

Abridged English version
Data collected in the context of the REPHY monitoring network constitute a significant basis for examining the large-scale temporal (1992–2000) and geographical variability (the whole French coasts) of microphytoplankton communities. Using these data, the objective was to identify the global temporal variability patterns of microphytoplankton populations observed along French coasts and to determine biogeographical areas based on the temporal variability of microphytoplankton populations. First, the temporal variability of microphytoplankton communities was studied at a large geographical scale. The methodological approach used for the analysis was based on descriptive multivariate techniques allowing multidimensional data to be summarized in a geometric space (Principal Component Analysis). The results allowed us to describe the temporal variability scheme, and particularly to illustrate the global seasonal trend for the most frequently observed species along French coasts. In a second step, a multitable comparison method (STATIS) was used to compare the temporal variability patterns of microphytoplankton communities observed at 17 sites along the coasts. Between-site differences were studied first, and a ‘common pattern’ (shared by all sites) was then identified. Finally, comparative analysis of the properties specific to each site indicated groups of sites with similar characteristics. Thus, three ‘homogeneous’ geographical areas exhibiting similar microphytoplankton population dynamics were distinguished: the western English Channel, the Bay of Biscay and the Mediterranean Sea. The microphytoplankton communities specific to each region were then identified. Ecological features showed the importance of hydrodynamic factors (freshwater inputs, tidal currents, weather conditions, etc.) in the geographical structure of microphytoplankton communities along French coasts. As REPHY sampling sites are located near the coast (average distance to the coast equal to 1.10 km), the identification of homogeneous geographical areas was not a predictable result. Indeed, in the context of REPHY sampling, local characteristics, as anthropogenic impacts, were supposed to dominate.
1 Introduction
Les communautés phytoplanctoniques sont constituées d'assemblages d'espèces co-occurrentes, présentant des caractéristiques biologiques (taille, forme...) et physiologiques (mode de nutrition, taux de croissance...) très différentes [1]. De nombreux facteurs environnementaux (hydroclimatiques et physicochimiques et biologiques) agissent sur la dynamique des populations phytoplanctoniques et, par conséquent, sur la structure des communautés phytoplanctoniques. Cette dynamique est fréquemment examinée à travers la réponse de la communauté dans son ensemble aux variations environnementales, à l'aide de variables synthétiques telles que la biomasse phytoplanctonique totale ou la teneur en pigments chlorophylliens [2]. Ces études sont essentielles pour comprendre le rôle des producteurs autotrophes dans le fonctionnement de l'écosystème pélagique. En revanche, elles ne permettent pas de répondre à d'autres problématiques, telles que la compréhension des mécanismes qui favorisent la dominance d'une ou de plusieurs population(s) phytoplanctonique(s) à l'intérieur de la communauté [1,3]. Ainsi, la connaissance des facteurs qui contrôlent la dynamique des communautés phytoplanctoniques dans leur ensemble peut s'avérer nécessaire à la compréhension des mécanismes favorisant les efflorescences phytoplanctoniques toxiques ou nuisibles.
L'objectif de cette étude est de décrire les principaux schémas de variabilité (temporelle et géographique) des communautés microphytoplanctoniques observées sur le littoral français. Cette étude à caractère exploratoire utilise les données du Réseau de surveillance du phytoplancton (REPHY) de l'Institut français de recherche pour l'exploitation de la mer (Ifremer). Dans un premier temps, la variabilité temporelle à grande échelle géographique est examinée. Une méthode statistique d'analyse conjointe de tableaux (STATIS) est ensuite utilisée afin d'identifier les spécificités géographiques des communautés microphytoplanctoniques sur le littoral français métropolitain.
2 Variabilité temporelle à grande échelle géographique
Le Réseau de surveillance du phytoplancton a été crée en 1984 par l'Ifremer pour répondre à deux objectifs complémentaires : le premier vise à documenter une base de connaissances en décrivant la distribution spatio-temporelle des différentes espèces phytoplanctoniques des eaux côtières et lagunaires de France métropolitaine. Le second objectif du REPHY répond à une préoccupation de protection de la santé publique ; il s'agit de détecter l'apparition d'espèces productrices de phycotoxines (toxines diarrhéiques paralysantes et amnésiantes), et de comparer les concentrations en phycotoxines accumulées dans la chair des coquillages à des normes réglementaires. Afin de répondre au premier objectif du REPHY, les abondances de l'ensemble des populations phytoplanctoniques observées sur le littoral de France métropolitaine sont dénombrées ; ces données sont disponibles depuis 1987 [4]. Vingt-neuf sites, répartis sur le littoral français, sont échantillonnés deux fois par mois. Les prélèvements et les identifications phytoplanctoniques sont assurés par les 12 laboratoires côtiers de l'Ifremer. Des problèmes dans la qualité des données (difficulté d'identification taxonomique, confusions entre certains genres et/ou espèces...) ont conduit (i) au regroupement de genres et d'espèces présentant des caractéristiques écologiques similaires et (ii) à la réduction de la fenêtre temporelle aux années 1992 à 2000. Un questionnaire présentant l'ensemble des regroupements contraints par des difficultés d'identification morphologique a été soumis à des experts du phytoplancton (cf. § Remerciements) afin de s'assurer de leur pertinence écologique. La diagnose des experts a permis de disposer de données inter-comparables dans le temps et entre sites d'échantillonnage [5].
Seuls les 17 sites régulièrement échantillonnés entre 1992 et 2000 (au moins une fois par mois) ont été conservés ; leur localisation est indiquée sur la Fig. 1. Ils sont décrits par les abondances mensuelles de 44 unités taxonomiques. Ces unités correspondent à des espèces, des groupes d'espèces, des genres ou des groupes de genres phytoplanctoniques (cf. Tableau 1).
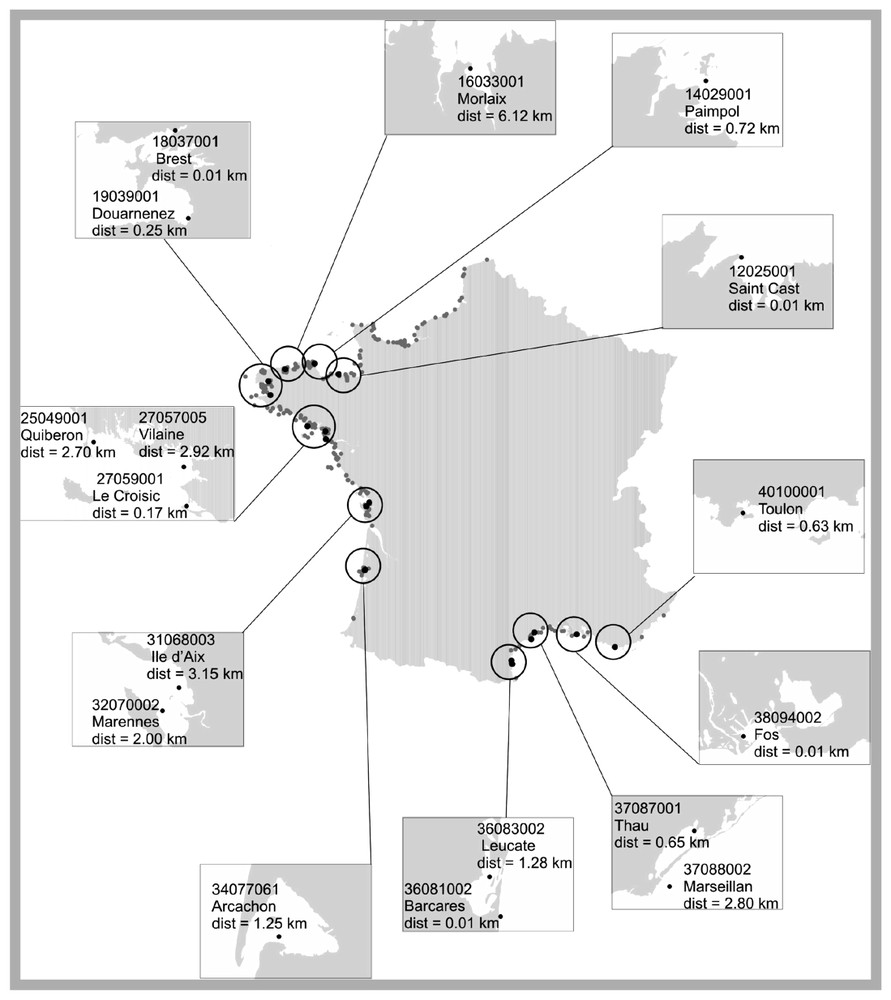
Localisation géographique des points d'échantillonnage du REPHY. Parmi ces sites, ceux sélectionnés pour l'analyse (17 sites) apparaissent en noir. Les numéros représentent les codes utilisés dans la base QUADRIGE pour décrire les différents sites. La profondeur de la colonne d'eau associée aux sites d'échantillonnage est comprise entre 0 et 10 m, la distance à la côte (« dist ») entre 0 et 3,15 km.
Liste des 44 unités taxonomiques sélectionnées pour l'analyse
| Diatomées | |
| ACHN | Achnanthes spp. |
| ASTE | Asterionella spp. (sauf A. glacialis + A. formosa) |
| ASTEGLA | Asterionella glacialis (= A. japonica) |
| BACIPAX | Bacillaria paxillifer (= B. paradoxa) |
| BACT | Bacteriastrum spp. |
| BELLITH | Bellerochea spp. + Lithodesmium spp. |
| BIDODON | Biddulphia spp. + Odontella spp. |
| CERAPEL | Cerataulina pelagica |
| CHAE | Chaetoceros spp. |
| DETLAUD | Lauderia spp. + Detonula spp. |
| DITYBRI | Ditylum brightwellii |
| EUCPZOD | Eucampia zodiacus |
| GRAM | Grammatophora spp. |
| GUINFLA | Guinardia flaccida |
| HEMI | Hemiaulus spp. |
| LEPT | Leptocylindrus spp. |
| LICM | Licmophora spp. |
| NAVIC | Amphora spp. + Diploneis spp. + Navicula spp. |
| NITZCYL | Nitzschia longissima + Cylindrotheca closterium |
| PORTHAL | Porosira spp. + Thalassiosira spp. |
| PRORHIZ | Proboscia spp. + Rhizosolenia spp. |
| PSNZ | Pseudo-nitzschia spp. |
| RHABSTRI | Rhabdonema spp. + Striatella spp. |
| SKELCOS | Skeletonema costatum |
| THAA | Thalassionema spp. + Thalassiothrix spp. |
| Dinoflagellés | |
| ALEX | Alexandrium spp. (sauf A. affine + A. andersoni + A. leei + A. hiranoi + A. pseudogonyaulax + A. insuetum + A. margalefi |
| + A. minutum + A. ostenfeldii + A. tamarense) | |
| ALEXMIN | Alexandrium minutum |
| CERI | Ceratium spp. |
| DINOAC | Complexe Dinophysis acuminata + Dinophysis sacculus |
| DINODI | Dinophysis caudate + D. diegensis |
| DINOROT | Dinophysis rotundata |
| EBRARTRI | Ebria tripartita |
| GONY | Gonyaulax spp. (sauf G. spinifera) |
| GONYSPI | Gonyaulax spinifera |
| GYMN-82 | Gymnodinium chlorophorum |
| GYMNNAG | Gymnodinium nagasakiense (= G. mikimotoi = Karenia mikimotoi) |
| GYMNO | Warnowia spp. + Nematodinium spp. + Amphidinium spp. + Cochlodinium spp. + Gyrodinium spp. + Katodinium spp. |
| + Gymnodinium spp. (sauf G. chlorophorum + G. breve + G. cf. breve + G. mikimotoi = Karenia mikimotoi) | |
| NOCTSCI | Noctiluca scintillans |
| PERID | Diplopsalis spp. + Diplopelta spp. + Diplopsalopsis spp. + Zygabikodinium spp. + Oblea spp. |
| PLESGYR | Pleurosigma spp. + Gyrosigma spp. |
| POLYSCH | Polykrykos schwartzii |
| PROR | Prorocentrum spp. (= Exuviaella sp.) |
| PRORLIME | Prorocentrum lima + P. marinum + P. mexicanum |
| CLEUGLE | Euglenophycées |
Soit Astu l'abondance (exprimée en nombre de cellules par litre) de l'unité taxonomique u (U=44) au temps t (un des mois de la période janvier 1992 à avril 2000 ; T=100) sur le site s (S=17). Afin de stabiliser les variances, les abondances sont log-transformées : Xstu=log10(Astu+1). Dans un premier temps, on s'intéresse à la description de la variabilité temporelle à long terme des populations microphytoplanctoniques. Afin de dégager la composante temporelle commune à l'ensemble des sites d'échantillonnage, la moyenne m.tu, ainsi que la variance var.tu, pour une date t, et sur l'ensemble des sites, sont estimées pour chaque unité taxonomique u. Deux tableaux des moyennes géographiques (M), ainsi que des variances associées (V), sont obtenus. Une analyse en composantes principales (ACP) du tableau M, de dimension T×U, est ensuite utilisée afin de décrire les principales structures observées entre individus (les dates) et les corrélations entre variables (unités taxonomiques). Environ 40 % de la variabilité totale (variance) sont expliqués par les deux premières composantes principales. L'examen de la Fig. 2, qui décrit la structure des individus dans le premier plan factoriel de l'ACP du tableau M, met en évidence une forte composante saisonnière. L'axe 1 oppose les mois hivernaux (coordonnées négatives) aux mois estivaux (coordonnées positives) ; l'axe 2 oppose les mois printaniers (coordonnées négatives) aux mois automnaux (coordonnées positives). La représentation des variables dans l'espace dual (figure non présentée) permet d'identifier le cycle saisonnier des populations phytoplanctoniques observées sur le littoral français. La Fig. 3 décrit, à titre d'exemple, le cycle saisonnier de 12 unités taxonomiques identifiées comme contribuant à l'explication de la variabilité temporelle par l'ACP du tableau M. Elle met en évidence le schéma de succession des communautés phytoplanctoniques dans les eaux côtières de France métropolitaine. La progression saisonnière de communautés dominées par des Diatomées en hiver et en début de printemps (Fig. 3A) vers des communautés dominées par des Dinoflagellés en été (Fig. 3B) mise en évidence dans cette étude est caractéristique du schéma de succession saisonnière des communautés phytoplanctoniques en domaine tempéré [6]. Ce schéma classique, décrit par de nombreuses études (par exemple, [7–9]), est associé à des propriétés physicochimiques de la colonne d'eau : le mélange vertical de la colonne d'eau favorise le développement des floraisons à Diatomées hivernales, printanières et automnales, les floraisons estivales de Dinoflagellés étant associées à des conditions de stratification de la colonne d'eau.
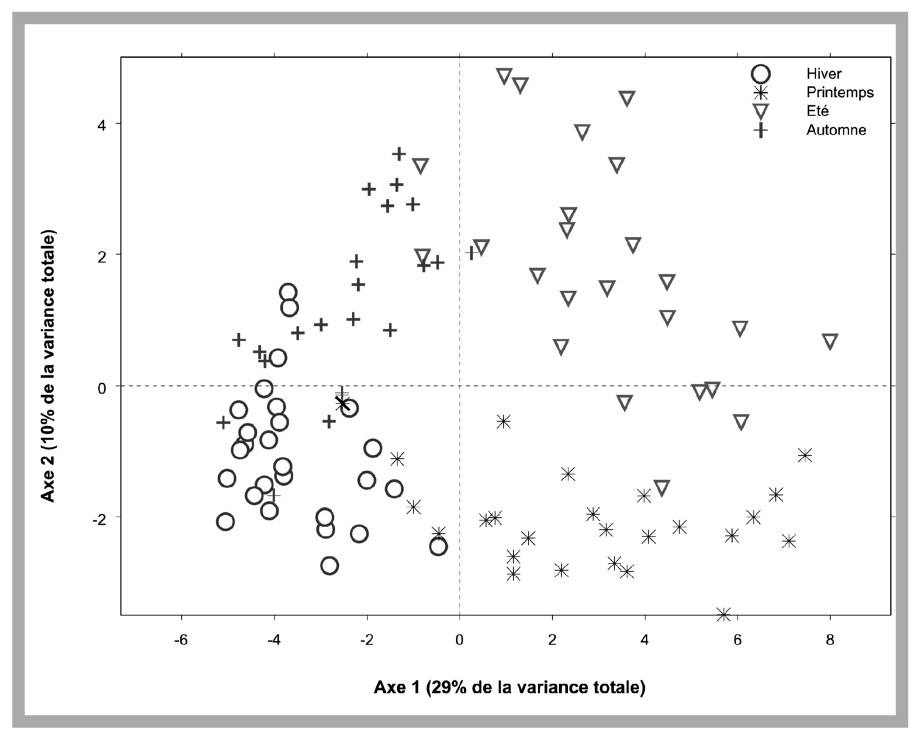
Représentation des individus (les dates) dans le premier plan factoriel de l'ACP du tableau M. Différents symboles sont utilisés pour distinguer les dates associées aux différentes saisons.
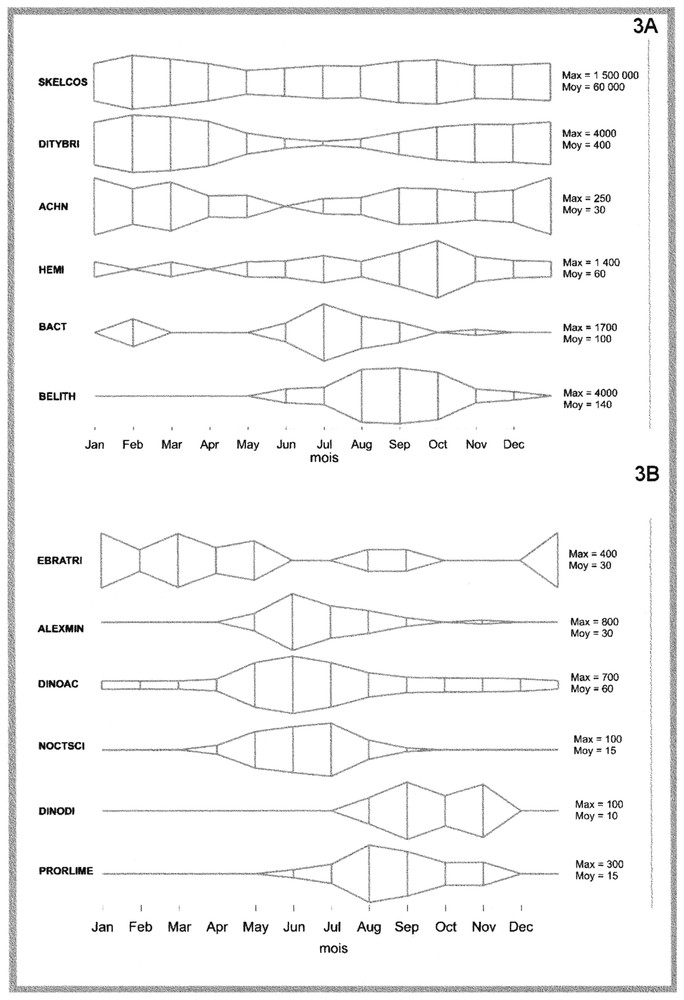
Cycle saisonnier de six Diatomées (Fig. 3A) et de six Dinoflagellés (Fig. 3B) identifiées comme contribuant à l'explication de la variabilité temporelle. Les graphiques représentent les variations de l'abondance mensuelle moyenne (pour les neuf années sélectionnées). L'échelle des abondances varie d'un taxon à l'autre, la concentration moyenne (« moy ») ainsi que le maximum d'abondance (« max ») sont indiqués pour chaque taxon (nombre de cellules par litre). La plupart des Diatomées atteignent leurs maxima d'abondance en hiver et à l'automne, à l'exception de l'espèce Skeletonema costatum (code SKELCOS) qui est présente toute l'année. Les Dinoflagellés présentent des maxima d'abondance en été (codes ALEXMIN, DINOAC et NOCTSCI), en fin d'été pour les espèces Dinophysis caudata + D. diegensis (code DINODI) et Prorocentrum lima + P. mexicanum + P. marinum (code PRORLIME). L'espèce Ebria tripartita (code EBRATRI) atteint deux maxima de concentrations, un au printemps et un en fin d'été. Masquer
Cycle saisonnier de six Diatomées (Fig. 3A) et de six Dinoflagellés (Fig. 3B) identifiées comme contribuant à l'explication de la variabilité temporelle. Les graphiques représentent les variations de l'abondance mensuelle moyenne (pour les neuf années sélectionnées). L'échelle des abondances varie ... Lire la suite
3 Variabilité géographique : comparaison inter-sites
L'objectif est désormais de comparer la variabilité temporelle des communautés phytoplanctoniques entre sites d'échantillonnage. L'analyse de la variabilité temporelle à grande échelle géographique a mis en évidence une importante composante saisonnière commune à l'ensemble des sites (40 % de variance expliquée). Afin de retirer cette composante commune et de ne conserver que les spécificités locales, les données sont, dans un premier temps, standardisées :
3.1 Méthode et résultats
La méthode STATIS [10–12] est utilisée ici. Cette méthode, dont on trouve des applications en océanologie (par exemple, [13,14]), permet d'analyser conjointement plusieurs tableaux de données. Elle a pour but d'analyser dans quelle mesure les distances entre les différentes unités taxonomiques restent comparables d'un site à l'autre. Elle procède en trois étapes :
- • l'analyse des différences globales entre les tableaux associés aux différents sites ; deux tableaux sont considérés comme d'autant plus différents que les distances entre unités taxonomiques, calculées à partir de chacun d'entre eux au moyen d'une ACP, sont différentes (à une constante multiplicative près) ;
- • la recherche d'un tableau « compromis » susceptible de résumer l'ensemble de tous les tableaux associés aux sites ; ce tableau est d'autant plus représentatif de l'ensemble des tableaux qu'ils sont peu différents entre eux ; les variations des positions relatives des unités taxonomiques d'un site à l'autre peuvent être analysées à l'aide de ce compromis ;
- • l'analyse des positions des unités taxonomiques dans tous les sites par rapport au tableau compromis.
Le détail de la méthode est présenté par Lavit [15] ainsi que par Dazy et Le Barzic [16].
3.1.1 Interstructure : différences entre sites
L'analyse des différences entre les tableaux sites, ou interstructure, nécessite la définition d'une distance. La distance choisie entre deux tableaux dans la méthode STATIS est obtenue à partir du coefficient de corrélation vectorielle d'Escoufier (1973) :
Une ACP du tableau RV des coefficients de corrélation vectorielle permet de visualiser l'interstructure (Fig. 4). Les deux premiers axes de l'ACP du tableau RV représentent respectivement 87 % et 2 % de la variance totale. Le tableau des coefficients RV est une matrice symétrique, dont les valeurs sont comprises par définition entre zéro et 1 ; ainsi, tous les éléments du premier vecteur propre sont positifs. L'examen de la Fig. 4 permet de distinguer trois zones géographiques : la Manche occidentale (Saint-Cast, Paimpol, Morlaix et Brest, coordonnée négative sur l'axe 2), le golfe de Gascogne (de Douarnenez à Arcachon, coordonnées positives sur l'axe 2) et la Méditerranée (Barcarès, Leucate, Thau, Marseillan, Fos et Toulon, coordonnées proches de zéro sur l'axe 2).
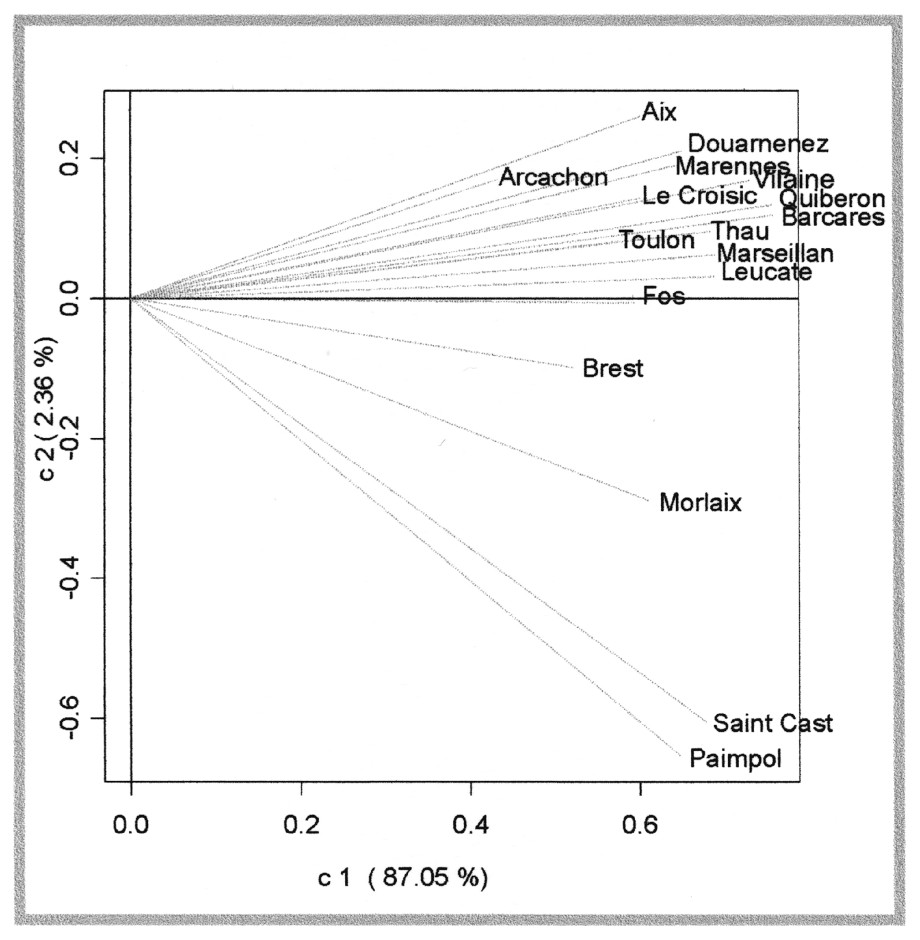
Analyse de l'interstructure, proximités entre tableaux sites (unités taxonomiques × dates). Les corrélations entre tableaux sites sont calculées en utilisant le coefficient RV. Trois zones géographiques sont identifiées : la Manche occidentale (Saint-Cast, Paimpol, Morlaix et Brest, coordonnées négatives sur l'axe 2), les sites de Méditerranée (Barcares, Toulon, Thau Marseillan, Leucate et Fos, coordonnées proches de zéro sur l'axe 2) et les sites du golfe de Gascogne (de Douarnenez à Arcachon).
3.1.2 Compromis : description d'un site « moyen »
Dans la mesure où les tableaux se ressemblent, il est particulièrement utile de mettre en évidence les caractéristiques essentielles de cette ressemblance. Dans la méthode STATIS, le tableau « compromis » est la combinaison linéaire de tous les tableaux qui maximise la somme de ses coefficients RV avec les tableaux initiaux. Le tableau compromis présente donc une « corrélation maximum » avec l'ensemble des tableaux associés aux sites. Il est obtenu en calculant une somme pondérée des tableaux
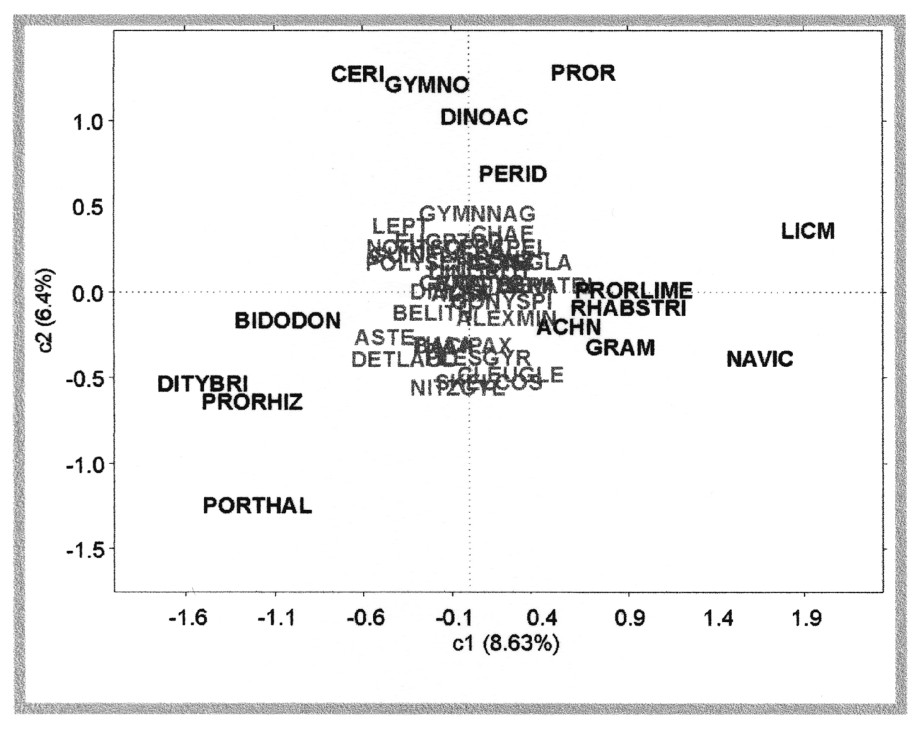
Image euclidienne compromis des individus dans le plan 1–2 (15% de la variance totale sont expliqués). Les unités taxonomiques indiquées en grisé contribuent peu à la construction du compromis. L'axe 1 oppose un groupe d'unités taxonomiques appartenant à la classe des Diatomées (PORTHAL, DITYBRI, PRORHIZ et BIDDODON) à un autre groupe également majoritairement constitué de Diatomées (LICM, NAVIC, GRAM et RHABSTRI), à l'exception d'un groupe d'espèce appartenant à la classe des Dinoflagellés (PRORLIME). L'axe 2 permet d'isoler un groupe constitué exclusivement par des Dinoflagellés (CERI, GYMNO, DINOAC, PERID et PROR, coordonnées positives). Masquer
Image euclidienne compromis des individus dans le plan 1–2 (15% de la variance totale sont expliqués). Les unités taxonomiques indiquées en grisé contribuent peu à la construction du compromis. L'axe 1 oppose un groupe d'unités taxonomiques appartenant à la classe des ... Lire la suite
3.1.3 Intrastructure : comparaison tableaux sites/compromis
Le calcul des corrélations des variables de chaque tableau (les 100 dates) avec les axes du compromis permet d'avoir une représentation de la relation existant entre les tableaux sites et le tableau compromis. Ces corrélations peuvent être résumées sur un graphique dont l'étude permet d'interpréter les positions compromis des individus. Les corrélations des 100 variables (dates) de chaque tableau site (17 tableaux) avec les axes du compromis sont calculées. Pour des raisons de lisibilité, un découpage par mois a été effectué. L'examen de la Fig. 6, qui illustre à titre d'exemple la structure des variables pour le mois de mai dans le plan 1–2 du compromis, permet ainsi d'interpréter plus facilement la position des individus dans « l'espace compromis » (Fig. 5). L'axe 1 oppose les sites de Manche occidentale et du golfe de Gascogne (coordonnées négatives ou proches de 0) aux sites méditerranéens (coordonnées positives) ; l'axe 2 oppose les sites de Manche occidentale (coordonnées négatives) aux sites du golfe de Gascogne et de Méditerranée (coordonnées positives ou proches de zéro) (Fig. 6). Ainsi, les unités taxonomiques présentant des coordonnées positives sur l'axe 1 du compromis (par exemple, RHABSTRI, GRAM, NAVIC et LICM) sont associées aux sites méditerranéens, tandis que les unités taxonomiques présentant des coordonnées négatives sur l'axe 1 et positives ou proches de zéro sur l'axe 2 (par exemple, CERI, GYMNO et DINOAC) sont associées aux sites du golfe de Gascogne, les unités taxonomiques présentant des coordonnées négatives sur les axes 1 et 2 (par exemple, PORTHAL, PRORHIZ et DITYBRI) étant associées aux sites de Manche occidentale.

Exemple de corrélations des variables (dates) de l'ensemble des 17 sites avec les deux premières composantes principales du compromis, pour le mois de mai. Sur chaque graphique, les différentes années (neuf années et 17 sites) sont représentées, pour des raisons de lisibilité (et dans l'absence d'une structure inter-annuelle, graphiques non présentés), les dates des différents sites ne sont pas indiquées. Les symboles permettent de différencier les dates des sites de Manche (losanges), du golfe de Gascogne (triangles) et de Méditerranée (carrés). Le plan 1–2 isole les sites de Méditerranée (coordonnées positives ou proches de zéro sur les deux axes) des sites de la Manche (coordonnées négatives ou proches de zéro sur l'axe 1 et négatives sur l'axe 2) et du golfe de Gascogne (coordonnées négatives ou proches de zéro sur l'axe 1 et positives ou proches de zéro sur l'axe 2). Masquer
Exemple de corrélations des variables (dates) de l'ensemble des 17 sites avec les deux premières composantes principales du compromis, pour le mois de mai. Sur chaque graphique, les différentes années (neuf années et 17 sites) sont représentées, pour des raisons ... Lire la suite
3.2 Discussion
Les analyses de l'interstructure (Fig. 4), du compromis (Fig. 5) et de l'infrastructure (Fig. 6), au moyen d'analyses en composantes principales et de représentations graphiques des variables, permettent d'identifier une typologie géographique des communautés microphytoplanctoniques observées le long des côtes françaises. Les unités taxonomiques associées aux sites de Manche occidentale (coordonnées négatives sur les axes 1 et 2 du compromis, Fig. 5) appartiennent à la classe des Diatomées, tandis que le golfe de Gascogne est caractérisé par des unités taxonomiques appartenant à la classe des Dinoflagellés (coordonnées positives sur l'axe 2 du compromis, Fig. 5) ; enfin, les unités taxonomiques présentant des coordonnées positives sur l'axe 1 du compromis, qui isole les sites de la Méditerranée, appartiennent majoritairement à la classe des Diatomées. L'identification d'une typologie des sites en région n'était pas un résultat acquis a priori. En effet, la localisation des sites d'échantillonnage du REPHY est conditionnée par la vocation de protection de la santé publique du réseau, c'est-à-dire à proximité de zones de production conchylicole, en domaine côtier ou lagunaire (distance moyenne à la côte de 1,1 km). Ces zones sont soumises à d'importants apports continentaux et présentent des spécificités biologiques et physicochimiques propres. Ainsi, l'influence de facteurs locaux (comme, par exemple, les apports plus ou moins importants en nutriments) aurait pu exercer une influence prépondérante sur la structure des communautés microphytoplanctoniques. La reconnaissance de zones géographiques au sein desquelles les communautés microphytoplanctoniques présentent des schémas de variabilité temporelle semblables montre que la composition des communautés semble associée aux caractéristiques hydrodynamiques des différentes régions. En effet, de nombreuses études ont montré qu'il existait une association marquée entre la structure des communautés phytoplanctoniques, et plus particulièrement entre la dominance Diatomées/Dinoflagellés, et les propriétés physiques de la colonne d'eau (par exemple, [17–19]). La turbulence inhibe la croissance des Dinoflagellés en causant des dommages physiques, des altérations physiologiques et des modifications comportementales, alors que les Diatomées sont beaucoup moins sensibles à la turbulence [1,3,20]. Or, les trois régions identifiées présentent des caractéristiques hydroclimatiques (apports en eau douce, courants de marée, stratification de la colonne d'eau...) différentes.
Les côtes françaises de Manche occidentale sont soumises à de faibles apports d'eau douce. Du point de vue hydrodynamique, cette région est caractérisée par de forts courants de marée, qui ont une influence majeure sur les propriétés physiques de la colonne d'eau. En effet, les forçages physiques induits par les courants de marée entraı̂nent un brassage vertical permanent empêchant toute stratification de la colonne d'eau tout au long de l'année [21]. L'ensemble de ces caractéristiques crée des conditions favorables à la croissance de Diatomées, telles que celles qui ont été identifiées dans notre étude. Ce résultat est confirmé par d'autres études portant sur la composition des communautés phytoplanctoniques de Manche occidentale, qui mettent également en évidence la dominance des Diatomées dans cette région (par exemple, [22–24]). Parmi les populations phytoplanctoniques identifiées, comme caractéristiques de Manche occidentale dans notre étude, les genres Ditylum (code DITYBRI) et Thalassiosira (code PORTHAL) sont décrits comme caractéristiques des floraisons hivernales et printanières des côtes françaises de Manche occidentale, et le genre Rhizosolenia (code PRORHIZ) comme dominant dans les communautés phytoplanctoniques estivales [25–27].
Contrairement aux côtes de Manche occidentale, les côtes françaises du golfe de Gascogne sont soumises à d'importants apports d'eau douce (principalement en provenance de la Loire, de la Gironde et de l'Adour). Les courants de marée sont plus faibles, et la circulation hydrodynamique est principalement contrôlée par le vent. Dans cette région, en été, les vents de nord-ouest induisent des upwellings qui favorisent la dispersion des eaux dessalées le long du plateau continental. Ce phénomène, ainsi que la formation d'une thermocline, provoquent une stratification de la colonne d'eau [28]. La stratification estivale est favorable au développement des Dinoflagellés, classe à laquelle appartiennent les unités taxonomiques identifiées comme caractéristiques de cette zone dans notre étude. Parmi les populations phytoplanctoniques identifiées par l'analyse, les Dinoflagellés Ceratium (code CERI) et Protoperidinium (code PERID) sont décrits comme dominants dans les communautés phytoplanctoniques estivales du golfe de Gascogne sud [29]. Des travaux récents à l'échelle des côtes françaises du golfe de Gascogne [30] identifient la classe des Dinoflagellés (et notamment, le genre Gymnodinium) comme dominants dans la partie sud.
La Méditerranée est un bassin d'évaporation semi-fermé, caractérisé par d'importants échanges avec l'Atlantique au niveau du détroit de Gibraltar. Le climat (et particulièrement l'irradiance) exerce une influence majeure sur les propriétés physiques du bassin méditerranéen (température et salinité élevées, et système oligotrophe) [31]. Les caractéristiques hydrodynamiques de la Méditerranée sont essentiellement marquées par de très faibles amplitudes de marée et par l'importance du rôle de la pression atmosphérique et du vent. Dans cette région, la période de stratification de la colonne d'eau, induite par l'établissement d'une thermocline, s'étend de mai à octobre [32]. Ce phénomène favorise le développement de Dinoflagellés caractéristiques d'eau tempérées à chaudes, tels que les espèces Prorocentrum lima + P. marinum + P. mexicanum (associées aux sites méditerranéens dans notre étude, coordonnées positives sur l'axe 1 du compromis, cf. Fig. 5). Les lagunes méditerranéennes (telles que les lagunes de Leucate et de Thau, dont les données sont utilisées ici) constituent des écosystèmes particuliers, les échanges avec la mer et la circulation sont contrôlés par le vent et la pression atmosphérique. Leur faible profondeur entraı̂ne un brassage permanent de la colonne d'eau [33]. Ces lagunes sont décrites comme « écosystèmes à Diatomées » [34]. En effet, le brassage permanent de la colonne d'eau favorise la croissance des Diatomées, et permet notamment la remise en suspension dans la colonne d'eau de genres benthiques à tychopélagiques, tels que les genres Achnanthes, Rhabdonema, Striatella, Grammatophora, Licmophora ou Navicula, identifiés dans notre étude.
4 Conclusion
L'analyse de la variabilité temporelle à grande échelle géographique a permis d'identifier la composante temporelle commune à l'ensemble des sites, et de décrire le cycle saisonnier des populations microphytoplanctoniques observées sur le littoral français. L'analyse de la variabilité géographique a mis en évidence une typologie géographique des communautés microphytoplanctoniques observées sur le littoral de France métropolitaine. Trois zones géographiques homogènes ont été distinguées : la Manche occidentale, le golfe de Gascogne et la Méditerranée. Les espèces responsables de la structure géographique ont ensuite été identifiées. La connaissance de l'écologie des populations microphytoplanctoniques associées aux différentes régions confirme l'importance du rôle des facteurs hydrodynamiques sur la distribution géographique des communautés microphytoplanctoniques observées sur le littoral français. Malgré la localisation côtière des points d'échantillonnage du REPHY, qui sont soumis à d'importants apports anthropiques, les propriétés hydrodynamiques générales des différentes régions (conditionnées par les apports fluviaux, les courants de marée, le vent...) semblent jouer un rôle prépondérant. Ce résultat constitue une base pertinente pour une analyse approfondie de l'influence des facteurs environnementaux sur la structure des communautés phytoplanctoniques.
Remerciements
Les auteurs remercient C. Billard, J.D. Dodge, V. Martin-Jézéquel, E. Nézan, G. Paulmier, Y. Rincé et D. Vaulot, ainsi que les 12 laboratoires côtiers d'Ifremer. Cette étude s'inscrit dans le cadre du Programme national d'environnement côtier (PNEC).