 | 85 | 104 | – | – | – | ||
| E8 | 93 | 76 | 82 | – | – | – | ||
| [49] | Algeria | Min | 70 | 57 | 81 | 20 | 11 | – |
| Max | 95 | 84 | 88 | 22 | 14 | – | ||
| [50] | Tunisia (mainland) | T1 | 75.3 | 71 | 94 | – | – | – |
| T2 | 69 | 71 | 103 | – | – | – | ||
| T3 | 71 | 69 | 97 | – | – | – | ||
| T4 | 85.8 | 73.8 | 86 | – | – | – | ||
| [29] | Libya | L1 | 81 | 83 | 102 | 21 | 12 | – |
| L2 | 88 | 81 | 92 | 21 | 12 | – | ||
| L3 | 81 | 77 | 94 | 20 | 12 | – | ||
| [23] | Morocco | M1 | 81 | 71 | 88 | 19 | 13 | 18 |
| M2 | 83 | 71 | 86 | 20 | 13 | 16 | ||
| Egypt | E9 | 85 | 92 | 108 | 22 | 13 | – | |
| E10 | 85 | 87 | 102 | 22 | 12.5 | – | ||
| Libya | Min | 81 | 71 | 88 | 21 | 12 | – | |
| Max | 85 | 84 | 99 | 21 | 13 | – | ||
| [19] | Tunisia (Kerkennah Islands) | K1 | 97 | 78 | 80 | 21 | 13 | 26 |
| K2 | 89 | 83 | 93 | 21 | 14 | 20 | ||
| K3 | 85.5 | 85.5 | 100 | – | – | – | ||
| K4 | 97 | 78 | 80 | – | – | – | ||
| K5 | 86 | 79 | 92 | – | – | – | ||
| Tunisia (mainland) | T5 | 84 | 72 | 86 | – | – | – | |
| T6 | 77.5 | 73 | 94 | – | – | – | ||
| T7 | 78 | 73 | 94 | – | – | – | ||
| T8 | 81.8 | 72.8 | 89 | – | – | – | ||
| T9 | 87.3 | 73 | 84 | – | – | – | ||
| T10 | 82 | 72 | 88 | – | – | – | ||
| T11 | 83 | 70.6 | 85 | – | – | – | ||
| [13] | Egypt | Min | 72 | 72 | 100 | 19 | 12 | 12 |
| Max | 89 | 96 | 108 | 22 | 14 | 22 | ||
| [51] | Algeria | A3 | 73.7 | 74 | 100 | 22 | 12.9 | 16.81 |
Cranial measurements of the newly collected specimen and comparison with measurements of Gerbillus simoni according to literature data.
| Reference | Country | Specimen | GLS | BB | IO | LN | LAF | M1M3 | LTB | %TB |
| This work | Morocco | BMT3 | 25.49 | 12.84 | 4.55 | 9.94 | 4.91 | 3.7 | 7.48 | 29.3 |
| [49] | Algeria | Min | 23.5 | – | – | – | – | 3.5 | – | – |
| Max | 26 | – | – | – | – | 4 | – | – | ||
| [50] | Tunisia | Min | – | 11.5 | – | – | – | – | 7.3 | – |
| Max | – | 11.7 | – | – | – | – | 8.1 | – | ||
| [29] | Libya | L1 | 25.8 | – | 5.1 | 9.6 | – | 3.5 | 8.4 | 32.6 |
| L2 | 26.3 | – | 4.7 | 9.7 | – | 3.8 | 8.6 | 32.7 | ||
| L3 | 24.4 | – | 4.6 | 9.4 | – | 3.5 | 8.1 | 33.2 | ||
| [23] | Morocco | M1 | 24.4 | – | – | 9.6 | 4.8 | 3.4 | 6.6 | 27 |
| M2 | 24.4 | – | – | 9.4 | 4.7 | 3.4 | 6.5 | 26.6 | ||
| Egypt | E1 | 25.8 | – | – | 9.5 | 4.8 | 3.5 | 7.2 | 27.9 | |
| E2 | 26 | – | – | 9.6 | 4.9 | 3.4 | 7.1 | 27.3 | ||
| Libya | Min | 24.6 | – | – | 9.1 | 4.5 | 3.2 | 6.9 | 28 | |
| Max | 26.5 | – | – | 10.3 | 5.2 | 3.4 | 7.5 | 28.3 | ||
| [19] | Tunisia (Kerkennah Islands) | K1 | 27.2 | 13.4 | 4.7 | 10.5 | 4.9 | 4 | 8.2 | 30.1 |
| K2 | 25.8 | 13 | 4.5 | 9.8 | 4.8 | 4 | 7.6 | 29.5 | ||
| Tunisia (Kerkennah Islands) | Min | 25.3 | 12.7 | 4.5 | 9.6 | 4.7 | 4 | 7.6 | 30 | |
| Max | 26.7 | 13 | 4.8 | 10.3 | 4.9 | 4 | 8.3 | 31.1 | ||
| Tunisia | Min | 24.5 | 12.7 | 4.4 | 8.9 | 4.6 | 3.3 | 7.5 | 30.6 | |
| Max | 24.7 | 12.9 | 4.6 | 9.7 | 5.3 | 3.6 | 8 | 32.4 | ||
| [13] | Egypt | Min | 23.2 | 11.5 | – | 8.4 | 4.4 | 3.2 | 6.7 | 28.9 |
| Max | 26.7 | 12.6 | – | 10 | 5.6 | 4 | 7.4 | 27.7 |
In a similar way to external body variation, we show an important variability in cranial measurements in G. simoni across its distribution in North Africa (Table 3). We can observe that the cranial measurements of our specimen fit within the variability of the other specimens of G. simoni.
The four specimens BMT1 (adult male), BMT2 (adult female), BMT4 (adult male), and BMT18 (adult female) were initially identified as Gerbillus henleyi using the body and skull features. G. henleyi is a very small gerbil with naked and unpigmented plantar soles. The tail is long, thin, and has a thin, dark and inconspicuous, about one-fourth or less of the length, terminal brush. The back coat is dark grey-brown with more or less distinct white areas behind the ear and on the rump. The ventral pelage is white and the ears are unpigmented. The skull has large tympanic bullae, compared to the size of the skull, that slightly exceed the occipital condyle [13,27]. The molars are particularly small (upper molar row ≤ 3 mm) [3,4].
The body and cranial measurements of these four specimens, along with the measurements found in the literature, are presented in Tables 4 and 5. We can observe that the body measurements of our four specimens fit within the variability of the other specimens of Gerbillus henleyi, from North Africa, the Sahel, and the Middle East, even if the %T values tend to be small in our specimens.
Body measurements of the four new specimens and comparison with measurements of Gerbillus henleyi according to literature data.
| Reference | Country | Specimen | HB | T | % T | HF | E | WT |
| This work | Morocco | BMT1 | 60 | 73 | 122 | 20 | 9 | 8 |
| BMT2 | 70 | 83 | 119 | 19 | 11 | 12.5 | ||
| BMT4 | 62 | 73 | 118 | 20 | 10 | 8.7 | ||
| BMT18 | 71 | 85 | 120 | 18 | 9 | 14 | ||
| [27] | Egypt | E1 | 74 | 98 | 132 | 21 | 9 | 10 |
| [49] | Algeria | Min | 60 | 70 | 117 | 18 | 8 | – |
| Max | 70 | 90 | 129 | 19 | 10 | – | ||
| [20] | Egypt (western desert) | Min | – | – | 102 | – | – | – |
| Max | – | – | 148 | – | – | – | ||
| Mean(N = 15) | – | – | 129 | – | – | – | ||
| Egypt (eastern desert) | Min | – | – | 129 | – | – | – | |
| Max | – | – | 159 | – | – | – | ||
| Mean(N = 8) | – | – | 140 | – | – | – | ||
| Saudi Arabia | Min | – | – | 113 | – | – | – | |
| Max | – | – | 170 | – | – | – | ||
| Mean(N = 12) | – | – | 148 | – | – | – | ||
| Duplantier 1989 (unpublished) | Senegal | S1 | 64 | 76 | 119 | 17 | 8.5 | 9 |
| S2 | 67 | 103 | 154 | 17 | 9 | 10 | ||
| [28] | Arabia, Jordan, Israel | Min | 52 | 73 | 140 | 15.8 | 7.8 | – |
| Max | 75 | 107 | 143 | 20 | 10 | – | ||
| Mean(N = 11) | 65.8 | 85.6 | – | 19 | 9 | – | ||
| [31] | Israel | Min | 60 | 93.1 | 155 | 18.2 | 8.4 | 8.01 |
| Max | 65.6 | 103.5 | 158 | 19.4 | 9.6 | 10.99 | ||
| Mean(N = 24) | 62.8 | 98.3 | – | 18.8 | 9 | 9.5 | ||
| Garba 2000 (unpublished) | Niger | N1 | 68 | 92 | 135 | 18 | 10 | 13.99 |
| [52] | Niger | Min | 64 | 85 | 133 | 18 | 9.5 | – |
| Max | 69 | 100 | 145 | 20 | 10 | – | ||
| Mean(N = 4) | 66.3 | 91.3 | – | 19.3 | 9.9 | – | ||
| [32] | Jordan | Min | 60 | 74 | 123 | 19 | 7 | 6 |
| Max | 68 | 96 | 141 | 20 | 9 | 11 | ||
| Papillon 2003 (unpublished) | Mali | M1 | 68 | 83 | 122 | 18 | 9 | 11 |
| [34] | Iran | I1 | 62 | 78 | 126 | 20 | 10 | 10 |
| [53] | Algeria | A1 | 71 | 101 | 142 | 20.48 | 10.87 | 10.41 |
| A2 | 69 | 105 | 152 | 20.44 | 10.25 | 11.67 | ||
| A3 | 78 | 104 | 133 | 19.27 | – | 10.2 | ||
| A4 | 68 | 115 | 169 | 21.15 | 9.7 | 16.66 | ||
| A5 | 79 | 108 | 137 | 20.3 | 10.98 | 22 | ||
| [54] | Israel (Agur south) | Min | – | 85 | – | 16.8 | – | 7.5 |
| Max | – | 94.5 | – | 19 | – | 9 | ||
| Israel (Agur north) | Min | – | 75 | – | 17 | – | 6.5 | |
| Max | – | 102 | – | 20 | – | 12.5 | ||
| Israel (Mifrasit) | Min | – | 83 | – | 16.1 | – | 7.6 | |
| Max | – | 99 | – | 19 | – | 14 | ||
| Israel (Mashabim) | Min | – | 82 | – | 17 | – | 7 | |
| Max | – | 97 | – | 19 | – | 10 | ||
| Israel (Mamshit) | Min | – | 84 | – | 17 | – | 7.2 | |
| Max | – | 94.5 | – | 19.5 | – | 8.7 | ||
| [55] | Algeria | A6 | 75 | 67 | 89 | 17.6 | 10 | 11.7 |
| A7 | 88 | 103 | 117 | 21.13 | 10 | 14.6 | ||
| A8 | 81 | 71 | 88 | 14.1 | 11 | 17.2 | ||
| A9 | 71 | 98 | 138 | 17.26 | 10 | 11.6 | ||
| A10 | 75 | 67 | 89 | 17.6 | 10 | 11.7 | ||
| [33] | Algeria, Tunisia, Senegal, Mali, Burkina Faso, Niger, Chad | Min | 55 | 75 | 136 | 17 | 8 | 8 |
| Max | 73 | 103 | 141 | 20 | 11 | 13.9 | ||
| Mean(N = 16) | 64.9 | 86.7 | – | 17.8 | 9.3 | 10.6 | ||
| [38] | Burkina Faso, Mali, Mauritania, Niger, Senegal | Min | 51 | 64 | 125 | 16 | 7.5 | 6.5 |
| Max | 72 | 95 | 132 | 19 | 10 | 13.9 | ||
| Mean(N = 15) | 65.47 | 84.67 | – | 17.87 | 9.21 | 10.16 |
Cranial measurements of the four new specimens and comparison with measurements of Gerbillus henleyi according to literature data.
| Reference | Country | Specimen | GLS | BB | IO | LN | LAF | M1M3 | LTB | %TB |
| This work | Morocco | BMT1 | 21.52 | 11.98 | 3.93 | 8.83 | 3.55 | 2.8 | 6.74 | 31.3 |
| BMT2 | 22.38 | 12.45 | 4.02 | 9.1 | 3.82 | 3.03 | 7.22 | 32.3 | ||
| BMT4 | 22.35 | 12.5 | 4.15 | 8.81 | 3.78 | 3.02 | 7.14 | 31.9 | ||
| BMT18 | 22.49 | 12.39 | 4.07 | 8.84 | 3.55 | 3.09 | 6.9 | 30.7 | ||
| Alluaud 1906 (unpublished) | Sudan | S1 | 22.4 | – | – | – | – | 2.9 | – | – |
| [27] | Egypt | E1 | 21.3 | – | 3.8 | – | 3.8 | 2.7 | 7.5 | 35.2 |
| [49] | Algeria | Min | 21.3 | – | – | – | – | 3 | 6.18 | 29 |
| Max | 22.2 | – | – | – | – | 3 | 7.1 | 32 | ||
| [28] | Arabia | Min | – | 10.5 | – | – | – | 2.2 | – | – |
| Max | – | 11.6 | – | – | – | 2.7 | – | – | ||
| [32] | Jordan | Min | 20.5 | 10.4 | 3.7 | – | 2.8 | 2.4 | 8.1 | 39.5 |
| Max | 22.2 | 11 | 4.2 | – | 3.7 | 2.7 | 9.2 | 41.4 | ||
| [34] | Iran | I1 | – | 11.6 | – | – | – | 3.3 | – | – |
| [53] | Algeria (Lagraff) | A1 | 23.28 | 10.76 | 4.39 | – | 3.74 | 3.55 | 7.61 | 32.7 |
| A2 | 24.72 | 11.71 | 4.66 | – | 3.8 | 3.71 | 8.64 | 35 | ||
| Algeria (Nacer) | Min | 24.82 | 10.09 | 4.34 | – | 3.72 | 3.6 | 7.85 | 31.6 | |
| Max | 25.59 | 11 | 5.74 | – | 4.05 | 3.68 | 8.29 | 32.4 | ||
| Mean(N = 3) | 25.18 | 10.61 | 4.97 | – | 3.87 | 3.64 | 8.14 | – | ||
| [55] | Algeria (Ghamra) | Min | 22.1 | 9.5 | 3.7 | 6.6 | 2.5 | 2.8 | 6.3 | 28.5 |
| Max | 24.2 | 11.2 | 5.1 | 9.1 | 3.9 | 3.8 | 8.3 | 34.3 | ||
| Algeria (Hassi Khalifa) | Min | 23 | 10.3 | 4.5 | 8.3 | 2.5 | 2.6 | 6.9 | 30 | |
| Max | 24.4 | 11.2 | 5.4 | 9.1 | 3.3 | 3 | 7.5 | 30.7 | ||
| [33] | Algeria, Tunisia, Senegal, Mali, Burkina Faso, Niger, Chad | Min | 20.5 | 10.6 | – | – | – | 2.7 | – | – |
| Max | 22.3 | 11.6 | – | – | – | 2.9 | – | – | ||
| [38] | Senegal and Niger | Min | 18.57 | 10.82 | 3.57 | – | – | 3.07 | 6.99 | 37.6 |
| Max | 22.78 | 11.78 | 4.58 | – | – | 3.41 | 8.98 | 39.4 | ||
| Mean(N = 11) | 21.35 | 11.53 | 4.12 | – | – | 3.26 | 8.31 | – |
Similarly, the skull measurements of the four newly collected specimens correspond well with the other published values for Gerbillus henleyi, except for the breadth of the braincase (BB), which appears to be larger in our specimens. This difference may be due to the different way of measuring this part of the skull by each author. Individuals from Algeria have the longest skulls and those from the Middle East (Jordan) and the Sahel (Senegal and Niger) have the longest tympanic bullas and especially the biggest ratio of tympanic bulla length to skull length.
3.2 Genetic analysis
According to the phylogenetic tree (Fig. 1), our specimen BMT3 clusters unambiguously (bootstrap value of 100%) with Tunisian specimens of Gerbillus simoni (mainland Tunisia and Kerkennah Islands). The K2P genetic distance between all the G. simoni specimens varies from 0.4 to 2.2%. The genetic distance between G. simoni and the three closest relative species (G. dasyurus, G. campestris and G. rupicola) varies from 7.7 to 14.2%.

Phylogenetic tree of Gerbillus simoni based on mitochondrial DNA resulting from the Maximum-Likelihood analysis (GTR + I + G substitution model). Numbers at nodes represent ML bootstrap support. The scale bar represents the branch length measured in the number of substitutions per site. The origin of the specimens of G. simoni is written on the right-hand side of the figure.
According to the phylogenetic tree (Fig. 2), our four specimens (BMT1, BMT2, BMT4, BMT18) cluster with Gerbillus henleyi. Our analysis allowed us to identify three lineages within this species:
- • lineage 1 groups specimens from Mali, Niger, Burkina Faso, Senegal, and Israel;
- • lineage 2 groups specimens from Mali and Niger;
- • lineage 3 contains all our specimens from Morocco.
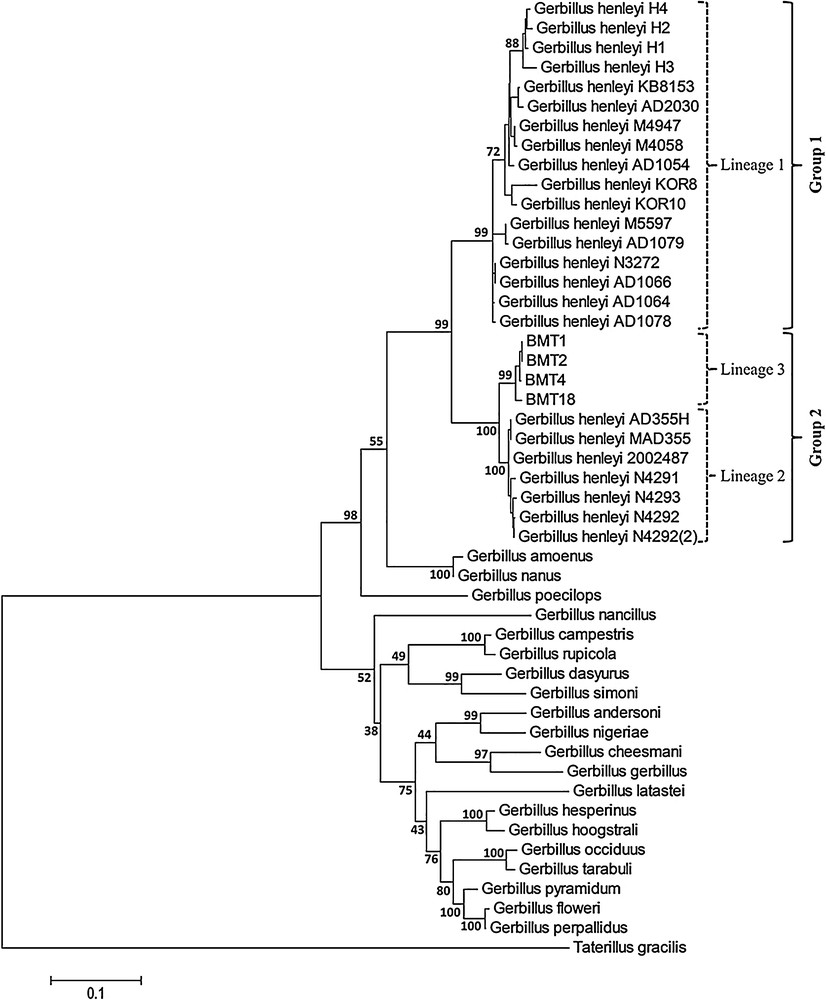
Phylogenetic tree of Gerbillus henleyi based on mitochondrial DNA resulting from the Maximum-Likelihood analysis (GTR + I + G substitution model). Numbers at nodes represent ML bootstrap support. To improve clarity, values of the most apical nodes are not included. The scale bar represents the branch length measured in the number of substitutions per site. The identified lineages of G. henleyi are on the right border.
Lineages 2 and 3 form a strongly-supported monophyletic group (Group 2).
The K2P genetic distance between these three lineages varies from 3.8% (between lineages 2 and 3) to 10.7% (between lineages 1 and 3). The genetic distance between G. henleyi and the three closest relative species (G. amoenus, G. nanus and G. poecilops) varies from 12.5 to 15.8%.
4 Discussion
Gerbillus simoni
Our specimen has a short tail with no pencil, which is a characteristic of the species Gerbillus simoni. Other species of gerbils from Morocco generally have a longer tail and the ratio of tail length to head-body length is generally comprised between 115% and 145%, in G. amoenus, G. henleyi, G. hoogstrali, G. occiduus, G. hesperinus, and G. campestris [16]. Only G. maghrebi has a relatively short tail (ratio of 95%), but it is a larger species and it displays a pencil at the tip of its tail [23]. The sequencing of the cytochrome b gene allowed us to confirm this species identification. This represents the first genetic characterization of this species in Morocco and the only one outside Tunisia [6,15].
In agreement with Cockrum et al. [19] and Osborn & Helmy [13], we observed an increase in tail length throughout North Africa from west to east. We also show an important variability in cranial measurements in G. simoni throughout North Africa. Osborn & Helmy [13] observed, in data from Algeria to Egypt, a geographic variation from west to east: decrease in the greatest width of the skull, reduced inflation of the auditory bullae, and considerable variation in colour. In our study, we could not clearly confirm this west–east variation due to the lack of sufficient data from the different countries.
Contrary to morphometric data, genetic data show little genetic variability in North Africa (less than 2.2% of sequence divergence between specimens from Morocco, mainland Tunisia, and Kerkennah Islands). Low genetic divergence between populations from north-eastern Morocco and Tunisia was also observed in other Gerbillinae rodents. For example, Lalis et al. [8] and Nicolas et al. [7], showed that Meriones shawii and Gerbillus campestris individuals from eastern Morocco belong to the same genetic clade than those present in Algeria and Tunisia.
According to Cockrum et al. [19], the population discovered on Kerkennah Islands was completely unexpected and those individuals were so morphologically distinct from the mainland samples that their relations appear best expressed by describing them as a distinct species (Gerbillus zakariai). Our genetic data show an average K2P distance of 1.0% between the specimens from Kerkennah Islands and those of mainland Tunisia. This genetic distance is much lower than the one recorded between G. simoni and its closest related species G. dasyurus (8.4%) or between clearly characterized but closely related species of the genus Gerbillus, like G. occiduus and G. tarabuli (1.8%) [5].
This low genetic divergence is another proof that these two geographically distant and isolated populations are indeed part of the same species Gerbillus simoni. The limited data concerning the karyotypes of this species reinforce this hypothesis, as they show an identical diploid number of 2n = 60. Wassif et al. [56] reported the karyotype in Egypt as 2n = 60, 8 to 10 biarms, FN = 68, 69. Karyotypes of individuals from mainland Tunisia obtained by Cockrum et al. [19] are the same (2n = 60 with 8 to 10 biarms). The first karyotype described for specimens of G. simoni from Kerkennah Islands was 2n = 60, aFN = 72 [57]. The karyotype (2n = 60, aFN = 72) was later described by Abiadh et al. [15] for the three individuals of Kerkennah Islands used in our genetic study.
Even though this species has a discontinuous distribution in North Africa, and despite the fact that it is rarely trapped, it is not considered as endangered by the IUCN. Its conservation status in Morocco should be “Not Applicable”, because it is present exclusively in a small area of the Oriental region.
Gerbillus henleyi
Our morphometric study has allowed us to identify our four specimens as Gerbillus henleyi, but a genetic confirmation was needed because the measurements of G. henleyi can be similar to those of other long-tailed small gerbils with naked hind feet, for example Gerbillus amoenus. According to Granjon [33], the latter species is slightly larger (HB = 73–96 mm, T = 99–116 mm, GLS = 24.4–26.3 mm) compared to G. henleyi (HB = 55–73 mm, T = 75–103 mm, GLS = 20.5–22.3 mm), but all samples are very small and juveniles may be confusing for identifications.
In this study, we present the first genetic characterization of this species in Morocco and North Africa, a species considered difficult to catch using classical live traps [58]. Furthermore, our study allows the extension of the known distribution of G. henleyi in Morocco 370 km to the northeast of the country, compared to the previous northernmost record that was in the south of Tafilalt from pellet remains [4].
This result could be interpreted as a possible northward extension of the geographic range of the species following desertification, as opposed to the possible southward extension of the range, in north Burkina Faso and north Senegal [33,59,60]. This species, along with desert and pre-desert Palaearctic fauna, are advancing northward due to the climatic change that touches North Africa, namely the aridification tendency. This helps in documenting the modifications in plant and animal composition of this region. Consequently, the occurrence of G. henleyi, previously unknown at this latitude in Morocco, seems to reflect a wider and perhaps continuous distribution along the northwestern border of the Sahara. It is also possible that this species was already present since a long time in this region of Morocco, characterized by an arid bioclimate, where several “southern” species extend north of the High Atlas Mountains (e.g., Paraechinus aethiopicus, Pipistrellus rueppellii, Poecilictis libyca, Jaculus, Pachyuromys duprasi, Meriones libycus, Psammomys obesus, Ctenodactylus gundi) and where “Mediterranean” species are absent (e.g., Crocidura russula, Crocidura whitakeri, Oryctolagus cuniculus, Apodemus sylvaticus, Lemniscomys barbarus). And its presence may have been unnoticed because it is a rare small gerbil, difficult to catch [54], and due to the lack of trapping campaigns and owl pellet identification. More detailed investigations in neighbouring areas can confirm whether the presence of G. henleyi in the Northeast of Morocco is exceptional or whether the recent climatic changes in North Africa favour the expansion of this species.
Our phylogeographic analyses highlighted the existence of three cyt b lineages within G. henleyi. The first is represented by individuals from Mali, Niger, Burkina Faso, Senegal, and Israel. The second contains individuals from Mali and Niger. Ndiaye et al. [6], showed a divergence as early as 1.5 Mya between these two lineages. The third lineage contains the Moroccan individuals and appears to be closer to the ‘Sahelian’ lineage 2 (3.8%) than to the ‘Sahelian–Middle Eastern’ lineage 1 (10.7%). The small genetic distance between lineages 2 and 3 may be due to a relatively recent differentiation, resulting from the last episodes of drying of the Sahara that further separated the Sahelian population from the North African population of this species. Unexpectedly, we found that the genetic distance between some geographically distant populations was low, and that the genetic distance between some neighbouring populations was high. Notably, individuals from the same locality (Gangara and Tanout in the south of Niger) can be found both in lineages 1 and 2.
More importantly, we can observe that the genetic distance between lineage 1 and lineages 2 + 3 is 10.4%. This value is far greater than the one documented between distinct species of the genus Gerbillus, like between G. occiduus and G. tarabuli (1.8%) or between G. pyramidum and G. perpallidus (3.1%) [5]. This value is also higher to what is commonly observed between sister species of rodents [61], and specifically between the two twin species G. amoenus and G. nanus (6.5%) of the subgenus Hendecapleura, that G. henleyi belongs to [47]. These preliminary genetic data (high genetic divergence in parapatric lineages) suggest the existence of two cryptic species within G. henleyi.
Cytogenetic data show that G. henleyi shares a common diploid number of chromosomes (2n = 52) with several other species of the subgenus Hendecapleura (G. poecilops, G. amoenus, and G. nanus) [62,63]. Only banding analysis can unambiguously distinguish G. henleyi (2n = 52; aFN = 58 to 62) from its sister species G. amoenus (2n = 52; aFN = 58 to 60) [52,62]. Within G. henleyi, the chromosome formula varies little despite the wide distribution of the species: aFN = 59 to 61 in Egypt [56], aFN = 58 in Morocco [35], NF = 66 and aFN = 62 in Burkina Faso [59,62], NF = 64 in Senegal [60], aFN = 62 in Tunisia [62], aFN = 59 to 62 in Niger [52,63]. The only apparent changes concern a variation of the acrocentric/metacentric proportion [59], e.g., eight bi-armed autosomes in Morocco [35], 11 to 13 in Egypt [56], 12 in Burkina Faso [59]. Similarly, in the subgenus Hendecapleura, the cytb divergence that differentiated the twin species G. amoenus and G. nanus (6.5%) was not complemented by substantial chromosomal modification, as individuals from both species had very similar karyotypes (2n = 52, aFN = 58) [47]. This weak chromosomal differentiation between the populations of G. henleyi needs to be further investigated with more specimens from the species range. Moreover, it brings us to the necessity of using nuclear DNA for a deeper understanding of the divergence between the two suggested cryptic species of G. henleyi.
Finally, the two species presented in this study were caught in the same locality and habitat along with a species of jirds and a species of jerboas. A similar situation was found in Egypt [13], where Gerbillus simoni was parapatric in salt marshes with Gerbillus henleyi, G. amoenus, G. campestris and two species of jerboas. Hence, more research must be conducted on the spatial, temporal, and trophic partitioning in these rodent communities.
Acknowledgements
The present study was supported by the Laboratory ‘Biodiversity, Ecology and Genome’ of the Faculty of Sciences of Rabat, by the UMR7205 ISYEB of the ‘Museum national d’histoire naturelle’ of Paris and by the CNRS–CNRST corporation project (2014–2015), ‘Caractérisation de la diversité génétique des rongeurs Gerbillidae du Maroc, de leur distribution et de leurs ectoparasites: implications pour la santé et la conservation’. Molecular analyses were supported by the ‘Service de systématique moléculaire’ of the ‘Museum national d’histoire naturelle’ of Paris (UMS 2700, Paris, France).