

1 Introduction
The specific identification and systematics of triatomines have been based fundamentally on morphological observations, mainly by means of optical [1] and electron scanning microscopy [2,3]. These organisms are classified into complexes and specific subcomplexes [4].
Triatoma lenti and T. sherlocki are hemipterans that belong to the Brasiliensis subcomplex [4]. This subcomplex is present in South America and consists of nine species, i.e., T. brasiliensis, T. juazeirensis, T. melanica, T. melanocephala, T. petrochiae, T. lenti, T. sherlocki, T. tibiamaculata and T. vitticeps [4]. However, the parameters used to group the subcomplex in triatomines were principally morphological and geographical disposition.
Costa et al. proposed the T. brasiliensis complex using egg morphology [3], morphological [3], biological [5], isoenzymes [6], and ecological [5] data. This complex comprises the subspecies T. b. brasiliensis and T. b. macromelanosoma, and species T. juazeirensis and T. melanica. Mendonça et al. (2009), by means of a phylogenetic reconstruction, proposed the recent inclusion of the T. sherlocki to this complex [7]. Thus, the complex T. brasiliensis shows up as a monophyletic group. It is possible that T. brasiliensis species complex may encompass other species such T. lenti. However, new approaches, such as molecular analysis, should be performed such as in T. sherlocki, to propose the grouping of this species in the complex [7,8].
T. lenti is an endemic triatomine of the States of Bahia, Goias, and Roraima [9]. The species is considered a potential vector for Chagas disease of secondary importance and it was found infected with the etiologic agent, the protozoan Trypanosoma cruzi [10]. T. sherlocki is an endemic species of Bahia [9]. It presents unique morphological characteristics that resemble those of T. lenti, as reduced hemelytra, reddish orange colored rings on the femur, and spots in the connective [11]. It was also reported that these organisms live in sympatry in some regions [12]. T. sherlocki has been found in human habitations, infected with T. cruzi, indicating a process of domiciliation of this vector [13]. Although both species were found infected with T. cruzi, only T. sherlocki presents epidemiological importance, because T. lenti is an exclusively sylvatic species [14].
The use of cytogenetic analyzes has been represented as a tool in systematic and taxonomy of cryptic species of triatomine [15,16]. In addition, it has been shown recently that cytogenetic is very important in the study of subcomplexes, since karyotype analysis was used to propose the exclusion of T. melanocephala, T. vitticeps, and T. tibiamaculata from the Brasiliensis subcomplex for approaching the triatomines in South America [17].
Thus, the present work, through the analysis of spermiogenesis, aims to characterize this stage of spermatogenesis in triatomines little studied, and especially to compare it among the species T. lenti and T. sherlocki, to assist in the diagnosis of differentiation of these insects.
2 Material and methods
In this study, five males of each species was used, assigned by the ‘Triatominae Insectarium’ installed at the Department of Biological Sciences, Faculty of Pharmaceutical Sciences, Araraquara campus. The seminiferous tubules of adult males, after being torn and fixated on cover slip, underwent the cytogenetic technique of lacto-acetic orcein [18]. The biological material was analyzed by a Jenaval light microscope (Zeiss) coupled with a digital camera and an image analyzer Axio Vision LE 4.8 (Copyright© 2006–2009 Carl Zeiss Imaging Solutions Gmb H). The images were magnified by a factor of 1000.
3 Results
By using the cytogenetic technique of lacto-acetic orcein, it was observed the elongation of the spermatid until differentiation into spermatozoa (Fig. 1(I–VI) and Fig. 2(I–VI)). During spermiogenesis of T. lenti, it was possible to detect a peripheral heteropycnotic filament in all cells (arrows). Already in T. sherlocki, the early spermatids showed a heteropyknotic corpuscle (Fig. 2I, arrowhead) that took the cell periphery during cell elongation (Fig. 2II–VI, arrowheads). During cell elongation of T. sherlocki, it was possible to observe the flagellum (Fig. 2II–VI, arrows). Note that the cells of this species are smaller and more tapered.
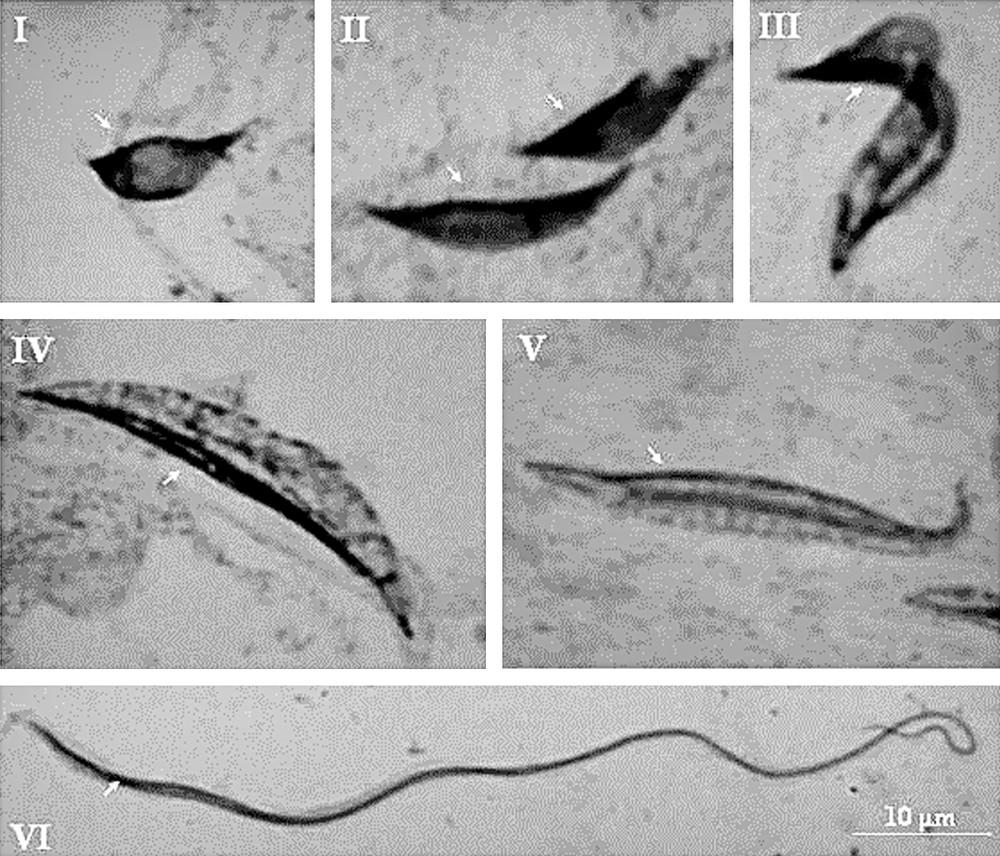
Spermiogenesis of Triatoma lenti (I–VI). Note a peripheral heteropcnotic filament in all cells (arrows).
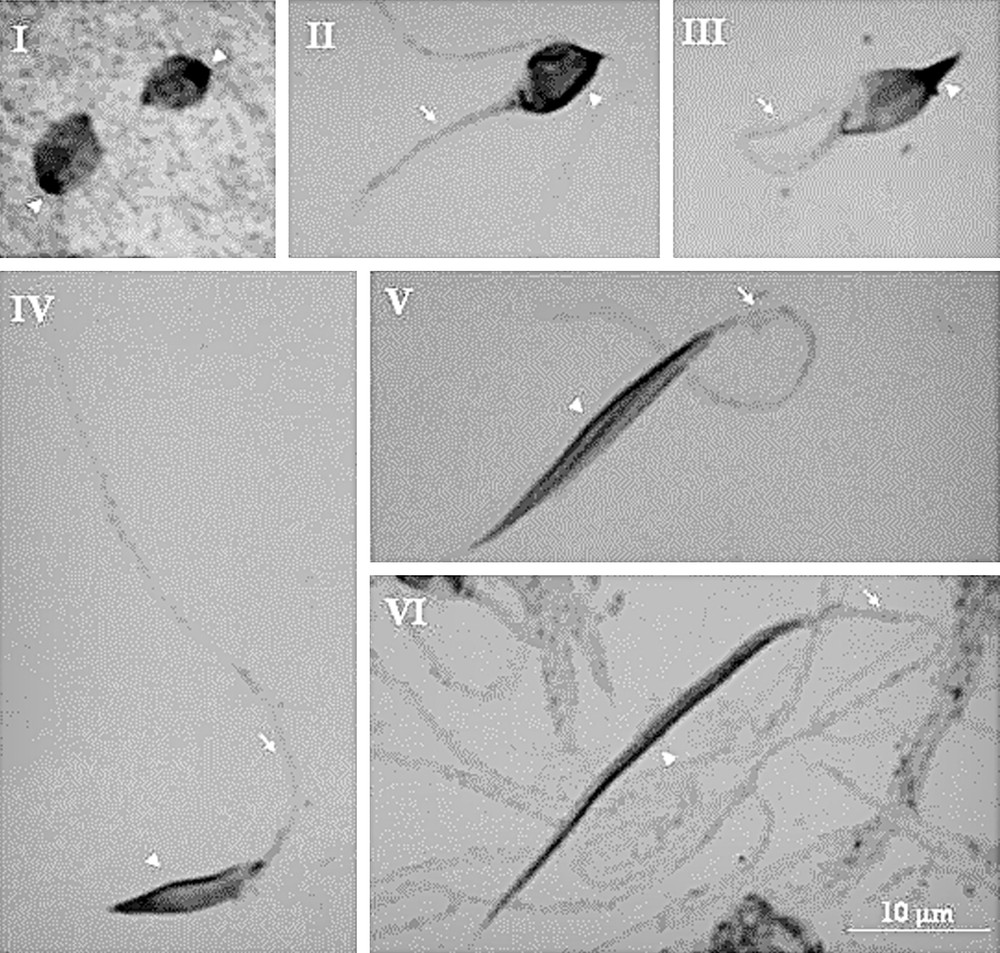
Spermiogenesis of Triatoma sherlocki (I–VI). I: note the heteropyknotic corpuscle (arrowheads); II–VI: note a peripheral heteropcnotic filament in all cells (arrowheads) and the flagellum (arrows).
4 Discussion
Spermatogenesis occurs in the seminiferous tubules and consists, respectively, of three major divisions, out more, spermatocitogenesis, meiosis and spermiogenesis, which results in the production of sperm [19]. Data on spermatogenesis in triatomines is commonly discussed in the literature, but focus mainly on meiosis, more specifically on the behavior of chromosomes.
By analyzing the spermiogenesis on T. lenti and T. sherlocki, we observed that both have a heteropyknotic filament on the periphery of the cells. This characteristic was also observed in T. pseudomaculata [20]. These organisms also share characteristics of egg morphology [21] and karyotypic, since all have 22 chromosomes [20,22,23].
The analysis of the initial spermatid of T. sherlocki allowed the visualization of a heteropyknotic corpuscle. This characteristic was also observed in T. klugi [24], T. maculata [20] and T. melanosoma [25]. T. maculata, T. sherlocki, and T. melanosoma also share characteristics of egg morphology [21]. Furthermore, all species have the diploid chromosome set 2n = 22A (20A + XY) [20,22,25].
The presence of the corpuscle is shown as a diagnostic tool to differentiate T. sherlocki and T. lenti, since it is absent in T. lenti. The analysis of the spermiogenesis in T. sherlocki also allowed us to address morphological differences between elongating cells, which were relatively smaller and more filamentous when compared to T lenti. Furthermore, the flagellum was observed in all stages of cell differentiation and elongation. This structure, which helps in the locomotion of the sperm, is hardly observed in cytogenetic analysis, especially throughout spermiogenesis.
Thus, although other comparative approaches should be taken, this paper allowed emphasize the analysis of spermiogenesis as an important cytotaxonomic tool that assists in the differentiation of morphologically related species, such as T. lenti and T. sherlocki.
Disclosure of interest
The authors declare that they have no conflicts of interest concerning this article.
Acknowledgments
This work was supported by the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and Coordenação de Aperfeiçoamento Pessoal de Nível Superior (CAPES).