

Abridged English version
Most of the parental behaviors that increase fitness of the offspring have been demonstrated in the presence of the parents. A parental contribution that effects their young's survival can exist even after the death of the parents. The mother of the subsocial spider, Amaurobius ferox (Araneae, Amaurobiidae), is sacrificed for its young who actively cannibalize her. After matriphagy, the siblings do not leave the maternal nest and live in a group until 3–4 weeks later, when they disperse, in contrast to other subsocial spiders leaving the natal nest as soon as the mother disappears. During this post-maternal period, the siblings develop capturing ability and show collective capturing activity against large prey.
Why do the young of A. ferox remain in the natal nest when the mother is no longer present? I investigated the benefits that the young obtain from the maternal web in their collective prey capture. During the maternal period, females of A. ferox build their largest webs, using adhesive and elastic cribellate silk, the production of which is very expensive compared to ecribellate silk.
I compared collective capturing behaviors of the young against a prey 10 times larger than a single spiderling. The experiment had two treatments: presentation of the prey within the maternal web ( clutches) and presentation within a web built by the siblings ( clutches). From video-recorded capturing sequences, I observed the proportion of the clutches that paralyzed the prey, counted the number of participants, and analyzed successive attacks (contacts) and seizures against the prey. I also compared the treatments' effects on agonistic interactions between siblings.
The adaptive value of the maternal web was evident in the results. Prey-capturing activity was more effective when the young remained on the maternal web than when they used the web constructed by themselves (70% vs. 28%). Maternal webs may also provide better transmission of information on the prey's state and siblings' activities, because more spiderlings were recruited to contact and seize the prey in a shorter time. Time taken by the first individual within the clutch to attack and seize the prey did not differ between the two treatments. However, the time intervals between the first and second attacks and between the first and second seizures were shorter on the maternal web. Moreover, the second, third and fourth attacks and seizures occurred more rapidly on the maternal web. Agonistic interactions between the siblings were rarely observed in either treatment.
Amaurobius ferox has traits characteristic of an intermediate evolutionary stage between the maternal social stage and permanently social stage in spiders. Hence the species provides an opportunity to understand how sociality might evolve via a subsocial pathway. The young's inheritance of the maternal nest is probably one of the crucial ways in which the mother influences the post-maternal social period of the young. With her web, the mother shares in the cost of web construction, protects her young against predators, and assists in prey capture well after her death.
1 Introduction
L'aspect adaptatif des soins parentaux est de plus en plus perçu [1–4]. Ces comportements parentaux, qui augmentent le fitness des jeunes, sont mis en évidence en présence de leurs parents. Une contribution des parents, même après leur mort, peut toujours exister, en intervenant sur la survie des jeunes (par exemple, [5]).
Chez Amaurobius ferox (Araneae, Amaurobiidae), la mère se sacrifie pour ses jeunes, qui cannibalisent activement leur mère environ une semaine après l'émergence du cocon [6]. Après avoir dévoré la mère (matriphagie), les jeunes d'A. ferox ne quittent pas le nid maternel. Ils vivent en groupe pendant encore trois à quatre semaines avant la dispersion [7], contrairement aux autres araignées maternelles étudiées, pour lesquelles les jeunes quittent le nid dès que la mère disparaît [8–10]. Environ une semaine après la matriphagie, les jeunes effectuent leur seconde mue et présentent une activité prédatrice vis-à-vis d'une proie quelques jours plus tard. Cependant la capacité à filer la soie cribellée commence à s'observer entre la première et la deuxième mue, c'est-à-dire au second stade [11].
Pourquoi les jeunes d'Amaurobius ne quittent-ils pas le nid natal après la disparition de la mère ? On peut s'interroger sur le bénéfice qu'ils sont susceptibles de retirer de la présence d'une toile qu'ils n'ont pas eu à construire. La mère d'A. ferox tisse sa plus grande toile avant la période d'incubation, et pendant la période maternelle (construction du cocon – matriphagie) elle continue de tisser, renforçant l'intérieur de la toile et réparant les dégâts. Cette toile représente un héritage maternel qui pourrait être un facteur important dans l'évolution de la vie sociale chez les araignées [5,12] : une extension graduelle de la vie grégaire des jeunes, renforcée par les bénéfices de la toile maternelle.
Cet éventuel bénéfice de la toile maternelle peut être recherché au niveau de la protection contre des prédateurs potentiels ou de la capture des grandes proies. La toile d'Amaurobius est constituée d'un réseau complexe de fils cribellés, à l'intérieur duquel existe une retraite en forme de tube où les jeunes se regroupent. La soie cribellée, qui a un coût de fabrication bien supérieur à celui de la soie non cribellée [13], rend la toile d'Amaurobius remarquablement adhésive et élastique [14,15]. Ces caractéristiques persistent tant que la toile n'est pas abîmée par la pluie ou la poussière [13,16], et rendent l'approche de proies ou de prédateurs difficile, au risque de se faire piéger par la toile. Le bénéfice protecteur de la toile construite par la mère semble évident pour les jeunes. D'ailleurs, des observations de laboratoire et de terrain ont montré l'efficacité de cette toile vis-à-vis d'insectes et d'araignées écribellates sympatriques, les empêchant d'atteindre la retraite [11]. En revanche, des congénères et des individus d'une autre espèce d'araignée cribellate (Amaurobius fenestralis) ont réussi à entrer dans la retraite sans être piégés par la soie, et se sont révélés être de dangereux prédateurs pour les jeunes [11].
Par ailleurs, les jeunes d'Amaurobius pourraient adopter à cet âge, c'est-à-dire après la mort de la mère, deux stratégies de vie différentes. La première consisterait à quitter le nid maternel dans lequel il n'y a plus de mère, tandis la seconde serait d'y rester encore avec les frères et sœurs. Cependant, dans la nature et au laboratoire, si on détruit la toile maternelle, les jeunes se regroupent ailleurs et construisent une toile commune. Il est bien possible que, dans la nature, il y ait des prédateurs ou des facteurs abiotiques qui pourraient détruire le nid maternel et pousser les jeunes d'Amaurobius à le quitter.
Cette étude s'intéresse au bénéfice que les jeunes d'A. ferox peuvent retirer de la toile maternelle pour la prédation collective au cours de la période sociale post-maternelle, qui va de la mort de la mère à la dispersion des jeunes. Pendant cette période, les jeunes développent leur activité de capture, à savoir essentiellement une activité collective vis-à-vis d'une proie beaucoup plus grande qu'eux. Cette étude compare la capture d'une proie par des jeunes vivant dans la toile maternelle à la même action dans une toile construite par les jeunes eux-mêmes. L'efficacité de la capture, l'enchaînement des actes des jeunes vis-à-vis de la proie et les manifestations agonistiques entre les jeunes sont analysés.
2 Matériel et méthodes
Amaurobius ferox Walckenaer (Araneidae, Amaurobiidae) est répandue en Europe [17,18]. Elle vit dans des endroits sombres et humides, à l'abri du soleil : principalement de vieux murs, sous des pierres, dans de vieilles caves ou des tunnels [17,18].
La mère d'Amaurobius commence à tisser son cocon au début de l'été, et reste à proximité pendant les trois semaines d'incubation. À l'émergence, les jeunes dépendent de leur mère pour leur alimentation : consommation d'une ponte trophique un jour après l'émergence, puis consommation de la mère elle-même quelques jours plus tard. Après la matriphagie, les jeunes mènent une vie communautaire au sein du nid maternel pendant 24 jours en moyenne, puis ils se dispersent [19].
Des femelles fécondées, repérées grâce à la présence d'un bouchon vaginal sous la plaque génitale [20], ont été récoltées en mai–juin (c'est-à-dire au début du cycle parental) sur des parcelles forestières aux abords de l'université Nancy-1 (Nord-Est de la France). Ces femelles ont été placées individuellement dans des terrariums fournissant les conditions les plus proches possibles des conditions naturelles [7,10]. Toutes les araignées ont bien tissé une toile avec une retraite, dont le fond est recouvert d'une couche très dense de soie non cribellée. Ce travail s'est déroulé dans une salle maintenue à une température de 20 °C , éclairée par des tubes fluorescents de 100 lux sous un cycle de 12 h de lumière/12 h d'obscurité (allumage à 7 h). Des larves de grillon (Gryllus bimaculatus) ont été utilisées pour nourrir les araignées et pour tester la prédation collective des jeunes. A. ferox se satisfait pleinement de cette nourriture, d'autant plus que ce type de proies est présent dans son biotope naturel.
Les portées retenues pour le test ont été celles pour lesquelles les événements de la période maternelle (de l'incubation à la matriphagie) se sont déroulés normalement. J'ai alors suscité deux situations expérimentales différentes : (1) des groupes de 50 jeunes issus de la même portée ont été maintenus dans la toile maternelle après la matriphagie ( portées) ; (2) des groupes de 50 jeunes issus de la même portée ont été transférés dans une enceinte vide au lendemain de la matriphagie ( portées). Après leur transfert, ces jeunes se regroupent tous et construisent une toile commune dans les 10–12 jours qui suivent. Cette toile fabriquée par les jeunes diffère de la toile maternelle par sa taille réduite, sa plus faible densité en soie, et par l'absence de division nette entre la retraite et la zone de capture des proies. Les jeunes araignées effectuent également leur seconde mue pendant cette période (environ une semaine après la matriphagie).
Il semble évident que, pour les jeunes impliqués dans la situation 2, il y a des coûts d'isolement de la toile maternelle et de construction de la toile de toutes pièces. Cette expérience inclut ces coûts, que les jeunes qui choisissent la stratégie consistant à quitter le maternel devraient bien envisager dans la nature.
Le test a été réalisé quatre jours après la seconde mue (les mues des jeunes sont très synchronisées, la durée de la deuxième mue pour une portée totale est en moyenne de 22 h [21]). Un grillon de 20 mg , lequel constitue une proie potentielle 10 fois plus grosse que chacun des jeunes, a été déposé sur la toile (le poids moyen d'un jeune de cet âge est de 1,8 à 2,5 mg, selon les portées). Cette proie provoque une prédation collective bien caractéristique dans la plupart des portées. À chaque fois, je me suis efforcée d'introduire la proie dans des conditions identiques, notamment en le plaçant dans la toile à une distance de 6–10 cm par rapport au groupe des jeunes.
Des enregistrements vidéo ont été réalisés pour toutes les portées étudiées. Le nombre de portées où le grillon a été paralysé au bout des 30 min que dure le test et le nombre de jeunes de la portée ayant participé à la prédation (c'est-à-dire ayant contacté au moins une fois le grillon) ont été relevés. Pour analyser l'enchaînement des contacts et le maintien de la proie par les jeunes, j'ai relevé les délais successifs mis par les jeunes pour contacter et maintenir la proie à partir de l'introduction du grillon dans la toile. « Contact 1 (2, 3, 4) » représente le temps mis par le premier jeune (puis le deuxième, puis le troisième, puis le quatrième) de la portée pour toucher la proie, et « Maintien 1 (2, 3, 4) » représente le temps mis par le premier jeune (puis le deuxième, puis le troisième, puis le quatrième) pour saisir la proie avec ses chélicères pendant plus d'une seconde.
Le nombre de manifestations agonistiques (fuites, menaces, attaques) entre les jeunes de chaque portée a été relevé pendant les 30 min qui ont suivi le dépôt du grillon dans la toile. J'ai considéré qu'il y a eu « fuite » quand un jeune s'est enfuit à l'approche d'un autre jeune, « menace » quand un jeune a levé sa première paire de pattes face à un autre jeune, et « attaque » quand un jeune a dressé ses chélicères face à un autre jeune et est entré en contact brutal avec lui.
Les comparaisons des données obtenues entre les deux situations ont été réalisées en utilisant le logiciel StatView 4.0 pour Macintosh. Le test du a été utilisé pour comparer le nombre de portées qui ont paralysé le grillon, et le test U de Mann–Whitney pour comparer l'enchaînement des différentes étapes de la capture : le temps mis pour le premier contact, le temps mis pour le premier maintien, les délais entre les contacts et entre les maintiens. Le nombre moyen de participants à la capture dans les deux situations a aussi été comparé par un test U de Mann–Whitney. Une ANOVA permet de comparer le temps pris par les contacts successifs et par les maintiens successifs.
3 Résultats
La prédation a été plus efficace lorsque les jeunes ont bénéficié de la toile maternelle que lorsqu'ils ont utilisé la toile qu'ils ont eux-mêmes tissée. La capacité à paralyser le grillon a été significativement différente d'une situation à l'autre. Dans la situation 1, sept portées sur 10 (70 %) ont réussi à paralyser le grillon au bout de 30 min, contre quatre portées sur 14 (28 %) dans la situation 2 (test du : , , ).
Dans les deux situations, les jeunes ont montré des activités identiques au cours de la capture : les jeunes n'ont tout d'abord manifesté aucune réaction vis-à-vis du grillon, puis ils se sont orientés et déplacés dans sa direction, avant de le contacter et le saisir. Dans toutes les portées étudiées, les jeunes ont réalisé au moins un contact et un maintien (saisie de la proie pendant plus d'une seconde). Cependant, le nombre moyen de participants à la capture a été plus important dans la situation 1 que dans la situation 2 (Tableau 1).
Comparaisons des activités prédatoires entre la situation 1 (sur la toile maternelle) et la situation 2 (sur la toile des jeunes)
| Situation 1 ( portées) | Situation 2 ( portées) | Test U de Mann–Whitney | |
| Nombre de participants1 | 17,6 ± 6,5 | 8,7 ± 5,9 | p<0,005, U=21,0 |
| Temps mis pour le premier contact2 | 198 ± 117 | 323 ± 252 | NS, U=51,0 |
| Temps mis pour le premier maintien3 | 325 ± 175 | 432 ± 248 | NS, U=54,0 |
| C1–C24 | 65 ± 94 | 126 ± 111 | p=0,035, U=34,0 |
| M1–M25 | 83 ± 148 | 249 ± 318 (n=13) | p=0,038, U=31,5 |
1 Nombre de participants (moyenne ± écart type) à la capture ;
2 délai (en secondes, moyenne ± écart type) entre le dépôt de la proie et le premier contact ;
3 délai (en secondes, moyenne ± écart type) entre le dépôt de la proie et le premier maintien ;
4 délai (en secondes, moyenne ± écart type) entre le premier et le deuxième contact (C1–C2) ;
5 délai entre le premier et le deuxième maintiens (M1–M2).
Le délai entre le début du test (dépôt du grillon) et le premier contact n'a pas différé entre les deux situations, mais aussi comme le délai au premier maintien (Tableau 1). En revanche, le délai entre le premier et le second contact, de même que le délai entre le premier et le second maintien ont été significativement plus courts dans la situation 1 que dans la situation 2 (Tableau 1). Les jeunes venus successivement se mettre en contact avec la proie sont arrivés significativement plus vite sur la proie dans la situation 1 que dans la situation 2 (Fig. 1 ; ANOVA : , ). De plus, les maintiens successifs ont été plus rapides dans la situation 1 que dans la situation 2 (Fig. 2 ; ANOVA : , ). Les actions prédatrices des jeunes sur la toile maternelle se sont enchaînées plus vite que celles des jeunes sur leur propre toile.
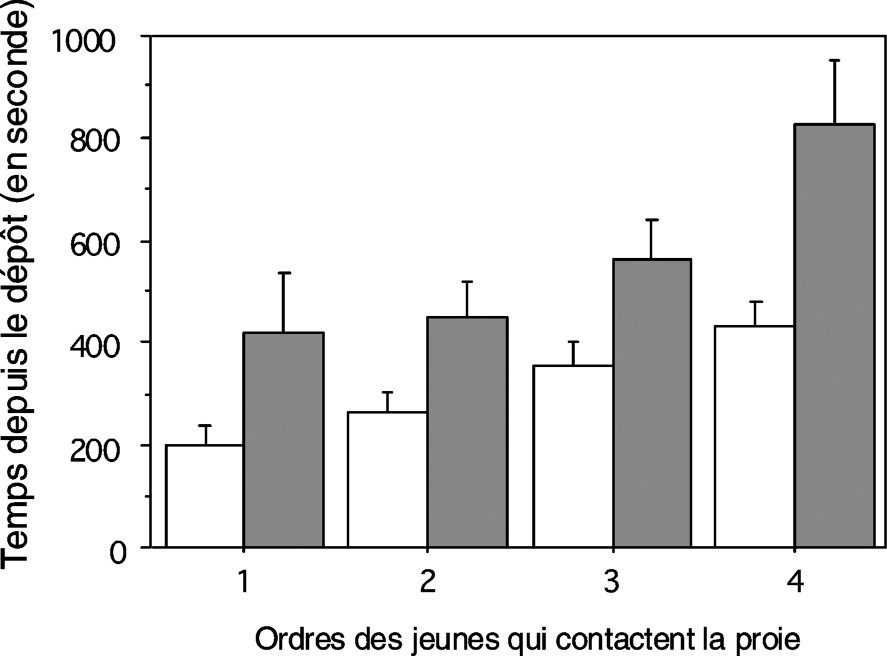
Délais (moyenne ± erreur standard) entre le dépôt de la proie dans la toile et chacun des quatre premiers contacts dans les situations 1 (□, sur la toile maternelle, n=10 portées) et 2 (■, sur la toile des jeunes, n=14 portées).
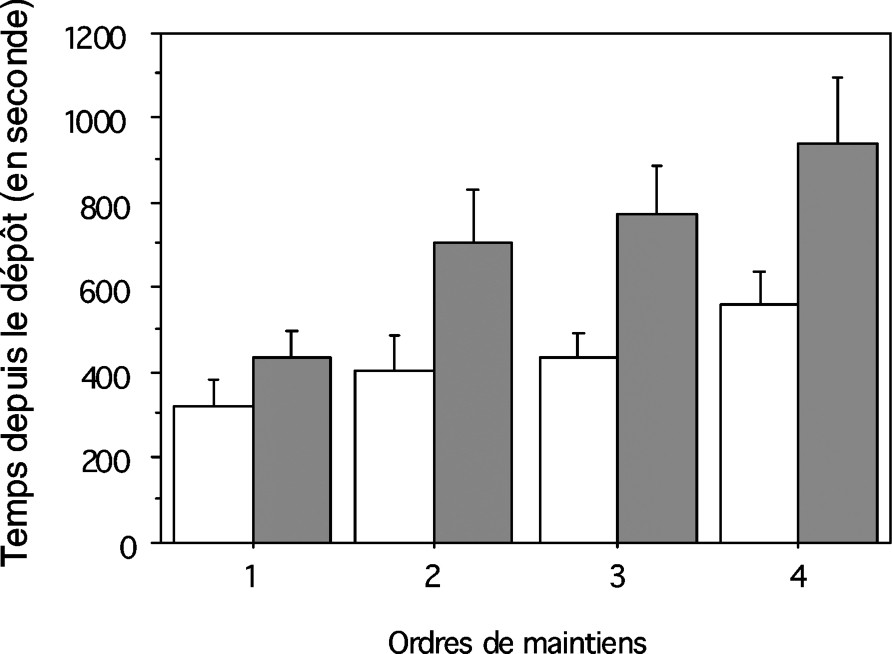
Délais (moyenne ± erreur standard) entre le dépôt de la proie dans la toile et chacun des quatre premiers maintiens dans les situations 1 (□, sur la toile maternelle, n=10 portées) et 2 (■, sur la toile des jeunes, n=14 portées).
Au cours de la capture de la proie, les jeunes n'ont presque pas montré de manifestations agonistiques entre eux. Au total, j'ai observé quatre fuites dans la situation 1, et une fuite et trois menaces dans la situation 2.
4 Discussion
L'ensemble des résultats suggère la valeur adaptative de la toile maternelle d'A. ferox, évaluée en termes d'efficacité de la capture collective des jeunes pendant la période sociale post-maternelle. Elle présente un grand avantage pour la capture d'une grande proie par rapport à la situation où les jeunes quittant le nid maternel capturent la proie dans leur toile commune. La capacité des jeunes à paralyser la proie a été plus importante dans la toile maternelle que dans la toile construite par les jeunes (70 % contre 28 %), davantage d'individus participant à la capture.
La mesure quantitative de la toile maternelle et de la toile des jeunes n'a pas été faite dans cette étude. Cependant, la toile des jeunes se voit différente de la toile maternelle, qui présente un réseau complexe de fils cribellés, par sa taille réduite, sa plus faible densité en soie, et par l'absence de division nette entre la retraite et la zone de capture des proies (observation personnelle). La structure plus solide de la toile maternelle semble plus efficace pour transmettre entre les jeunes un signal communicateur (probablement vibratoire). Les contacts et les maintiens de la proie s'enchaînent plus rapidement entre les jeunes, même si le délai jusqu'au premier contact et au premier maintien n'ont pas été significativement différents.
La phase post-maternelle des jeunes d'Amaurobius dépend très étroitement des apports alimentaires de leur mère [22,23]. La suppression de ces apports entraîne, non seulement un retard de développement et une mortalité accrue des jeunes, mais également leur dispersion plus précocement, autrement dit une phase post-maternelle plus courte [23]. Il est probable que l'héritage de la toile maternelle, avec ces soins alimentaires, est l'un des facteurs cruciaux par lesquels la mère intervient sur la phase post-maternelle.
Chez les araignées, il existe deux formes différentes de la vie « subsociale » ou « sociale maternelle ». La première forme est représentée par Cœlotes terrestris (Agelenidae) : les jeunes restent dans le nid maternel tant que la mère est vivante. Ils quittent généralement le nid avant la mort de la mère et, mis en présence d'une mère morte, ils se dispersent immédiatement [10]. Les jeunes n'ont pas de vie sociale propre en l'absence de la mère. Cette forme se rencontre également chez d'autres espèces d'araignées subsociales [8,9]. A. ferox représente une forme différente de vie subsociale : après la matriphagie, les jeunes ne quittent pas le nid maternel, ils vivent en groupe jusqu'à la dispersion, qui a lieu en moyenne 24 jours après la matriphagie. Cette seconde forme de vie subsociale existe également chez Diaea ergandros (Thomisidae), dont les jeunes issus d'une même portée restent groupés dans le nid fait de feuilles d'Eucalyptus [24]. Evans [5] a démontré la fonction protectrice du nid construit par la mère de D. ergandros vis-à-vis de sa portée : en présence d'un prédateur, 94,6 % des jeunes restant dans le nid maternel survivent, tandis que tous les jeunes quittant le nid maternel sont tués par le prédateur. Ces jeunes de D. ergandros cannibalisent leur mère et héritent du nid maternel, où ils mènent une vie grégaire jusqu'à la dispersion [24], ce qui existe également chez A. ferox. Les jeunes de D. ergandros semblent coopérer dans la capture des proies en restant dans leur nid maternel [5].
La forme de la toile maternelle de C. terrestris ressemble à celle d'A. ferox (une retraite s'ouvre à l'arrière de la surface de capture). Ces deux espèces sont sympatriques, donc exposées aux mêmes conditions écologiques, elles sont de taille équivalente, semblent exploiter les mêmes proies et ont des cycles vitaux globalement très proches. Elles présentent toutes deux une activité maternelle très développée (pour C. terrestris, voir [25,26]). Mais, étant donné que la toile de Cœlotes est constituée uniquement de soie non cribellée, elle n'a pas le caractère adhésif qui pourrait empêcher un insecte de s'approcher de la retraite et permettre de piéger et retenir les proies [15,27]. Il est probable que la fonction protectrice de la toile maternelle de C. terrestris est moins importante que celle d'A. ferox. La capacité de la toile à immobiliser une proie est également beaucoup moins importante, un insecte pouvant très bien s'échapper de la toile de C. terrestris. Cette différence dans la toile maternelle peut être un élément explicatif de la différence de mode de vie grégaire des jeunes de ces deux espèces subsociales.
Kullmann [12] suggère que la construction d'une retraite sûre soit le premier pas vers la vie sociale chez les araignées. De même, chez les insectes, les soins maternels sont rares chez les espèces qui ne construisent pas de nid [28]. Dans les organisations sociales permanentes non territoriales, les jeunes araignées agrandissent le nid maternel après la mort de la mère [29]. En revanche, l'activité de tissage chez les jeunes d'Amaurobius semble bien limitée à l'intérieur de la toile maternelle et la taille de la toile n'augmente pas.
Acknowledgments
Je tiens à remercier Chantal Roland et André Horel pour leur aide dans la réalisation de ce travail. Je remercie également Laurent Thevenard pour ses conseils précieux. Ce travail a été partiellement supporté par le Brain Korea 21 Research Fellowship.