

1 Introduction
Efficient control of plant parasites has been one of the targets of the green revolution worldwide. Since the early days when broadly generalist pesticides such as DDT were used, progress has been made in producing better-targeted products to limit the noxious effects on non-target organisms and alleviate the toxic burden on ecosystems. The result of this policy is controversial for the significant negative effects that pesticides have on ecosystem and human health [1].
At the same time, the amount of products applied has increased enormously, boosted initially by the recommendations of agricultural technicians to make systematic preventive applications. Later strategies to reduce pesticide use through development of ‘action thresholds’ – or treatments implemented beyond critical pest densities – were a step in the right direction. Yet Action Thresholds have shown in most cases to still lead to unnecessary chemical use. Far too often thresholds overestimate potential damage and underestimate or ignore negative direct and indirect effects on beneficial organisms, and human and environmental health. With thresholds the focus has remained on the plant pest, and has ignored the larger system.
In some cases also, continuous application of pesticides has selected resistant species or populations of pests, while their competitors and predators were severely hit by the toxic chemical. In such conditions control of pests requires increasing frequency and amounts of applications and direct and indirect negative effects on human health are increasing dramatically.
The continued use of pesticides has resulted in selection of genetically resistant populations of pests, and triggered resurgent outbreak of pests due to suppression of natural control factors [2–4]. Due to incomplete understanding by farmers of the phenomena of resurgence, the most common response to pesticide-induced outbreaks is the application of more pesticide, leading to the classic ‘pesticide treadmill’.
While more than 80% of pesticides are applied in developed countries, 99% of all poisoning cases occur in developing countries where regulatory, health and education systems are weakest. Faced with such evidence, FAO has edited, since 1985, an International Code of Conduct on the Distribution and Use of Pesticides and periodically updated it [1]. The Code tries to prevent health and environmental toxicity problems and promotes Integrated Pest Management (IPM) strategies that reduce the reliance on pesticides. Experience from numerous FAO projects shows that this approach offers the chance to drastically reduce the use of pesticides and to increase yields. IPM emphasises the growth of healthy crops through natural pest control systems and relies on the effective function of the natural biodiversity of the ecosystem for the regulation of pest populations.
In this paper we present three case studies, in which control by natural ecological processes has been achieved, and we discuss the implication of these results for future cropping systems that would use biodiversity for biological control.
2 Application of pesticides benefits the targeted pest [5]
In paddy rice fields of Indonesia, a number of insects may severely affect rice production. These insects develop in the rice field and their populations become important during the maturation phase of rice, creating significant damage. The dominant pest-control strategy over the past 30 years has been to select pest-resistant varieties and use chemicals to reduce populations of a small handful of pest species, without considering the linkage to the rest of the ecosystem.
Subsidised pesticide applications started in the 1960s to combat the yellow stem borer. By 1974, a new pest, the brown plant hopper Nilaparvata ligens (Stel) started to develop in many areas in spite of heavy pesticide applications, with much worse effects than those caused by the yellow stem borers. After some time, increase in pesticide applications and selection of resistant varieties were no longer able to control this pest [6]. In 1986, the Indonesian government suppressed subsidies for pesticide applications and banned 57 types of pesticides for rice. An alternative policy promoted Integrated Pest Management techniques (IPM) and 500 000 farmers were trained to become better observers and managers of their own fields to implement this new strategy. Statistics on 5000 of these farmers show that they have slightly better and less variable crop yields and better economic returns (Fig. 1).
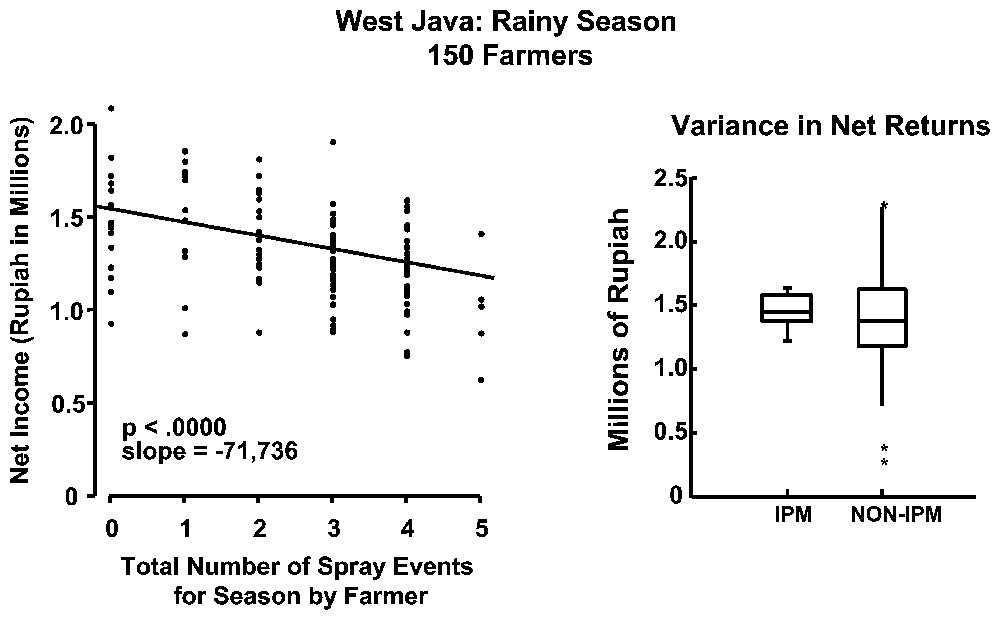
Insecticide use in irrigated rice fields in Indonesia provide increased risk and decreased return to farmers (left hand graph). Integrated Pest Management (IPM) techniques improve returns [5]. In the right-hand graph, the box represents 50% of the observations, the horizontal line inside the box is the mean, the lines coming of the box cover 95% of all observations, and the stars beyond the lines are statistical outliers.
Experiments were carried out to investigate the ecological processes that could explain these observations. The hypothesis was that generalist predators that are able to control the brown plant hopper feed on detritivores in the early phases of rice growth. They thus maintain high numbers of individuals throughout the season, allowing immediate control of the brown plant hopper as soon as their numbers start to increase. In three of six farmer's plots in Western Java, pesticides were no longer applied, whereas the other three plots received pesticide applications according to local farmer's habits.
Sampling of all the invertebrates found in the rice agro ecosystem supported the hypothesis (Fig. 2). In the no-pesticide treatment, the abundance of natural predators was almost constant during the whole growing season. When pest insects started to emerge, their control could be achieved immediately, thus preventing a significant impact on plant production.
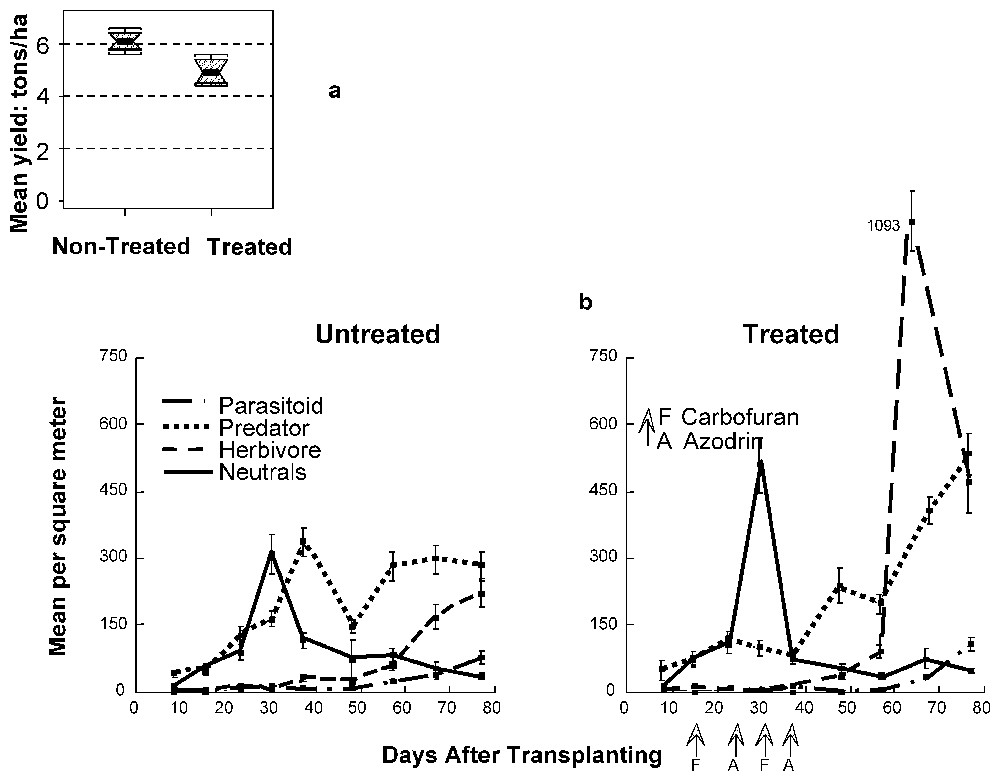
(a) Crop yield (representation as in Fig. 1) and (b) changes in abundance of different functional groups of invertebrates during the second crop of rice in Indonesian flooded rice fields treated or not with pesticides [5].
When pesticides were applied in the early stages of the cropping cycle, populations of natural predators were maintained at very low densities. These were too low for the individual species to adequately respond to increasing herbivore density. Plankton feeding populations, composed of species with high resistance to pesticides, such as Chironomidae larvae, had higher densities due to decreased predation pressure, whereas detritivores, mainly surface dwellers, were significantly depressed by pesticides. When the pest population expanded, its control was not efficient. Densities attained values 5-fold greater than in the no-pesticide treatment.
In this case a key component of pest control was the application of organic matter to sustain the development of numerous alternative prey species. The hypothesis suggests that a complex web of aquatic microorganisms develops in any flooded system. At the top of this aquatic foodweb, detritus-feeding and plankton-filtering insects function to channel the high productivity of the below-water planktonic food web to an above-surface array of generalist predators. This ubiquitous, highly-abundant and well-dispersed source of alternative prey, emerging from the water very early in the rice-growing season, effectively ‘decouples’ populations of generalist predators from the usual dependence on herbivore populations, and promotes the development of predator populations well in advance of the arrival of pest populations. Consequently, this promotes a higher likelihood of mortality for especially the early-season pest immigrants, which has a disproportionately greater impact in reducing total seasonal pest pressure on the system.
An experiment actually showed a significant increase in generalist predator populations in plots treated with organic matter. In six of 12 plots 20×20 m randomly distributed in a 1.5-ha rice field, composted cow manure was applied at a rate of 2 Mg ha−1 [5]. Rice was transplanted 10 days later and samples from on and below the water surface were taken each week for five weeks and sorted out for fauna. There was a significant increase in detritivores, plankton feeders and, ultimately, generalist predators. Although detailed mechanisms were not explored, it is hypothesised that generalist predators took advantage of increased numbers of detritivores allowed by increased availability of organic matter.
In another set of experiments, the authors showed that asynchrony between crop cycles in adjacent parcels significantly increased pest control by natural predators [7]. It took 65 days after transplanting rice in the synchronous system to reach a density of natural predators attained after only 11 days in asynchronous systems.
3 Earthworm control of plant parasitic nematodes
Soil-borne pathogens are diverse and can create severe problems for plant production. They comprise a number of fungal and bacterial pathogens, insects and nematodes. In natural ecosystems, plant diseases are generally controlled by the effect of biological trophic interactions (competition, predation and parasitism) and non-trophic interactions resulting from the effects of soil ecosystem engineers [8]. In agricultural systems, crops seem to be all the more threatened the more intensively they are cultivated. Many highly intensified plantations of banana, rubber or sugarcane are severely threatened by a number of specific pests (fungal diseases in rubber, nematodes in banana and sugarcane). Damage may, however, be important, although rarely assessed, in small-scale agriculture and even in natural ecosystems; this is for example the case with the nematode Hylonema ivorense, which impedes regeneration of Thureanthus africana in natural coastal forests of Ivory Coast and Ghana [9].
Nematodes develop in industrial plantations of such important crops as banana, sugarcane and rice and damage is estimated at over US$ 100 billion annually [10]. These parasites have a free-living phase during which interactions with other organisms may limit the density of their populations. This is, for example, the case for soil-ecosystem engineers, those soil invertebrates that influence the structure of the environment by their activities. By so doing, they also modify the availability of resources to other soil organisms [11,12].
They are represented by three major groups, viz., termites, earthworms and ants. In tropical soils, earthworms may ingest up to 20–30 times their own weight of soil daily.
Soil ingestion of the whole community of geophagous earthworm species has been estimated to be 800- to 1250-t equivalent dry soil per ha per year in a natural humid savannah of Ivory Coast [13]. This represents the equivalent of a 6- to 8-cm layer of soil and it is thus considered that the whole soil of the upper cm moves through earthworm guts at intervals of 1–2 years. Termite and ant activities are also important and it has been shown that soil artificially disaggregated, or naturally compacted by activities of a compacting invasive earthworm, could recover their original aggregated structure in less than two years in humid tropical soils [14,15].
Plant parasitic nematodes may be greatly affected by this active bioturbation [16,17].
Some of them may be ingested by invertebrates and move through their guts, with possible negative effects on their survival and the viability of eggs [18].
Ecosystem engineers also promote specific patterns in the amount, connectivity and size distribution of pores in soil, thus affecting movement of nematodes towards roots.
In laboratory experiments, we showed that control exerted by the African earthworm Millsonia anomala (Omodeo and Vaillaud) on Heterodera sacchari suppressed the negative effect of the nematodes on growth of rice (Oryza sativa) grown on a sandy ultisol. Four treatments, each replicated 6 times, were considered: a control (C) with no earthworms and nematodes, E with 4–6 earthworms, N with an initial inoculum of 2300 nematodes per pot, and EN with 4–6 earthworms and 2300 nematodes. At harvest, 90 days after the start of the experiment, average nematode abundance per pot was not statistically different in the EN (1946 ind.) and N (1526 ind.) treatments (Fig. 3a). However, the number of nematodes per unit of root biomass was 82% lower in number (Fig. 3b) since root biomass was much higher in EN than in N, and not different from non-infested treatments (Fig. 4a). Plant growth was very poor in the infested treatment (0.27 g above ground and 0.13 g below ground dry biomass per pot respectively as compared with 1.46 g and 0.45 g respectively in the control). Earthworm inoculations eliminated this effect and above ground and below ground biomass were not statistically different from the control (Fig. 4a and b).
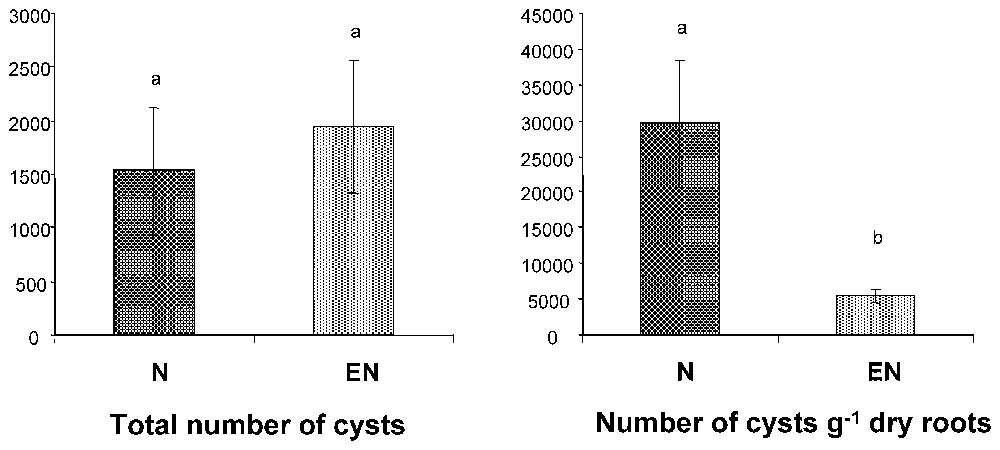
Density of the plant parasitic nematode Heterodera sacchari in laboratory cultures of rice inoculated (N) or not (EN) with earthworms (Millsonia anomala). (Left) total density; (Right) density of cysts per unit of root biomass. Treatments with different letters are significantly different (p<0.05).
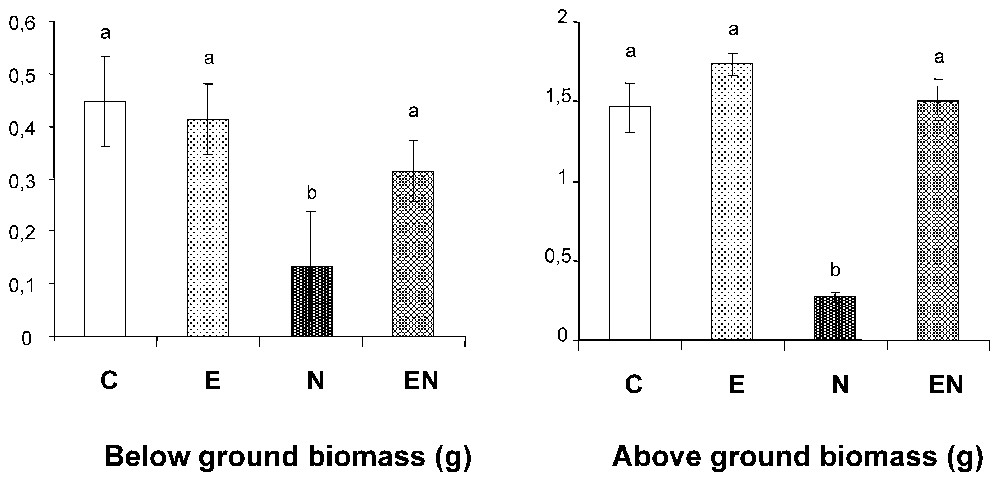
Biomass of rice plants after 90 days in different treatments: C: control; N: inoculated with parasitic nematodes (H. sacchari); E: inoculated with earthworms (M. anomala); EN: inoculated with nematodes and earthworms. Treatments with different letters are significantly different (p<0.05).
4 Diversity in nematode populations gives beneficial effects
In tropical regions, many plants are attacked simultaneously by several species of nematodes. For example, soil samples collected from the root zone of sugarcane in Ivory Coast, Chad, Burkina Faso and South Africa contained between 3 and 12 genera of plant parasitic nematodes (Table 1) [19].
Diversity of plant parasitic nematodes associated with sugarcane in individual soil samples (Cadet and Spaull, unpublished data)
| Minimum number of genera per soil sample | Maximum number of genera per soil sample | Average number of genera per sample in the survey | |
| Burkina Faso | 3 | 8 | 6.4 |
| Chad | 3 | 12 | 7.9 |
| Ivory Coast | 3 | 9 | 6.1 |
| South Africa | 3 | 9 | 5.7 |
Fallowing is a well-known cultural practice to alleviate nematode problems. Replacing the host plant by non-host or poor-host plants will inevitably lead to a reduction in the size of the populations. However, while fallowing may depress certain populations, it may increase that of other species (Fig. 5).
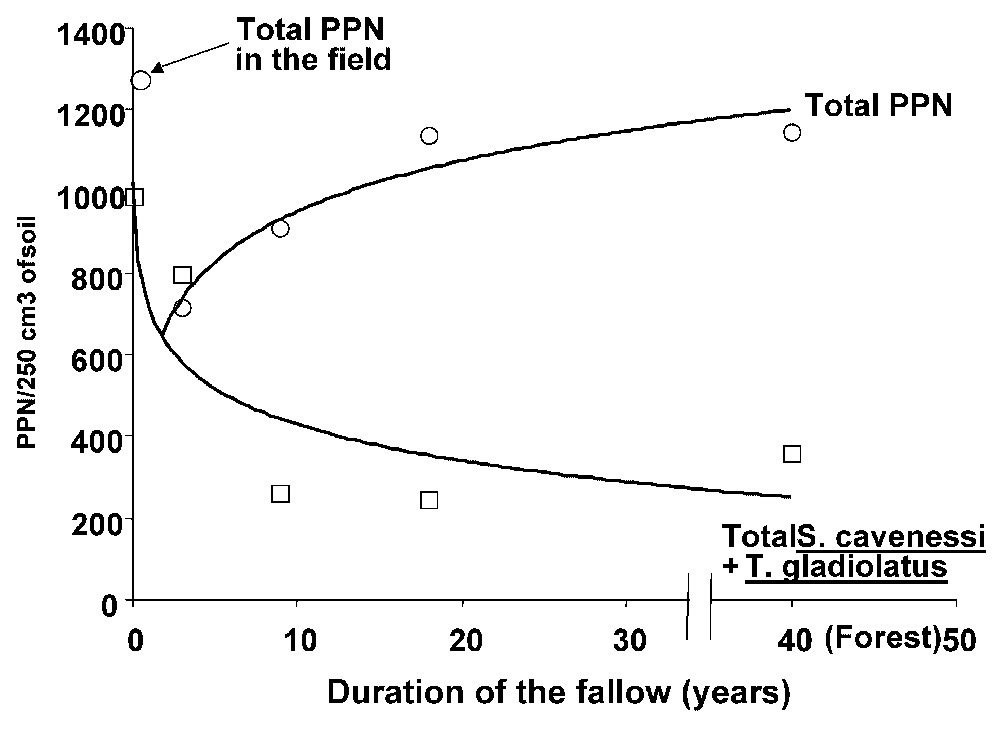
Change in total number of plant parasitic nematodes (top) and the sum of S. caveness+T. gladiolatus (bottom) according to fallow duration at Thyssé Kaymor (Senegal).
In Senegal, millet is attacked by two principal species, Tylenchorhynchus gladiolatus and Scutellonema cavenessi, which represent more than 95% of the nematode community. When millet fields were left fallow for 3, 9 or 18 years the number of T. gladiolatus and S. cavenessi decreased, while there was a marked increase in the numbers of Helicotylenchus dihystera, Pratylenchus pseudopratensis and Tylenchorhynchus mashoodi [20,21] (Table 2). All these species are able to multiply on millet [22–24] and pot experiments confirmed that all the species inoculated in isolation reduced plant and root biomass of millet [25]. However, the level of damage caused by the two principal pest species was reduced when they were associated with other species, and especially with Helicotylenchus dihystera [26]. This species seemingly mitigated the damage caused by P. pseudopratensis and T. gladiolatus.
Relative proportions (%) of the main plant-parasitic nematode genera according to fallow duration and in the fields, in the Sudanese–Sahelian region of Senegal
| Millet fields | 3 years | 9 years | 18 years | Forest | |
| Scutellonema cavenessi | 41.4 | 51.1 | 6.3 | 2.9 | 32.3 |
| Tylenchorhynchus gladiolatus | 53.5 | 36.4 | 24.5 | 18.9 | 11.2 |
| Helicotylenchus dihystera | 1.1 | 3.1 | 25.6 | 53.8 | 31.1 |
| Pratylenchus pseudopratensis | 1.2 | 1.5 | 16.9 | 12.7 | 12.6 |
| Tylenchorhynchus mashoodi | 2.6 | 7.7 | 25.4 | 10.9 | 4.5 |
| Tylenchorhynchus avaricus | 0.2 | 0.0 | 0.1 | 0.0 | 6.3 |
| Xiphinema spp. | 0.0 | 0.3 | 0.6 | 0.6 | 1.2 |
| Aphasmatylenchus variabilis | 0.1 | 0.0 | 0.6 | 0.1 | 0.8 |
The relationship was investigated by simulating successive two-month old millet crops in naturally infested soil collected from the 18-year-old fallow and from the neighbouring cultivated field. Half of each soil was sterilised. At the beginning of the experiment, the fallow soil contained five major plant parasitic species in large numbers and the cultivated soil two. On average, the root system of the plants in the infested fallow soil tended to be greater than that of the plants in the same soil that had been sterilised. The reverse was observed in the cultivated soil [27]. A separate study on the same soils confirmed that the leaf area and the root mass of the plants in the infested fallow soil, that is soil with five species of plant-parasitic nematodes, were, respectively, 43% and 34%, greater than that of the sterile fallow soil [28]. Thus, improved plant growth was associated with greater diversity of nematodes. Root proliferation in response to nematode attack has been reported for a number of plant-nematode associations [29]. The supplementary roots confer an advantage to the infested plant. They were not destroyed when the community was dominated by a wider spectrum of species, probably because the species competed with each other to occupy feeding sites. These results confirmed that a community of plant-parasitic nematodes is not always pathogenic and led to the concept of mitigating species, that is, species that are able to reduce the overall pathogenicity of a nematode community.
5 Mitigating species of nematode
This existence of mitigating species is further illustrated with data from a comparative study conducted in Burkina Faso (West Africa) and in South Africa. In both regions, which are similar, nematode susceptible sugarcane cultivars were grown on comparable sandy soils and each was attacked by nematode communities dominated by five genera: Meloidogyne spp., Pratylenchus zeae, Xiphinema spp., Paratrichodorus minor, and Helicotylenchus dihystera. These nematodes had a large and significant effect on the yield of plant cane in both localities, as measured by the response to treatment with a nematicide [30]. However, in Burkina Faso, nematodes had no significant effect on the subsequent ratoons, whereas in South Africa the damage was as great as in the plant crop (Table 3). This did not result from a decrease in yield of the treated plots in Burkina Faso, but, paradoxically, from an increase in the yield of the untreated plots, despite the presence of the same pathogenic nematode community as the year before. One of the most obvious differences was that in South Africa, the ratoon nematode community was dominated by Xiphinema spp. (71%) and in Burkina Faso, by Helicotylenchus dihystera (87%) [31]. A study of the within-field yield variation in South Africa confirmed that good growth patches of sugarcane hosted a community of plant-parasitic nematodes dominated by H. dihystera [32,33] (Figs. 6 and 7). These patches could not be explained by soil heterogeneity. This species was also the dominant plant parasitic nematode associated with the exceptionally well-grown sugarcane on levelled termitaria [20].
Response of sugarcane to nematicide treatment applied on sandy soil in South Africa and in Burkina Faso (in % over the non-treated control plots)
| Sugarcane stage | Response to nematicide treatment | |
| South Africa | Burkina Faso | |
| Plant cane | 43 | 67 |
| Ratoon 1 | 39 | 9 |
| Ratoon 2 | 68 | 0 |
| Ratoon 3 | 60 | 7 |
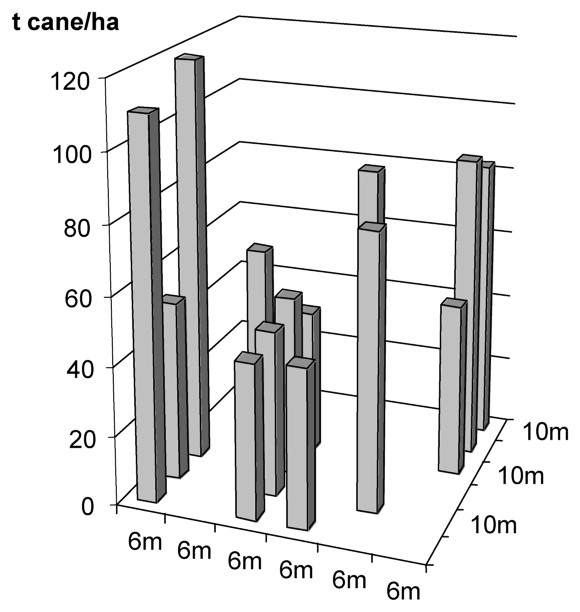
Spatial distribution of sugarcane yields on a 10×6-m grid in a field infested by plant-parasitic nematodes in South Africa.
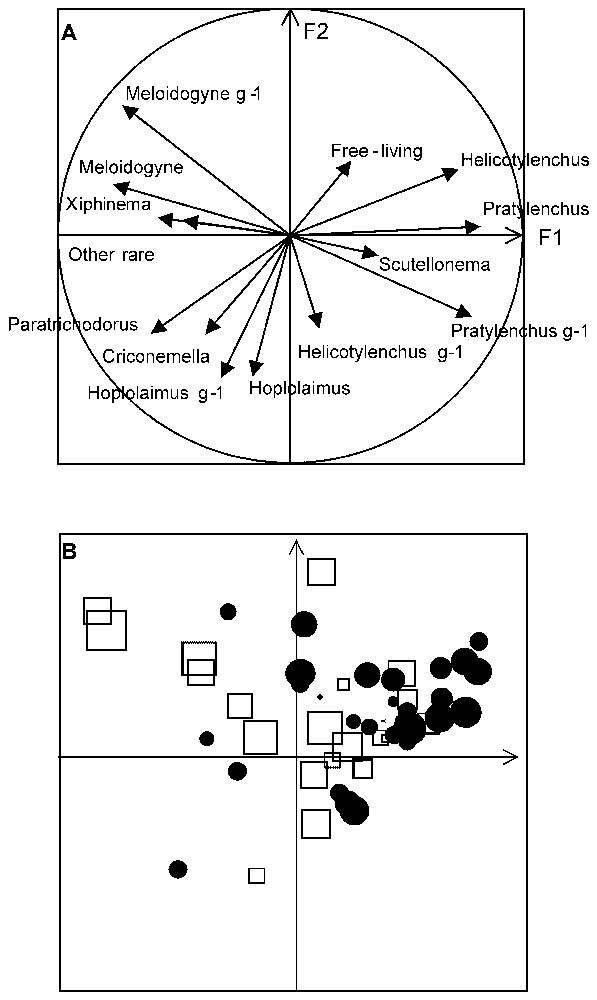
PCA performed on the relative proportions of nematode species in soil and roots measured at points located on a regular 10×6-m grid in a sugarcane plantation in South Africa. (A) Correlation of the variables (nematodes) with axes 1 and 2 of the PCA analysis. Note position of Helicotylenchus and Pratylenchus opposite to the other species. (B) Projection of the yields (normalised) of the different plots distributed on the same factorial plan according to their nematode characteristics. Yields above average are represented by circles and below average by squares. The size of the symbol is proportional to the absolute value of the normalised yield.
It is argued here that damage to plants may not be related to the number of plant parasitic individuals but rather to the species composition of the nematode community. Increasing nematode diversity by encouraging less pathogenic species to replace more pathogenic ones reduces the overall pathogenicity of the community. In other words, the management of the composition of the nematode community could achieve control of the plant pathogenic species. This alternative method of control, unlike the use of plant resistance or nematicide treatment, does not create any selection pressure or ecological vacuum, neither of which is desirable. From the agronomic point of view, the goal is not to gain the maximum, but to lose the minimum on a sustainable basis.
6 Conclusion
The different case studies presented in this paper provide convincing evidence that biological resources naturally present in agro-ecosystems may be used to reduce the effect of parasites. These resources, rather diverse in nature, comprise predaceous, decomposer and plant-parasitic species that act in different ways, but have a positive effect on plant growth.
Enhancing their effect must be part of comprehensive management strategies that aim to maintain biodiversity and high levels of activity for all these organisms. Elimination of non-target effects of pesticides is a first requisite. Adequate management of organic inputs may enhance these effects and significantly limit the need for pesticides. Finally, parasites may themselves participate in the control of their own pathogenicity. Much research is still needed to be able to generalise the concepts suggested by these case studies presented here. Experimentation is needed to check for the generality of these effects and extend the observation to microbial and fungal diseases that are known to be either stimulated or suppressed dependent on species and environmental factors. Evaluations of plant health and resistance to diseases should be developed to identify management options that allow desirable levels and qualities of plant production in environments with diverse communities in which noxious organisms are maintained at acceptable levels.
Finally, in face of the huge diversity of situations encountered in agro-ecosystems, involvement of farmers in the design of integrated pest management techniques is essential to multiply the observations and design techniques that build on scientific and indigenous knowledge, in addition to fundamental research that cannot consider the huge diversity of situations encountered in agro-ecosystems.