



 CC-BY 4.0
CC-BY 4.0
1. Introduction
With a global mean temperature increase of +1.4 °C relative to the pre-industrial period, combined with altered precipitation regimes and a rising frequency of extreme climatic events (IPCC, 2022), climate change is profoundly reshaping ecosystems across biomes worldwide. Europe is warming faster than the global average, and France has already experienced an increase of approximately +2.1 °C compared to pre-industrial levels (Copernicus Climate Change Service, 2024), with major consequences for forest ecosystems. Following a long period of expansion driven by increasing forest area and ecosystem fertilization—associated with rising atmospheric CO2 concentrations, nitrogen deposition, and a lengthening growing season—French forests are now increasingly exposed to the adverse effects of climate change (Ols et al., 2020). These impacts are already observable through shifts in species distributions, declining productivity, and rising rates of tree dieback across many taxa.
Climate projections indicate that these trends are likely to intensify over the 21st century, leading to substantial shifts in species’ suitable ranges. This raises critical questions about the ability of current tree species to persist, as well as about which species or populations may be better suited to future environmental conditions. Anticipating future forest composition therefore requires understanding not only species’ exposure to climate change, but also their intrinsic sensitivity and adaptive capacity. It also requires identifying those species capable of persisting or potentially replacing others under changing climatic conditions. This evaluation is an essential step for guiding forest management and conservation strategies. This paper aims to synthesize current evidence on climate-driven changes in French forests in order to identify the tree species that may shape tomorrow’s forests under ongoing and future climatic conditions.
2. Regional heterogeneity in temperature and precipitation changes in France
Across France, significant changes in temperature and precipitation have already been observed, with marked contrasts between regions and seasons. The year 2018 marks the onset of a sequence of hot and dry years, whose intensity varies depending on location (Schuldt et al., 2020). The most pronounced changes between the 1961–1990 and the 2018–2024 periods occurred during summer, with temperature increases exceeding 2.5 °C across large parts of eastern France and locally exceeding 3 °C, as illustrated in Figure 1. Summer precipitation changes show a near spatial pattern, characterized by increased precipitation along the western façade of the country and decreases in the eastern and southern regions (excepted for Corsica), accompanied by strong local heterogeneity. The combination of these temperature and precipitation changes results in contrasting trends in water availability for plants, depending on both the period and the location. Two regions appear to be particularly affected by these recent changes: northeastern France and the Mediterranean area. By contrast, Brittany, the southwest, and Corsica seem to be less affected.
3. Tree species are shifting their distributions in response to climate change
The distribution of tree species is strongly constrained by these climatic conditions. Warm, cold, or dry environments define the climatic limits within which species can establish and persist within their ecological niche (Hutchinson, 1957). In response to changes in climate, species may migrate, go extinct, or adapt. Over time, climatic fluctuations have caused shifts in species’ distribution ranges, with tree migrations. Historically, these dynamics occurred over long time scales, with species’ ranges expanding and contracting during glacial and interglacial cycles (Benito-Garzon et al., 2014). For example, deciduous Quercus species, one of the most important genera in Europe, were nearly absent from France at the end of the last glaciation but have since expanded substantially (Birks and Tinner, 2016). The current rate of climate change is unprecedented compared to these historical fluctuations, raising questions about species’ capacity to survive abrupt changes or migrate rapidly enough to track suitable habitats (Loarie et al., 2009).
In the absence of adaptation, climate change is expected to drive shifts in species distributions mainly along latitudinal and elevational gradients, potentially allowing colonization of colder margins while reducing regeneration and increasing mortality at the warm (southern) edges of species’ ranges (Lenoir and Svenning, 2015). Species distribution models (SDMs) are commonly used to estimate climatically suitable areas for future periods (Franklin, 2010; Guisan and Zimmermann, 2000). These models typically project substantial changes in suitable habitat by the end of the 21st century, with contractions for temperate and montane species and expansions for Mediterranean species across much of France (Hanewinkel et al., 2013; Mauri et al., 2022), as illustrated in Figure 2. While SDMs offer valuable insights, they assume equilibrium between species distributions and climate and often neglect demographic processes, dispersal limitations, biotic interactions, and adaptive capacity, potentially leading to an overestimation of range shifts (Buisson et al., 2010). They primarily provide long-term perspectives on potential distributional trends rather than short-term dynamics.
Evolution of summer temperatures (°C) and summer precipitation (mm) between the 1961–1990 and the 2018–2024 periods.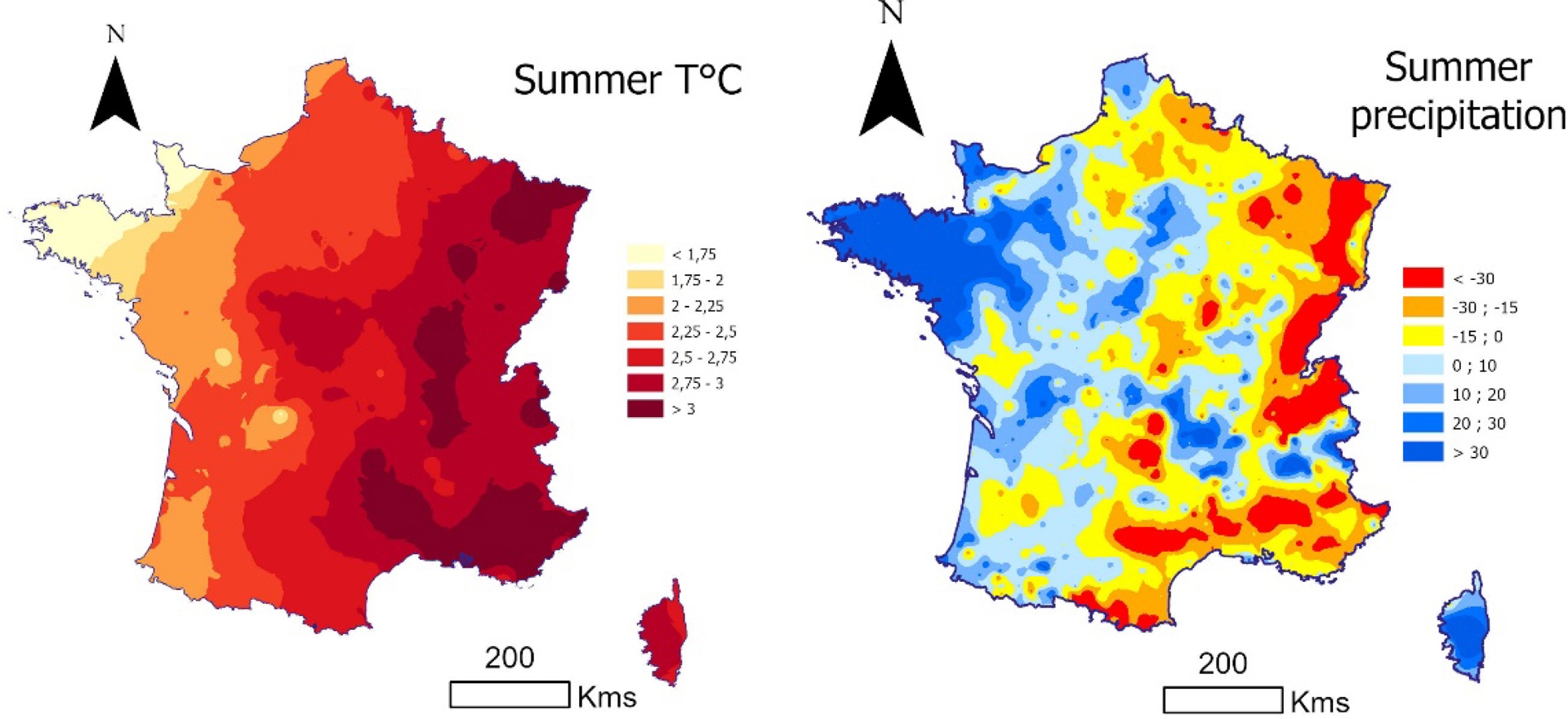
Example of the projected evolution of suitable areas for Fagus sylvatica stands (probability of presence) between the 1961–1990 period (left) and the 2071–2100 period under SSP3-7.0 (center) and SSP5-8.5 (right). Shared Socioeconomic Pathways (SSPs) are standardized scenarios describing alternative trajectories of societal development. Projections for 2071–2100 correspond to the average of three General Circulation Models (GCMs): MRI-ESM2-0, IPSL-CM6A-LR, and GFDL-ESM4.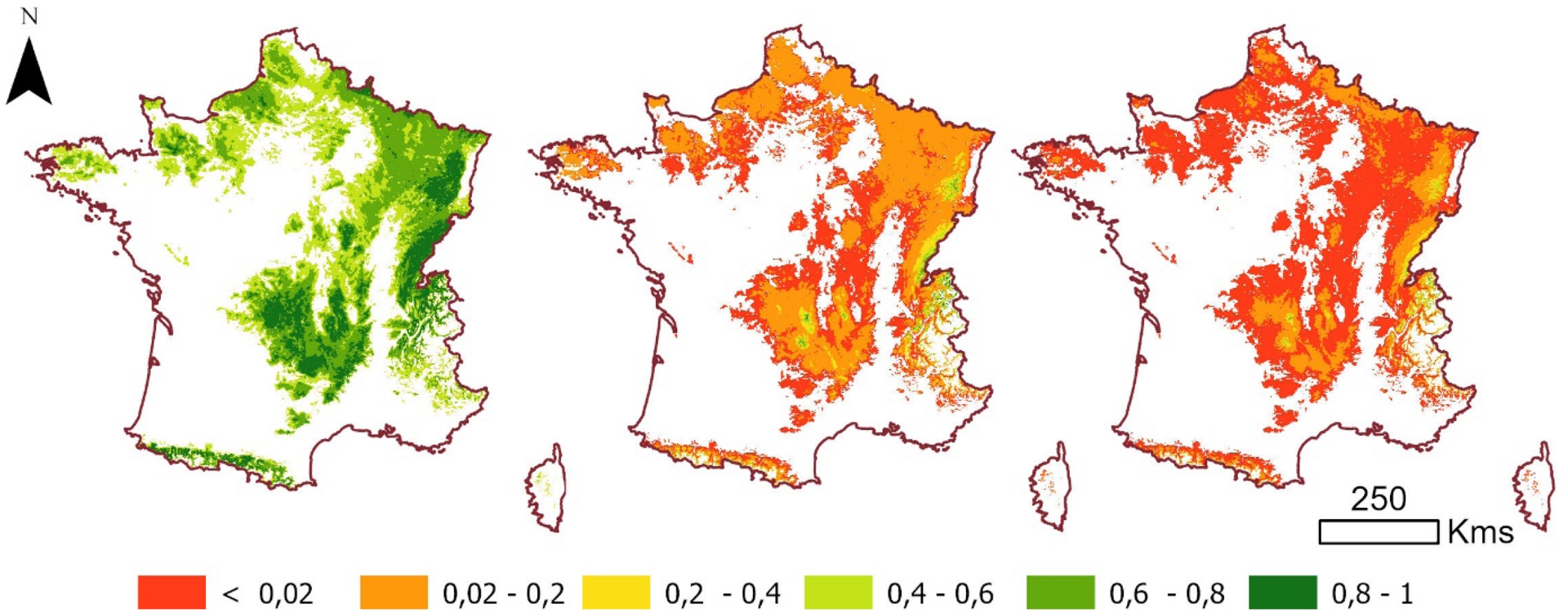
Different studies indicate that tree species’ ranges are already responding to climate warming (Rigling et al., 2013). Comparisons of juvenile and mature tree distributions show shifts toward cooler conditions at range edges in some regions (Monleon and Lintz, 2015). Some species, such as Fagus sylvatica, exhibit distributional changes along climatic gradients consistent with recent warming (Klopcic et al., 2022). These shifts are especially pronounced in mountainous regions (Crimmins et al., 2011), where species can more readily move upward than poleward. For instance, (Lenoir, Gegout, et al., 2008) reported an upward migration of approximately 29 m per decade since the beginning of the twentieth century for tree species in France. Bertrand et al. (2011) observed smaller changes in lowland areas, where longer-distance dispersal is required to maintain species within their ecological niches.
4. Climate-driven changes in tree demographic processes
Numerous ground observations illustrate that these climate changes are already affecting demographic processes (McDowell et al., 2020). Forest productivity provides an indicator of tree fitness, even in the absence of visible symptoms of decline. Recent studies have documented widespread decreases in productivity across large areas of French forests, with declines occurring earlier and more severely in Mediterranean regions than in temperate areas (Hertzog et al., 2025). In northern France, the recent decline in forest productivity was preceded by a period of accelerated growth, driven by fertilization effects and a longer growing season. In addition, climate change reduces natural forest regeneration, as heat and drought limit seedling survival and establishment (Bolte, Mansourian, et al., 2023; Kremer et al., 2025). In Mediterranean and semi-arid regions, these effects are particularly pronounced, further compromising forest resilience under ongoing climatic stress (Enríquez-de-Salamanca, 2022).
Concurrently, a marked rise in forest dieback has been observed, particularly in the center and north-east of France (Figure 3). According to estimates by the French National Forest Inventory (IGN, 2024), nearly 4% of metropolitan forests are currently affected by dieback, with the impacted area more than doubling between 2017 and 2022. These trends are consistent with observations at the European scale, indicating a marked decline in tree vitality, with increasing defoliation trends, and approximately one third of trees exhibiting moderate to severe defoliation in recent years (Hammond et al., 2022; Michel and Seidling, 2018).
Comparison of tree mortality rates across ecological regions (SER, IGN) for the major tree species in France during 2009–2017 and 2018–2024 (n = 1,321,769 trees). Plots affected by harvesting, wind, or fire damage were excluded (source: forest inventory, IGN). The year 2018 marks the beginning of a sequence of hot and dry years.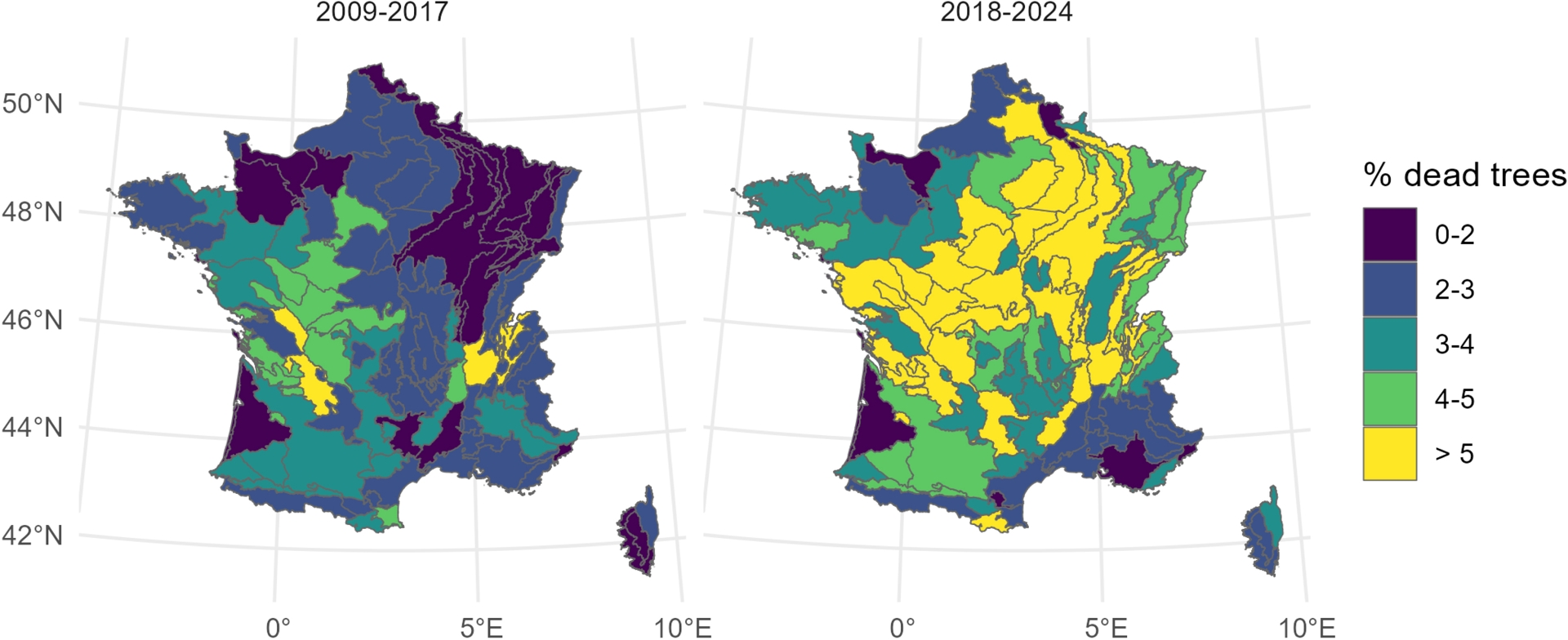
Increasing evidence indicates that tree mortality is not confined to the warm margins of species’ distribution ranges but also occurs within the core of their ecological niches (Bose et al., 2024; Cavin and Jump, 2017). Accordingly, we observe high mortality rates in northeastern France, in regions historically characterized by temperate climates and non-limiting water availability, but recently exposed to repeated climatic stresses (Figure 1). Such mortality patterns may partly reflect a phenomenon of structural overshoot, whereby forest biomass that developed under historically favorable climatic conditions exceeds the level that current environmental conditions can sustainably support (Jump et al., 2017; Y. X. Zhang et al., 2024). Conversely, limited dieback can be observed in regions with historically warm and dry climates, likely due to long-term acclimatization (Isaac-Renton et al., 2018).
These changes are influenced by stand characteristics—such as age, species composition, density, and structural complexity—which are key drivers of forest dynamics (Manion, 1981; Taccoen, Piedallu, Seynave, et al., 2019), as well as by the increasing frequency and intensity of biotic disturbances, often exacerbated by climate change (Marini et al., 2017). The interplay of these multiple factors complicates the evaluation of species vulnerability, ultimately resulting in significant ecological consequences. For example, the combined effects of productivity decrease and mortality increase have led to a marked reduction in the French forest carbon sink since the beginning of the twenty-first century (Ciais et al., 2026; CITEPA, 2023).
5. Understanding adaptive capacity of the trees is a key factor
The observed spatial patterns of forest dieback may partly reflect differences in species’ adaptive capacity, with populations originating from historically warmer or drier regions often being more resilient than those less adapted to climatic stress (Depardieu et al., 2020). Adaptive capacity refers to the ability of a species to persist and maintain its functions under changing environmental conditions, thereby mitigating the impacts of climate change. It is shaped by several key components, including phenotypic plasticity, genetic diversity, and gene flow within and among populations (Lindner et al., 2010; Valladares et al., 2014). Phenotypic plasticity enables trees to adjust their morphology, physiology, and phenology in response to altered climatic conditions—for instance, through changes in leaf traits, stomatal conductance, water-use efficiency, or the timing of budburst (Aubin et al., 2016; Nicotra et al., 2010). Plastic responses may also involve adjustments in rooting depth or biomass allocation, facilitating access to soil water during drought (Schenk and Jackson, 2002).
Genetic diversity enhances population resilience to environmental change, while connectivity among populations facilitates the exchange of traits that improve local adaptive responses (Royer-Tardif et al., 2021). However, the long lifespan of trees limits the pace of evolutionary adaptation, increasing the risk of maladaptation to changing climatic conditions (del Castillo et al., 2022). Early life stages, such as seedlings, often display higher plasticity and experience stronger selective pressures, which can promote the persistence of locally adapted genotypes under novel climatic conditions (Vitasse et al., 2013). High variability in adaptive traits complicates comparisons among species, and substantial uncertainty remains regarding the capacity of trees to respond to rapid climate change (Royer-Tardif et al., 2021). While adaptive selection may favor individuals whose traits are better suited to current environmental conditions, the overall adaptive capacity of tree populations remains challenging to quantify, particularly in natural forest ecosystems (Garzon et al., 2019).
6. Mediterranean tree species appear less affected by climate change
In this context of forest shifts, some species are expected to decline, while others may benefit (Morin et al., 2008). Comparing changes in productivity and mortality rates among species provides a useful framework to identify those with contrasting sensitivities to ongoing climatic changes. According to the IGN (IGN, 2023), the most pronounced productivity losses over the 2006–2020 period were observed for Picea abies, Pseudotsuga menziesii, and Fagus sylvatica, particularly in northeastern France (IGN, 2024). More moderate declines were reported for Quercus species, slightly more marked for Quercus robur than for Quercus petraea. In contrast, Mediterranean pines—especially Pinus pinaster and Pinus halepensis—exhibited stable or even increasing productivity during the same period (IGN, 2023).
Similar global patterns emerge when considering mortality rates between 2009 and 2024, with higher mortality observed for Picea abies and lower rates for the Mediterranean species Pinus pinaster and Pinus halepensis (Figure 4). However, notable discrepancies between productivity changes and mortality were also identified for some species. For instance, Pseudotsuga menziesii, despite experiencing substantial productivity losses (ibid.), did not show an increase in mortality in recent years (Figure 4), while Fagus sylvatica exhibited moderate mortality rates during the 2018–2024 period, despite declining productivity. Conversely, Pinus sylvestris, which displayed relatively stable productivity, showed comparatively high and increasing mortality rates. Overall, the most severe dieback occurred in species subjected to strong biotic pressures, such as Castanea sativa affected by chestnut blight and ink disease, Fraxinus excelsior impacted by ash dieback (chalara), and Picea abies, increasingly damaged by bark beetle outbreaks (Trugman et al., 2021).
Comparison of tree mortality rates between the 31 major tree species in France during 2009–2017 and 2018–2024 (n = 1,321,769 trees). Plots affected by harvesting, wind, or fire damage were excluded (source: national forest inventory, data IGN).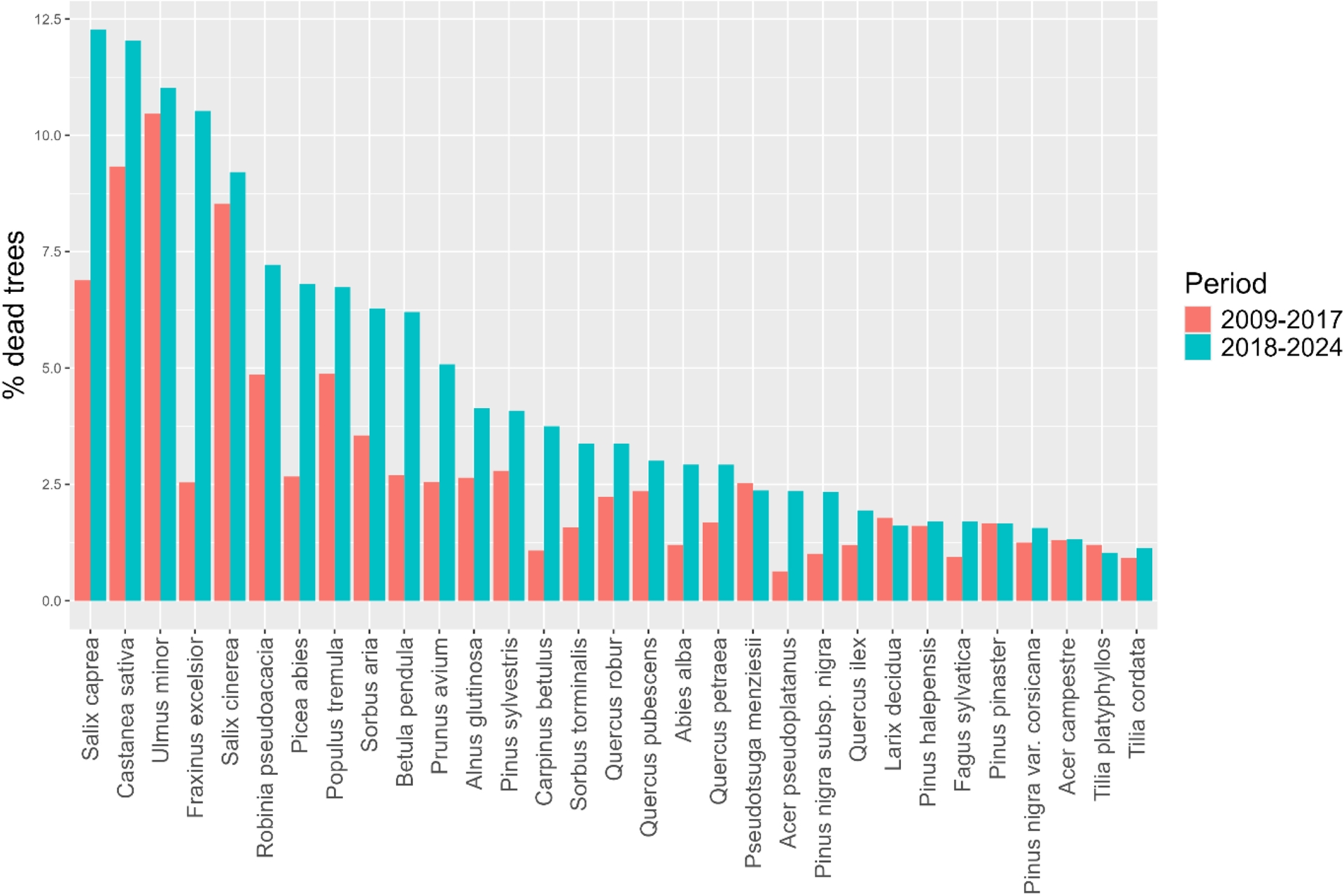
These patterns should be interpreted with caution, as they also reflect variability among stands, ecological conditions, and differences in the magnitude of climatic changes or in the intensity of pathogen outbreaks. Discrepancies between growth and mortality responses further highlight that the factors promoting tree growth may differ from those increasing mortality risk (Das et al., 2016). Nevertheless, the observed gradients support the idea of shifts in species’ distribution ranges toward higher altitudes or latitudes, with taxa originating from cool and moist environments being more disadvantaged than thermophilous species (Taccoen, Piedallu, Seynave, et al., 2022).
This comparison reveals an apparent resistance of Mediterranean tree species, especially Mediterranean pines, at the northern limit of their distribution, despite the pronounced warming observed in southern France (Figure 1). However, this resistance may be challenged in the future by the spread of emerging pathogens, such as the pine wood nematode, whose impacts can be exacerbated under warmer climatic conditions (Karmezi et al., 2022).
7. A multi-approach framework to anticipate forest responses to climate change
The significant changes in tree species distributions and demographic processes already observed are expected to persist or even intensify as climate warming continues. In this context, evaluating potential species composition to adapt forests to future climatic conditions is essential (Keenan, 2012). A wide range of tools is available to anticipate forest responses to climate change (Orazio et al., 2017). Given the complexity of forest systems and the high degree of uncertainty, we recommend combining multiple sources of information and complementary methods. A five-step methodological framework is proposed that integrates climate data, species-specific sensitivities, local environmental characteristics, and various decision-support tools to assess forest vulnerability and support adaptive management decisions at spatial scales ranging from small natural regions to the plot level.
Step 1: Characterizing past and future local climate change.
Because the impacts on vegetation dynamics generally increase with the intensity of climate change, vulnerability assessments should begin by characterizing climate changes already observed at the local scale, complemented by projections of future changes under different climate scenarios. Climate exposure should consider trends in temperature, water availability, and the frequency and intensity of extreme climatic events. The greater the magnitude and rate of changes, the higher the exposure of tree species.
Step 2: Evaluating site conditions.
A local assessment of site conditions is necessary to characterize ecological constraints. Relevant site information includes soil water-holding capacity (closely related to soil depth), soil fertility, and the occurrence of waterlogging. Certain topographic features can also be considered, as they can influence for example water fluxes and soil moisture redistribution. This information complements climate data and can be integrated with it, for example, to estimate changes in soil water availability. Site conditions are generally obtained from field surveys or existing site-type maps and may be supplemented by spatially explicit models when ground-based data are unavailable. Together, these variables refine the assessment of species exposure.
Step 3: Understanding species sensitivity.
Species sensitivity can be assessed using bibliographic information on species traits, ecological requirements, and tolerance to climatic constraints, as well as local responses to past climate events. Several databases synthesize species’ ecological traits and climatic preferences (Choat et al., 2012; Ellenberg et al., 1992; Kattge et al., 2020). In addition, indicators such as regeneration success, tree growth, or mortality rates can be assessed in the field and compared according to levels of climatic exposure. These assessments may rely on standardized protocols to describe the extent of tree decline or on dendrochronological analyses. They can be based on forest inventory data, health monitoring networks, or targeted field surveys.
Step 4: Comparison of climate-sensitive decision-support tools for forest dynamics.
Numerous decision-support tools are currently being developed to guide species selection and silvicultural practices aimed at improving forest adaptation to future climatic conditions (Cochard et al., 2021; Piedallu et al., 2023; Timberlake and Schultz, 2019). These tools span a gradient from empirical to process-based models. Empirical models rely on observed data and are primarily correlative. Among these, species distribution models are widely used to characterize climatic limits or ecological niches and to project potential distributions under future climate scenarios. If the data used to calibrate them adequately represent the ecological niche, they can be used to assess areas potentially suitable in the long term (Thuiller et al., 2005). Other common approaches focus on modeling and mapping forest dieback or tree mortality based on stand, soil, and climate variables. It provides a more realistic short-term perspective of vulnerability patterns, although extrapolating their results over longer time periods can be challenging (Piedallu et al., 2023). In contrast, process-based models explicitly represent ecological mechanisms and are commonly used to explore ecosystem responses to changing environmental conditions. Some can simulate long-term forest dynamics and assess potential impacts on regeneration, growth, or mortality (Chuine and Beaubien, 2001; Cochard et al., 2021; Ogee et al., 2003). Calibrating these models to adequately capture spatial variability can be challenging. Each modeling approach has inherent limitations and uncertainties, which should be analyzed in light of data- and method-related constraints. Although they should be compared and interpreted with caution, these different models provide essential guidance for adaptive management and for anticipating forest responses under changing climatic conditions.
Step 5: Synthesis and recommendations to estimate suitable tree species.
Assessing forest responses to climate change requires synthesizing information from these multiple sources. This synthesis could be structured through a new type of guidance framework inspired by traditional forest site-type catalogues and guides (Gaudin, 2008). This enables the evaluation of species suitability from multiple perspectives, using complementary methodological approaches. Such a synthesis requires substantial expertise due to the diversity of local contexts and the potential for contradictory results across information sources. The relevance and reliability of each source should be critically assessed, and unrealistic outcomes identified. The evaluation of suitable forest composition should also account for multiple risks, including those directly linked to climate warming and drying, as well as to forest fires, windstorms, and the emergence of novel pathogens. Although uncertainties may remain high, this integrated approach, which requires specialized expertise, can help anticipate future changes and support the design of adaptation measures tailored to local conditions.
8. A range of scenarios for future forest composition in the face of climate change
Forest composition may follow different trajectories depending on the interaction between species’ and stands’ exposure to stressors, their intrinsic sensitivity, and their adaptive capacity (Keenan, 2015) (Figure 5). When vulnerability is low, forests may persist close to their current state. As vulnerability increases, more resistant individuals or species may survive at the expense of weaker ones, potentially altering the composition of existing stands. Initially, the mortality of some individuals can benefit others by reducing competition for light and water, thereby accelerating stand regeneration dynamics. For example, seedlings established under more stressful conditions may develop traits that enhance their adaptation to future climatic conditions (X. X. Zhang et al., 2025). Conversely, if dieback is severe, the buffering effect of the forest canopy may be lost, and the microclimate may warm, limiting the success of regeneration or plantation efforts (Thom et al., 2023).
Potential adaptation pathways under increasing disturbance intensity.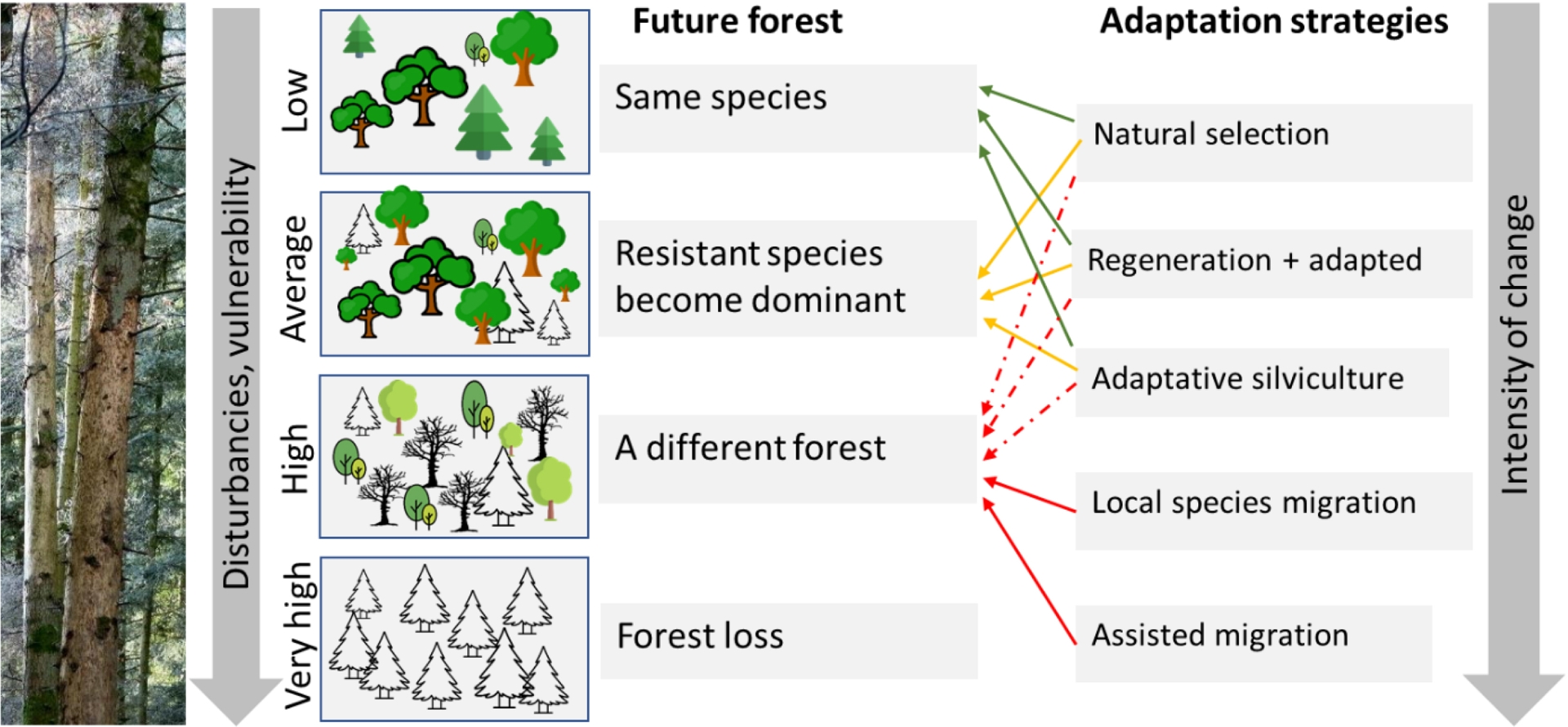
Adaptive forestry provides strategies to anticipate changes in stands that are currently minimally affected but potentially vulnerable in the future (Bolte, Ammer, et al., 2010). Key levers include stand structure adaptation, species mixture, density and water resource management, known to influence different demographic processes (Taccoen, Piedallu, Seynave, et al., 2021). Reducing basal area decreases competition and improves soil water balance by limiting rainfall interception and evapotranspiration (Sohn et al., 2016). Species mixtures diversify risks and mitigate problems from pathogen attacks, while also affecting tree interactions such as competition, facilitation, nutrient enhancement, and allelopathy (Jactel et al., 2021; Pretzsch et al., 2015). Depending on the species involved, some trees benefit from these interactions, while others may be disadvantaged, making mixture effects highly variable between species (Bose et al., 2024). Stand structure also appears particularly important, with uneven-aged stands often benefiting more (Searle et al., 2022). Additionally, water resources should be maintained or enhanced through forest practices that limit soil compaction and promote infiltration.
When species vulnerability increases to the point of threatening stand persistence, the question of species replacement arises. Several options can then be considered, such as promoting secondary species already present, encouraging their natural regeneration or arrival from nearby areas, or implementing enrichment planting. The overall objective is to increase species diversity and favor species or provenances better adapted to hot and dry climates, while minimizing reliance on artificial regeneration. When drought-tolerant species are already present nearby, they can be promoted to increase their importance within the stand. For example, the upward expansion of oak can be encouraged at higher elevations when mountain species (such as fir, spruce, or beech) are threatened by decline. Another solution is enrichment planting, which allows the diversification of an existing stand by introducing a limited number of new trees. The choice of species should be based on their tolerance for shade. For example, in northeastern France, species recommended for low-light conditions include Tilia spp., Acer spp., and Carpinus betulus, whereas Quercus robur, Pinus sylvestris, and Larix decidua are favored in areas with higher light availability (CNPF, 2024). In some situations—particularly when monospecific stands experience extensive mortality—complete reforestation may ultimately be required (Fady et al., 2021). This issue is especially acute in lowland Norway spruce forests that have experienced widespread dieback, where establishing fully stocked plantations is often required. In this case, the loss of forest cover further amplifies local temperatures and water stress, and can significantly complicate natural regeneration and the re-establishment of a continuous forest canopy.
When species better adapted to future climatic conditions are not available in the surrounding landscape, assisted migration may be considered, either by selecting new provenances of native species or by introducing non-native species with suitable ecological traits (Aitken, Yeaman, et al., 2008; Pedlar et al., 2012). Meta-analyses and common garden provenance trials have demonstrated substantial intraspecific variation in functional traits among tree populations, particularly in their responses to drought and climatic stress (Alberto et al., 2013; Copie et al., 2025). Populations originating from historically warmer or drier regions often exhibit greater drought tolerance, resistance, and survival under water-limited conditions, reflecting local adaptation of physiological traits related to water use, phenology, and hydraulic function (Isaac-Renton et al., 2018). These findings provide a scientific basis for the selection of appropriate seed sources in climate-adaptive forest management.
In France, these approaches are currently explored through experimental and operational programs such as Giono, ClimEssences, and RENEssence (Musch et al., 2022; ONF, 2022). These initiatives aim to evaluate the performance of alternative provenances and species under current and future climatic conditions. The ClimEssences platform1 (CNPF/ONF) provides detailed species and provenance profiles, including ecological traits, to guide species selection under future climate scenarios. These initiatives focus on southern or drought-tolerant provenances of native species—including, for example, Quercus species (Quercus robur, Q. petraea, Q. pubescens), Fagus sylvatica, Abies alba, and Pinus sylvestris. Experimental plantations within the Giono project have shown improved survival and drought resistance of southern provenances of Fagus sylvativa and Quercus petraea compared to local material during recent heatwaves and drought events, particularly on shallow or water-limited soils (ONF, 2022). They also target potentially suitable non-native species such as Cedrus atlantica, Abies species (Abies bornmuelleriana, cephalonica, nordmanniana, …), Quercus species (Quercus cerris or rubra), or Pinus species (Pinus radiata, taeda, pinaster, rigida, brutia), outside parts of their current distribution range.
Despite these initiatives, assisted migration remains associated with substantial uncertainties and risks. Species and provenance performance vary strongly across sites (Royer-Tardif et al., 2021). Short-term gains in growth or survival do not necessarily translate into long-term resilience, especially with respect to late frost sensitivity, pest and pathogen dynamics, or extreme events (Aitken and Bemmels, 2016). In addition, the introduction of non-native species raises concerns regarding potential impacts on native biodiversity, ecosystem functioning, and biotic interactions. Consequently, current recommendations emphasize a cautious and adaptive approach based on limited-scale trials, species mixtures, and diversification strategies rather than large-scale species replacement. Assisted migration should therefore be viewed as one component of a broader adaptation strategy, to be combined with natural regeneration, stand structural adaptation, and continuous monitoring.
9. Conclusion
Forests are already changing, reflecting the early impacts of climate change. These changes will continue as long as climate warming persists, but considerable uncertainties regarding their future trajectories. Projected climate changes suggest substantial transformations in vegetation, with a 4 °C temperature increase by the end of the 21st century potentially shifting temperate biomes toward Mediterranean-like conditions and profoundly altering species composition and ecosystem dynamics. Currently, both the southern range limits and the core areas of species’ ecological ranges appear to be affected by adverse impacts. These result from increased heat and drought, the proliferation of pathogen attacks, ecosystem degradation caused by more frequent wildfires, as well as from wind-related damage. The unprecedented pace of environmental change leaves little time for experimentation and evaluation of potential solutions, despite the critical importance of such efforts.
Assessing the composition of future forests remains challenging and will depend on various factors, including our ability to limit greenhouse gas emissions and the capacity of species to adapt to new conditions. While acclimation may provide some buffering capacity, future forest composition will also depend on our capacity to implement a diversified set of solutions, involving carefully assessed risks. Adaptive forest management can offer viable strategies by adjusting stand structure, diversifying species composition, managing density and water resources, and promoting natural regeneration, potentially delaying adverse impacts and buying valuable time for forests to adapt. Where necessary, better-adapted provenances or species may also be introduced. Despite all these possibilities, which require substantial efforts to be implemented and maintained, it is likely that, over time, a number of forests will reach silvicultural limits, and that, in some locations, forests may no longer be able to regenerate.
Evaluating potential adaptation strategies will require strong field expertise, as well as the capacity to effectively use and integrate diverse decision-support tools. Continued research is essential to better characterize vulnerable stands, assess adaptive options, and refine forest management strategies. Forests are of particular importance. They are not only ecosystems at risk but also key actors in climate change mitigation through carbon sequestration and the regulation of ecosystem processes. Preparing today’s forests for tomorrow’s climate is therefore an urgent and complex challenge that calls for immediate, informed, adaptive, and coordinated action.
Use of artificial intelligence techniques
The author declares that generative AI was used in the writing process. In accordance with the GAIDeT taxonomy (2025), the following tasks were delegated to generative AI (GAI) tools under full human supervision: Proofreading and editing, Translation. The GAI tools used were: Chat-GPT5 and DeepL. Responsibility for the final manuscript rests entirely with the authors. GAI tools are not listed as authors and bear no responsibility for the final outcomes.
Declaration of interests
The author does not work for, advise, own shares in, or receive funds from any organization that could benefit from this article, and has declared no affiliations other than their research organization.