

Version française abrégée
Au cours de l’infection par le virus de l’immunodéficience humaine (VIH), les molécules gp120 et gp41 sont impliquées dans les processus de fixation et de fusion entre l’enveloppe du virus et celle des cellules hôtes exprimant le CD4 et les corécepteurs. La molécule gp41 transmembranaire du VIH-1 joue un rôle crucial, avec ses domaines variés, présentant un enroulement structural critique dans les changements conformationnels survenant lors de la fusion de l’enveloppe virale avec la membrane cellulaire.
Par ailleurs, il a été constaté que le déficit immunitaire lié au sida est aggravé du fait de la progression du déficit en IL-2. Cette diminution de la production d’IL-2, chez les patients atteints de sida, est manifeste. Une étude structurale comparative de la gp41 et de l’IL-2 a suggéré certaines ressemblances pouvant être à l’origine d’un mimétisme moléculaire. L’objectif de notre travail est d’analyser les similarités entre IL-2 et gp41 et surtout leurs conséquences cellulaires fonctionnelles.
La prolifération cellulaire de la lignée CTLL2, dépendant strictement de l’IL-2 en culture, peut être fortement inhibée par un anticorps monoclonal ou des anticorps polyclonaux anti-IL-2. Par ailleurs, l’addition aux cellules CTLL2 de concentrations croissantes (0,005 à 50 μg/mL) de la molécule gp41 trimérisée (loop) ou d’un peptide (séquence 578–590) de la gp41 mélangés à l’IL-2 (10 ng/mL) se traduit par une inhibition fortement significative de la prolifération en culture, même avec de faibles concentrations de la gp41 trimérisée. En revanche, l’addition de sérums de patients HIV-1 présentant un profil positif (western blot) en anticorps anti-gp41, à une culture cellulaire contenant de la gp41 et de l’IL-2 conduit à une levée de l’inhibition de la culture des cellules CTLL2.
La réactivité croisée entre gp41 et IL-2 est confirmée par le fait que le sérum anti-gp41 pré-incubé avec de l’IL-2 entraîne une inhibition forte de la capacité qu’a l’IL-2 d’induire une prolifération des cellules CTLL2 en culture.
L’infection in vitro des cellules de la lignée MT4 par le VIH-1 est fortement inhibée (production de p24) par l’addition préalable de l’IL-2.
Les données rapportées dans cette étude suggèrent fortement une réaction croisée entre les molécules de la gp41 du VIH-1 et de l’IL-2. Il s’agirait d’un mimétisme moléculaire qui contribuerait au déficit immunitaire résultant de l’interaction entre les anticorps anti-gp41 élaborés chez le patient infecté et l’IL-2.
Les recherches et les études sur la molécule gp41 trimérisée peuvent représenter l’une des approches participant aux tentatives de développement d’un vaccin anti-VIH.
1 Introduction
During the course of human immunodeficiency (HIV) infection, the gp120 and gp41 molecules are involved in the binding and fusion processes between the membrane of the virus and the membrane of those host cells that express CD4 and coreceptors [1]. The transmembrane protein (gp41) of HIV-1 contains various domains, which play a key role, particularly in the amino-acid sequence 578–590, a portion that has been modeled in a coiled-coil (leucine zipper-like) structure that may be critical for fusion-related conformational changes [2–4].
In addition, it has been observed that the immune deficiency of AIDS is aggravated when IL-2 deficiency progresses [5]. A decrease in IL-2 production has regularly been found in AIDS patients [6]. A structural study has suggested mimicry between gp41 and IL-2 [7,8].
The objective of this article is to analyze whether similarities between IL-2 and gp41 have cellular and functional consequences. The competition between IL-2 and HIV-1 has been tested on MT4 cells. The capacity of a peptide to inhibit proliferation of CTLL2 cells by blocking the IL-2 receptor has also measured.
2 Materials and methods
2.1 Effects of anti-IL-2 antibodies on cell culture
IL-2-dependent cells (CTLL2 cell line) were maintained in culture in the continuous presence of human IL-2 to determine biological activity of the cytokine. Before culture, 200 μL of a cell suspension (2 × 104 cells) were dispensed in wells of a 96-well microplate and various concentrations (0.001 to 100 ng/mL) of IL-2 were tested (Fig. 1a).

Modification of CTLL2 proliferation by IL-2 or anti-IL-2 antibodies. a: Effect of IL-2 on CTLL2 proliferation (). b: Effect of anti-IL-2 antibodies on CTLL2 proliferation. Anti-IL-2 monoclonal (MAB202 ) or polyclonal (202NA ) antibodies and monoclonal anti-CD19 () or polyclonal anti-albumin () antibodies.
IL-2 effect was assayed on CTLL2 cell proliferation in the presence of mouse anti-IL-2 monoclonal antibody (MA 202, R&D, Abingdon, UK), goat anti-IL-2 polyclonal antibody (AF-202-NA, R&D, Abindon, UK) as potential inhibitors, and in the presence of goat anti-albumin (gift of Dr. Serres, Lyon, France) or mouse monoclonal anti-CD19 (Becton Dickinson, Meylan, France) as control antibodies. These antibodies were used at concentrations ranging from 0.0005 to 50 μg/mL.
2.2 Effect of IL-2 on HIV-1 infection in vitro
MT4 cells (human T4 cell line expressing IL-2 receptor) [9] were maintained in culture in RPMI-1640 medium. For IL-2 fixation and HIV-1 binding onto targets, cells were plated at 3 × 105 cells per well and were immediately pre-incubated for 15 minutes at +4 °C (to avoid IL-2 cell internalization) in the presence of human IL-2 (Roche Diagnostics, Meylan, France) at optimal concentrations of 10 ng/mL (titration of IL-2 was carried out on CTLL2 cell line cultures). Then, HIV-1 (Lai strain) was added at 0.001 MOI for 45 minutes at 37 °C. Excess virus and IL-2 were removed by extensive washing. Cells were then cultured at 37 °C in a 5% CO2–95% air atmosphere. Cultures were maintained and supernatants were collected every 2–3 days to test for HIV-1 p24 release (ELISA) and analysis of cytopathogenic effects.
2.3 In vitro competition between gp41 molecule and IL-2
Several forms of HIV-1 gp41 molecule were tested. Peptide #1 (578–590 amino-acid sequence of gp41) and peptide # 2 (587–599 amino-acid sequence of gp41) (ANRS, Paris, France), whole synthetic gp41 molecule (ProteinExpert, Grenoble, France) under trimeric (loop) or linear (HA) forms, at 0.005, 0.05, 0.5, 5 and 50 μg/mL, were pre-incubated for 1 hour with IL-2 at 10 ng/mL. The mixture was then added to cultures of CTLL2 for 2 days.
To analyze the specificity of the competition between gp41 molecules and IL-2, the various forms of gp41 molecules, at different concentrations, were mixed with IL-2 (10 ng/mL). The HIV-1 anti-gp41 serum (at the 1/10 optimal dilution) were added to the cultures of CTLL2. The anti-gp41 serum was obtained by collecting 32 sera obtained from consenting HIV-1 seropositive non-treated patients. The pooled sera were previously HIV-inactivated by heating at 56 °C for 30 minutes. Anti-gp41 antibodies contained in patients’ sera were detected by western blot technique (Genelabs Diagnostics, USA). Individual sera were positive for p24, p31, gp41 and gp120/gp160 antibodies as tested on antigens coated on strips and showed various degrees of inhibition in presence of IL-2 on CTLL2 cultures. All 32 sera were then pooled and used in the reported experiments. Eight negative HIV-1 sera were tested and used as control in the same experimental conditions.
To check for cross-reactivity between IL-2 and gp41, IL-2 (10 ng/mL) was mixed with anti-gp41 serum at 1/80, 1/40, 1/20 and 1/10 dilutions and added to CTLL2 cultures.
Eighteen hours before the end of cultures, 1 μCi [3H]thymidine (ICN, UK) was added to each well. Cells were collected using a cell harvester (Skatron, UK). Cell proliferation was evaluated by incorporation of [3H]thymidine in the DNA of CTLL2 cells. Cell viability was monitored in cultures at 24 and 48 hours using the Trypan blue exclusion assay.
3 Results
3.1 Antiproliferative effects of anti-IL-2 antibodies
Biological activity of IL-2 was previously determined for further assays. As shown in Fig. 1a, IL-2 was tested in culture on IL-2-dependent CTLL2 cell line. The mean of three independent experiments showed that 10 ng/mL of IL-2 resulted in a significant optimal dose in our experiments.
As shown in Fig. 1b, both anti-IL-2 antibodies demonstrated a significant inhibition of IL-2-dependent CTLL2 proliferation in culture. Fifty percent of inhibition was reached with only 0.0005 μg/mL of anti-IL-2 monoclonal MAB202 antibody while the inhibition level was obtained using 10 fold more anti-IL-2 polyclonal 202NA antibody.
Concerning IL-2 activity, a complete abolition of the IL-2 proliferative effect by anti-IL-2 antibodies was seen at concentrations above 0.05 μg/mL (Fig. 1b). In addition, incubation with anti-IL-2 antibodies alone did not alter cell viability, as demonstrated by the Trypan blue exclusion assay.
Anti-CD19 monoclonal antibody and anti-albumin polyclonal antibody were respectively used as controls showing no effects on CTLL2 proliferation in culture.
3.2 Inhibitory effect of IL-2 on HIV-infection in vitro
As shown in Fig. 2, pre-incubation of MT4 cells with both IL-2 and HIV-1 resulted in a strong inhibition of HIV-1 cell infection in vitro. After incubation, IL-2 and HIV-1 were removed by extensive washing, so HIV-1 p24 productive infection de novo was inhibited. A 70% reduction of HIV-1 p24 release was observed at 10 and 13 days of culture. The control cultures were negative for p24 release. MT4 cell proliferation was very significant in culture with IL-2 alone.
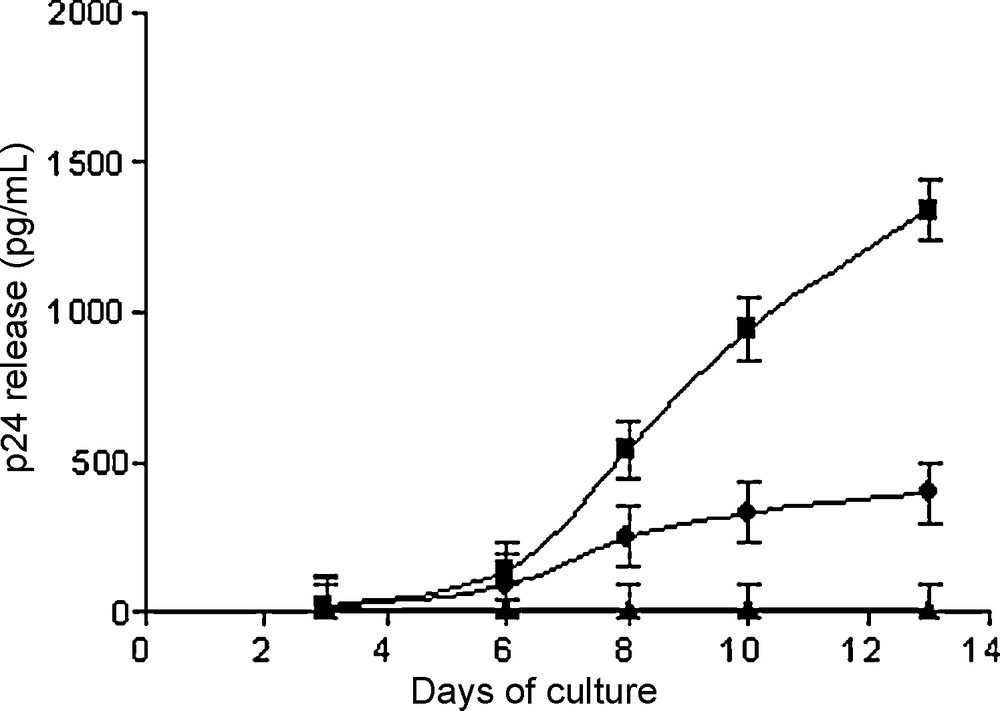
Effect of IL-2 on HIV-1 infection on MT4 cells in vitro. MT4 cells pre-incubated with IL-2 and HIV-1 Lai (), HIV-1 Lai alone (), and in the absence of both ().
In the Fig. 2, each individual value in this figure is the mean of results from three experiments (mean ± SE) and the differences were statistically significant (p < 0.01) at 8, 10 and 13 days.
In the presence of IL-2, a cytopathic effect was noted in HIV-infected MT4 cultures at only 10 days, whereas this effect occurred earlier, at day 6, in control cultures performed in the absence of IL-2.
3.3 Functional competition between gp41 peptides and IL-2
At concentrations of 0.005, 0.05, 0.5, 5 and 50 μg/mL (Fig. 3a), the trimeric gp41 molecule (loop) induced respectively 60, 70, 80, 85 and 90% of inhibition of CTLL2 cell proliferation. Comparative effects were observed with peptide #1 (residues 578–590) of HIV-1 gp41 showing respectively 40, 60, 65, 80 and 90% of proliferation inhibition. This dramatic inhibition of cell proliferation was not due to the peptide toxicity, since cell viability always remained above 90%.
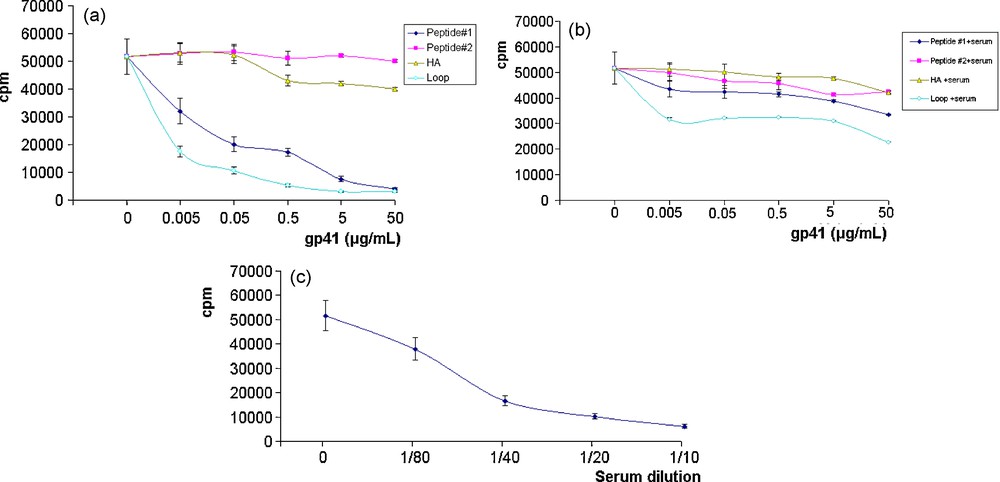
(a): effect of various gp41 peptides on CTLL2 cell proliferation; (b): effect of HIV-1 patients’ serum containing anti-gp41 antibodies on CTLL2 on cell proliferation in the presence of various gp41 peptides; (c): effect of HIV-1 patients’ serum containing anti-gp41 antibodies on CTLL2 on cell proliferation.
The primary structure of HIV-1 gp41 molecule (HA) provoked 18, 20, 22% inhibition in culture, at respectively 0.5, 5 and 50 μg/mL of HA gp41 molecule while peptide #2 (residues 587–599) of HIV-1 gp41 had no effect on CTLL2 proliferation (Fig. 3a).
In contrast, when the above cultures were carried out in the presence of pooled sera from HIV-1 positive patients (sera containing anti-gp41 antibodies), the proliferation of cells in culture was restored at a 50% level, as compared to proliferative control culture containing trimeric gp41 molecule (loop), at concentrations of 0.005, 0.05, 0.5 and 5 μg/mL (Fig. 3b). At 50 μg/mL of gp41 loop, cell proliferation in culture was restored at the level of 42%. This restoration reached 80% at 0.005, 0.05, 0.5 and 5 μg/mL and 70% at 50 μg/mL (peptide # 1).
As a test for reactivity between anti-gp41 antibodies with IL-2, pooled sera (1/10 to 1/80 dilutions) obtained from HIV-1 patients were mixed with IL-2 (10 ng/mL). This addition of anti-gp41 antibodies resulted in a dramatic inhibition of CTLL2 poliferation as shown in Fig. 3c. No effect was observed with control sera (data not shown).
The presented data are the mean ± SE of results from three experiments.
4 Discussion
A similarity between parts of IL-2 and gp41 molecules have previously been suggested [7,8], and we presently show this mimicry has functional consequences. By preventing the binding of gp41 to IL-2 receptor, IL-2 partially inhibited HIV in vitro infection with HIV-1 Lai. This finding did not exclude persistence of HIV infection in cells not expressing IL-2 receptor. Using another CD4+ cell line, the CEM cells (cell line expressing predominantly alpha and gamma but weakly beta chains of IL-2 receptor), we also observed comparable results in a 2-week experiment (data not shown). Further investigations will analyse the degree of inhibition in two other conditions, namely in peripheral blood mononuclear cells on the one hand, and using HIV primary isolates on the other hand. No reduction of cell viability and no cytopathogenic effect were observed, in comparison with controls, suggesting the lack of significant alteration of major cell signaling processes involving IL-2.
The functional relationship between IL-2 and HIV-1 may suggest either blocking of infection by IL-2 binding to receptor, or down-regulation of viral gene expression. Since the effect of the gp41 peptide #1 is likely to be the blockage of the IL-2 receptor of CTLL2 cells and strongly inhibited IL-2-induced proliferation of these cells, it is likely that the former mechanism explains the inhibitions.
The specificity in the inhibition of interaction between IL-2 and gp41 molecule, especially by trimeric gp41 molecule, was demonstrated by cross-reaction between anti-gp41 antibodies contained in HIV-1 patients’ sera and IL-2 in CTLL2 cultures. This cross-reaction resulted from a molecular mimicry between various regions of gp41 and IL-2 molecules. The gp41 molecule especially in its trimeric form might be suggested to require evaluation as a potential new candidate vaccine against HIV-1. However, detrimental effects on the IL-2 physiological activity might occur and would need to be ruled out before any investigation in humans.
The significance of our findings may shed some light on the relationships between IL-2 and HIV infection that are already known. The immune deficiency of AIDS patients results not only from the destruction of such cells as CD4+ lymphocytes and macrophages, but also from alteration of the cytokine network [10]. The decreased level of IL-2 plays a major role in the course of HIV infection. At comparable phases of evolution, IL-2 and interferons are diminished, while IL-10, IL-4 and tumor necrosis factor are increased [6,10]. IL-2 is known to exert amplifying activities on cytotoxic T lymphocytes, on NK cells, and to enhance lymphoproliferative responses [5,11,12]. IL-2 is an essential lymphokine for most functions of cell-mediated immunity and its use in the treatment of some AIDS patients has been demonstrated to be beneficial, from both the clinical and the immunological standpoints [13].
Acknowledgements
We are grateful to Dr. P-F. Serres for initial help to the project and Dr. R. Sembeil for data collection and presentation. We also thank Dr. T. Rousselle for the engineering of trimeric gp41 and Prof. L. Montagnier for numerous advicies and encouragement.