

Abridged English version
1 Introduction
The palynological study of the Dogger–Malm boundary in the central part of the Paris Basin showed a characteristic evolution of organic-walled microfossils assemblages [4]. Although they were consistent with scattered outcrop data, these data were essentially of regional interest, because of the nearby location of the two studied cores (named A and B). Thanks to the drilling of the ANDRA underground laboratory in Bure (Meuse department), a set of new data has been collected in the eastern part of the basin (Fig. 1). This new core (HTM 102), which also benefits from organic geochemistry studies [7], allowed us to test the reproducibility of the palynological signal at the Callovian–Oxfordian boundary in a completely different sedimentological context.
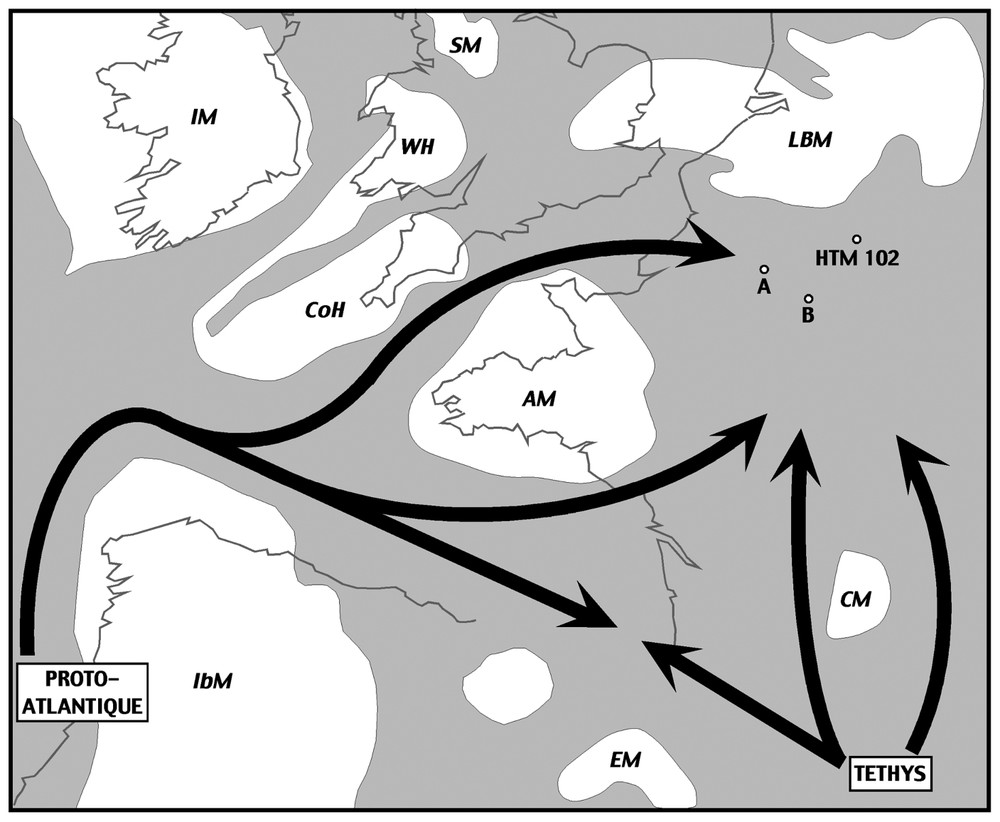
Localisation des sondages A, B et HTM 102 sur une carte paléogéographique de l'Europe occidentale au Callovien (d'après Thierry et al. [11], modifié). Les zones émergées sont représentées en blanc. AM: Massif armoricain, CM: Massif central, CoH: Cornouaille, EM: massif de l'Ebre, IbM: Massif ibérique, IM: Massif irlandais, LBM: massif Londres–Brabant, SM: Massif écossais, WH: Pays de Galles.
Location of the A, B and HTM 102 cores on a palaeogeographical map of western Europe at the Callovian (after Thierry et al. [11], modified). Emerged areas are white-coloured. AM : Armorican Massif, CM : Massif Central, CoH : Cornubian High, EM : Ebro Massif, IbM : Iberian Massif, IM : Irish Massif, LBM : London–Brabant Massif, SM : Scottish Massif, WH : Welsh High.
2 Geologic setting and sampling method
2.1 Geologic setting
The transition between Callovian and Oxfordian in the central part of the Paris Basin occurs in condensed calcareous series and is associated with ferruginous oolite deposits. In the Northeast of the Paris Basin, the same interval corresponds to the marly and dilated formation of the ‘Argiles de la Woëvre’, which seldom outcrops in good conditions. However, because it is a target for ANDRA's investigations, this formation has been reached by a series of cores, including the HTM 102 core.
2.2 Sampling method
Because of the inaccurate location of the Callovian–Oxfordian boundary in HTM 102 at the time of our sampling and according to the reduced number of samples (and cost) desirable in such a preliminary study, we decided to favour samples which benefited from organic geochemistry studies and which were surely located on both sides of the boundary. Even with such a low number of samples (8 on HTM 102, Fig. 2), the drastic palynological change observed in A and B cores [4] should be, if need be, obvious in HTM 102 too.
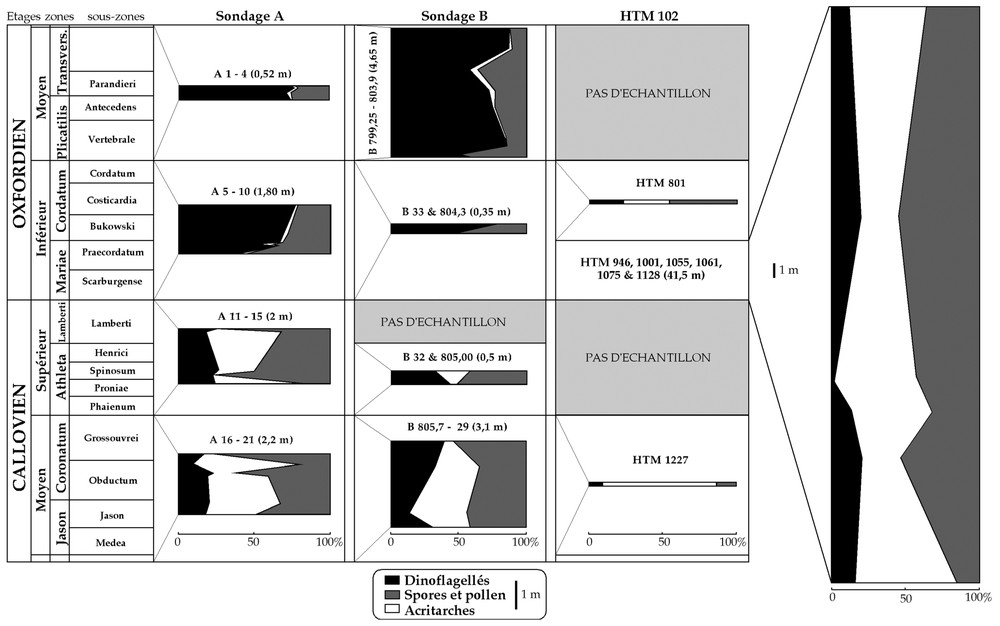
Évolution comparée de la composition des assemblages palynologiques dans trois puits des parties centrale (A et B) et orientale (HTM 102) du bassin de Paris. Les échelles verticales sont identiques pour les trois puits, sauf pour les échantillons oxfordiens du sondage HTM 102 (échelle réduite de moitié, en raison de la puissance de la série).
Comparison of the evolution of palynological assemblages in three boreholes located in the central part (A and B) and in the eastern part (HTM 102) of the Paris Basin. The vertical scale is the same for the three boreholes, except for the Oxfordian samples of HTM 102 (half-reduced scale, according to the thickness of sediment).
2.3 Correlations
Dating of samples of A and B cores having already been described [4], we will therefore focus exclusively on HTM 102 samples. On this core, ammonite (D. Marchand and J. Thierry, oral communication) and palynostratigraphic [6] data are available. Although the discoveries of new ammonites helped to improve initial dating (D. Marchand and J. Thierry, oral communication), the positioning of the Callovian–Oxfordian boundary remains controversial (between 434- and 437-m depth according to ammonites and between 410- and 420-m depth according to palynology). However, the occurrence of Cardioceras scarburgense (Mariae zone, Scarburgense sub-zone, Scarburgense horizon) around 434-m depth seems to be of paramount significance. Moreover, according to our previous work in the Paris Basin [5], the presence of dinoflagellate cysts like Trichodinium scarburghense from 440-m depth may already indicates an Early Oxfordian age, even if Wanaea fimbriata is not present under 395 meter. Therefore, we decided to assess the previously Callovian HTM 1128 sample to the Early Oxfordian. The main consequence of this reassessment is the grouping of the bulk of our samples (except HTM 1227) in the Early Oxfordian only. Such an unfortunate grouping does not compromise our comparative study, since HTM 1227 remains in the Middle Callovian, according to both palynology and ammonites.
3 Results
3.1 Organic geochemistry
The results of organic geochemistry analyses realised on HTM 102 prior to palynological investigations show a drastic difference between Callovian and Oxfordian samples [7]. Marine organic matter input predominates in Callovian, whereas Oxfordian samples are characterised by a higher terrestrial contribution. The stratigraphic reassessment of the HTM 102 samples leads us to place this change in the beginning of the Oxfordian rather than in coincidence with the Callovian–Oxfordian boundary. These results contradict the palynological observations on the A and B cores in the central part of the basin (predominant terrestrial input in the Callovian, predominant marine input in the Oxfordian) [4]. These contradictory data motivated complementary palynological investigations on HTM 102.
3.2 Palynology
The palynological analysis of HTM 102 samples does not reveal the drastic differences observed at the Dogger–Malm boundary in A and B cores (dramatic rise of dinoflagellate cysts at the expense of acritarchs and sporomorphs). Only the Callovian sample of HTM is consistent with A and B Callovian samples (high rates of acritarchs and sporomorphs versus low rates of dinoflagellate cysts). The differences between the three cores seem to be more obvious from the Lower Oxfordian, with a slight evolution of palynological assemblages in HTM 102 (lost of acritarchs to the sporomorphs). According to another palynological study in progress in the same area on Oxfordian and Kimmeridgian samples, the disappearance of acritarchs is achieved in the Middle Oxfordian, with a rise of foraminifer linings at the expense of sporomorphs. On a qualitative point of view, the replacements of cysts observed in A and B cores cannot be observed in HTM 102, probably because of the small interval of time corresponding to this series of samples and because of the relative paucity of dinoflagellate cysts in the palynological assemblages.
4 Discussion and perspective
The acritarchs abundance excepted, which should influence the organic geochemistry of Oxfordian samples and could give them a more marine signature, the tendencies inferred by both palynological and organic geochemistry data are consistent on HTM 102 core. Thus, both organic geochemistry and palynology display an obvious difference of composition of organic matter in the Oxfordian samples between HTM 102 and the two other cores. This difference could be due to several factors. First of all, the question of correlation between the three cores must be examined. Our results on scattered outcrops in Burgundy (southern margin of the Paris Basin) [5] show always the same trends as those observed in A and B cores during the Callovian–Oxfordian transition. Thus, more than a problem of correlation between condensed and comprehensive series, this observation is more likely to support a global interpretation. In this perspective, the contemporaneous connection of the proto-Atlantic with the Paris Basin [3,8] seems to be of major interest. The modifications of the palaeo-currents pattern involved by such a palaeogeographic evolution could easily be linked with production and transportation processes of organic walled microfossils [12]. Concerning the qualitative aspect of dinoflagellate cyst assemblages, the comparison of HTM 102 with the two other cores are difficult to set up, due to the paucity in dinoflagellate in HTM 102 samples. However, even if they result from a somewhat different palynological laboratory technique (e.g., sieving), the biostratigraphic data available on this core [6] seem to be consistent with the observations made on A and B cores [4]. Only few dinoflagellate cysts might show some signs of diachronism (e.g., Compositosphaeridium polonicum). Such a modification in the composition of dinoflagellate cyst assemblages at the Dogger–Malm boundary is well known in western Europe [9] and could probably be linked with a global event, like the connection of the Paris Basin with the proto-Atlantic, known to occur at that time. More surprising is the quantitative differentiation between the central and eastern parts of the basin. The abundance of acritarchs in Oxfordian samples in HTM 102 could be due to local conditions of production and/or of preservation, but the rise of sporomorphs in the same samples could also be linked with eustatic context (mostly proximal trapping during the Oxfordian sea-level rise, following a larger spreading during the relative low-stand of Callovian). Finally, the monotonous lithology of the ‘Argiles de la Woëvre’ seems to be a clue to little contrasted palaeoenvironmental conditions in the eastern part of the Paris Basin at the Dogger–Malm boundary.
5 Conclusion
The contrasted palynological signatures observed in the central and eastern parts of the Paris Basin result probably from an overlapping of local and global signals. The recognition of the respective parts of these signals could be achieved by a pluridisciplinary approach including palynology, organic geochemistry and clay mineralogy. The results that we can expect from such an approach are threefold. First, the palynological differences between eastern and central parts of the Paris Basin shall be validated by organic geochemistry data. Second, the recognition of further proofs of overlapping signals could be of major interest to decipher the organic matter preservation processes. Third, the analogy between clay particles and the smallest organic microfossils (like acritarchs) could help to understand the part of transport in organic matter distribution in sediments. Finally, the reconstitution of palaeoclimate using sporomorph assemblage composition [1,2,10] will be undertaken in order to supplement our understanding on palaeoenvironmental consequences of an event such as the connection of the proto-Atlantic with the Paris Basin, which probably occurred at the Dogger–Malm boundary.
1 Introduction
L'étude palynologique de deux forages pétroliers réalisés dans le centre du bassin de Paris montre une évolution caractéristique, tant qualitative que quantitative, des assemblages de microfossiles organiques entre le Callovien et l'Oxfordien, qui se traduit par une augmentation brutale du nombre et de la diversité des dinoflagellés au détriment des acritarches et des sporomorphes au début de l'Oxfordien [4]. Éloignés de plusieurs dizaines de kilomètres, ces puits donnent une illustration des enregistrements palynologique et sédimentaire d'une valeur essentiellement régionale, même si elle est soutenue par des données issues d'affleurements épars, en Bourgogne notamment. Les forages liés aux travaux préparatoires du site Andra de Bure (Meuse) fournissent l'opportunité d'élargir le champ des analyses palynologiques à la partie orientale du bassin de Paris (Fig. 1), permettant de tester les hypothèses émises précédemment sur les modifications des microflores à la limite Dogger–Malm, tout en bénéficiant de l'éclairage complémentaire des études de géochimie organique entreprises à cette occasion [7].
2 Cadre géologique et mode d'échantillonnage
2.1 Cadre géologique
Au centre du bassin de Paris, les sondages A et B étudiés précédemment recoupent des séries condensées et lacunaires, essentiellement carbonatées et marquées par la présence de niveaux d'oolithes ferrugineuses à proximité de la limite Dogger–Malm [4]. Dans l'Est du bassin de Paris, le passage du Callovien supérieur à l'Oxfordien inférieur intervient au sein de l'épaisse série argileuse (pluri-décamétrique) de la formation des Argiles de la Woëvre. Du fait de sa lithologie, cette série n'est que très rarement visible à l'affleurement dans des conditions satisfaisantes. Dans la perspective de l'installation d'un laboratoire consacré à l'étude du stockage des déchets radioactifs dans l'Est de la France, la formation des Argiles de la Woëvre a été atteinte par plusieurs forages Andra, dont le forage HTM 102, qui nous intéresse ici.
2.2 Mode d'échantillonnage
Dans le cadre de cette étude complémentaire, dont les coûts devaient être limités, nous avons souhaité contraindre l'échantillonnage par un certain nombre de critères. En premier lieu, nous avons privilégié les échantillons ayant bénéficié d'analyses de géochimie organique, de façon à pouvoir confronter leurs résultats aux données palynologiques. Nous avons ensuite cherché à assurer le recoupement effectif de la limite Dogger–Malm, dans un contexte où les datations fournies par la palynologie [6] et par ammonites (D. Marchand et J. Thierry, communication personnelle) s'avéraient divergentes au moment de l'échantillonnage. Nous avons donc privilégié les échantillons dont les datations paraissaient les mieux établies au moment où cette étude a débuté. Ce choix a certes réduit le nombre des échantillons éligibles et la précision de leurs datations, mais il a aussi permis de préserver nos chances de reconnaı̂tre une éventuelle modification des assemblages palynologiques, effectivement associée à la limite Dogger–Malm. Une analyse palynologique a donc été réalisée sur une série de 8 échantillons issus du sondage HTM 102 (Fig. 2).
2.3 Corrélations
Notre proposition de corrélation entre les trois puits est issue d'une synthèse des éléments de datation disponibles. Les datations des puits A et B ayant déjà été discutées [4], seul le sondage HTM 102 sera détaillé ici. Sur ce sondage, les données biostratigraphiques proviennent des ammonites (D. Marchand et J. Thierry, communication personnelle) et de la palynologie [6]. Malgré quelques modifications des datations par ammonites au gré des échantillonnages successifs, des divergences subsistent avec la palynologie à propos du positionnement de la limite entre Callovien et Oxfordien (entre 410 et 420 m pour la palynologie, entre 434 et 437 m d'après les ammonites). Après avoir pris en compte les arguments de l'une et l'autre datations, nous avons privilégié certains éléments qui nous paraissent déterminants, tels que la présence de Cardioceras scarburgense (zone à Mariae, sous-zone à Scarburgense, horizon à Scarburgense) vers 434 m de profondeur (D. Marchand et J. Thierry, communication personnelle). En comparaison, l'acmé de Wanaea fimbriata, (dinoflagellé indiquant avec certitude l'Oxfordien inférieur, zone à Mariae) paraı̂t située très haut dans la série (vers 395 m), mais il n'est pas exclu, à lire la description des assemblages palynologiques, que les échantillons situés au-dessus de 440 m soient déjà d'âge Oxfordien inférieur (fréquence de Trichodinium scarburghense, notamment [5]). Compte tenu de ces arguments, l'échantillon HTM 1128 (à 431 m de profondeur), initialement daté Callovien supérieur (zone à Athleta ou Lamberti) est donc finalement attribué à l'Oxfordien inférieur (zone à Mariae, sous-zone à Scarburgense). Malgré les précautions prises lors du choix initial, tous les échantillons prélevés sur HTM 102 se trouvent donc regroupés dans la zone à Mariae (sous-zone à Scarburgense), à l'exception de HTM 1227 (Callovien moyen) et de HTM 801 (zone à Cordatum). Ce regroupement malencontreux par rapport à nos objectifs initiaux ne nous prive cependant pas de l'opportunité d'étudier le passage du Dogger au Malm, puisque l'échantillon HTM 1227 reste clairement attribué au Callovien moyen, aussi bien par la palynologie que par les ammonites. Les résultats des corrélations entre les trois puits sont récapitulés dans la Fig. 2.
3 Résultats
3.1 Géochimie organique
Les analyses géochimiques réalisées sur HTM 102 antérieurement à l'étude palynologique ont montré l'existence d'une différenciation nette entre le Callovien et l'Oxfordien [7]. Les échantillons calloviens présentent en effet une prédominance des hydrocarbures de bas poids moléculaire, indicative d'apports essentiellement marins, tandis que l'enrichissement en composés de haut poids moléculaire des échantillons oxfordiens suggère une contribution terrestre plus marquée. Bien que la réattribution stratigraphique des échantillons de HTM 102 amène à placer la modification du signal géochimique au cours de l'Oxfordien inférieur plutôt qu'en coı̈ncidence exacte avec la limite Callovien–Oxfordien, ces résultats n'en restent pas moins opposés aux observations palynologiques faites sur les sondages A et B au centre du bassin de Paris, qui donnent des apports terrestres prédominants pour le Callovien et des apports marins prédominants pour l'Oxfordien [4].
3.2 Palynologie
Tout comme les échantillons des sondages A et B, les échantillons du sondage HTM 102 ont subi un traitement palynologique standard. Les données palynologiques quantitatives recueillies sur le seul échantillon callovien du sondage HTM 102 (HTM 1227) sont concordantes avec les données issues des sondages A et B (Fig. 2 ; forte proportion d'acritarches, spores et pollen plus abondants que les dinoflagellés). Cependant, dès l'Oxfordien inférieur, l'évolution des assemblages palynologiques du sondage HTM 102 se singularise. Dans ce sondage, en effet, la diminution du pourcentage d'acritarches est faible et n'atteint jamais l'ampleur observée dans les sondages A et B. De plus, cette évolution profite essentiellement aux sporomorphes et non aux dinoflagellés, comme dans A et B. Cette tendance est confirmée tout au long de l'Oxfordien inférieur. Si aucun échantillon plus récent que l'Oxfordien inférieur n'a été collecté sur HTM 102, une autre étude en cours sur un sondage voisin (EST 205) indique une disparition effective des acritarches à l'Oxfordien moyen, avec pour corollaire une augmentation considérable du pourcentage de basales de foraminifères, devenant prépondérantes dans les assemblages. Cette augmentation importante ne s'accompagne pas d'une hausse du pourcentage des dinoflagellés, mais d'une baisse significative des proportions de sporomorphes. Ainsi, le changement drastique reconnu dans les sondages du centre du bassin de Paris ne peut être mis en évidence vers l'est, où l'évolution des assemblages palynologiques paraı̂t très progressive et d'ampleur réduite entre le Dogger et le Malm. Les résultats géochimiques et palynologiques paraissent donc concordants sur HTM 102, à ceci près que l'abondance des acritarches, habituellement attribués aux environnements marins, devrait donner aux échantillons oxfordiens de ce sondage une signature géochimique plus marine. Il n'en est pourtant rien, l'augmentation des contributions terrestres par l'intermédiaire des spores et pollen paraissant prépondérante dans l'évolution du signal géochimique. Du point de vue qualitatif, les remplacements de kystes prédominants observés simultanément dans les deux sondages du centre du bassin [4] ne sont pas visibles dans le sondage HTM 102, où les associations paraissent très constantes d'un échantillon à l'autre. Les kystes de dinoflagellés prédominants restent (par ordre décroissant d'abondance) ceux des genres Escharisphaeridia, Sentusidinium, Adnatosphaeridium, Rhynchodiniopsis et Gonyaulacysta. Les dinoflagellés utilisés dans les sondages A et B pour établir une série de repères quantitatifs sont retrouvés dans HTM 102 (genres Compositosphaeridium, Liesbergia, Scriniodinium, Rigaudella et Systematophora), mais n'enregistrent pas de variations d'abondance significatives.
4 Discussion et perspectives
La comparaison de séries condensées et lacunaires (sondages A et B) avec une série dilatée (HTM 102) pourrait laisser craindre des corrélations abusives. Cependant, même s'il est très condensé (sans doute centimétrique dans le sondage B, [4]), l'Oxfordien inférieur est reconnu dans les puits A et B et s'accompagne d'une évolution similaire des assemblages palynologiques [4]. Ces caractéristiques paraissent d'autant moins fortuites qu'elles perdurent jusqu'à l'Oxfordien moyen et se reproduisent dans des échantillons isolés (remplissages d'ammonites) provenant de la marge méridionale du bassin. Il paraı̂t donc vraisemblable d'admettre une dissociation des évolutions palynologiques entre le centre et l'Est du bassin de Paris à partir du début de l'Oxfordien, probablement à la faveur d'une différenciation paléogéographique. Dans cette perspective, la mise en communication du bassin de Paris avec le proto-Atlantique à la faveur d'une transgression survenant à la base de l'Oxfordien inférieur, souvent évoquée dans les travaux récents [3,8], semble constituer un élément crédible d'interprétation. Les modifications induites par un tel événement dans les paléocirculations océaniques et atmosphériques paraissent pouvoir jouer un rôle important dans la répartition des palynomorphes [12]. En effet, indépendamment des facteurs climatiques et de leurs implications sur la production primaire, le transport par voie aquatique des microfossiles organiques joue un rôle majeur dans la composition des assemblages observés. Du point de vue qualitatif, les assemblages de dinoflagellés présentent un cachet caractéristique de la limite Dogger–Malm, mais il reste difficile de reconnaı̂tre, dans le forage HTM 102, les relais déjà observés dans les sondages A et B [4]. Cette difficulté tient probablement à la faible valeur statistique des rares dinoflagellés rencontrés dans les échantillons de ce sondage. La lecture du rapport d'étude palynostratigraphique du sondage HTM 102 [6] apporte des éléments de réponse à cette question. Le mode préparatoire des échantillons traités dans ce cas diffère du mode préparatoire que nous avons utilisé. Les procédés de concentration des kystes de dinoflagellés (tamisages notamment) qui ont été utilisés ne sont pas envisageables dans le cadre d'analyses quantitatives visant à préserver toutes les catégories de palynomorphes, y compris les acritarches. Pour l'intervalle Callovien supérieur–Oxfordien inférieur, les données disponibles indiquent que la base du Callovien supérieur est marquée par l'apparition des kystes de Liesbergia liesbergensis ([4], « repère 2 »), que le passage dans l'Oxfordien inférieur est caractérisé par l'apparition de Wanaea fimbriata et que la fin de l'Oxfordien inférieur serait riche en kystes de Systematophora areolata ([4], « repère 3 »). Les kystes de faible valeur biostratigraphique ne sont pas signalés dans ce rapport, nos propres résultats montrant cependant que les genres communs tels Escharisphaeridia et Sentusidinium sont bien représentés. Parmi les indications d'abondance, l'acmé de Compositosphaeridium polonicum semble contemporaine de celle de Rigaudella aemula dans HTM 102, alors que ces deux acmés étaient nettement séparées au cours du temps dans les sondages A et B, avec respectivement le début du Callovien moyen et la fin de l'Oxfordien inférieur pour C. polonicum et R. aemula. Dans ce cas précis, on peut soupçonner un diachronisme dans l'acmé de C. polonicum à l'échelle du bassin de Paris. En résumé, une étude détaillée de l'abondance des kystes de dinoflagellés montrerait que l'évolution dans le temps des assemblages de ces microfossiles est similaire, à quelques exceptions près, dans les parties centrale et orientale du bassin de Paris. L'impossibilité de reconnaı̂tre les repères biostratigraphiques définis précédemment au centre du bassin s'expliquerait par le faible nombre d'échantillons prélevés dans le cadre de cette étude complémentaire. L'analyse palynologique met donc en évidence une modification des proportions de palynomorphes au sein des assemblages, entre l'Est et le centre du bassin de Paris, sans que la composition intrinsèque des assemblages de kystes de dinoflagellés ne soit profondément altérée. Une modification à grande échelle des paléocourants et, par là même, des conditions trophiques et de transport paraı̂t pouvoir expliquer le renouvellement qualitatif généralisé qui intervient au sein des dinoflagellés à la limite Dogger–Malm (connu à l'échelle de toute l'Europe occidentale [9]). Il reste néanmoins à comprendre comment la mise en communication du bassin de Paris avec le proto-Atlantique peut s'accompagner de modifications palynologiques quantitatives de nature et d'ampleur variables suivant les régions. Le maintien des acritarches dans la partie orientale du bassin, par exemple, tient probablement à des conditions locales, difficiles à préciser, puisque nous ignorons presque tout des affinités taxinomiques et donc de la paléoécologie de ces microfossiles. De même, l'augmentation des proportions de spores et pollen à l'Oxfordien dans l'Est du bassin uniquement pourrait être due à la proximité des terres émergées (massif Londres–Brabant), ou au contexte eustatique, avec un piégeage proximal préférentiel lors d'une remontée du niveau marin à l'Oxfordien, à la suite d'une période d'épandage plus vaste durant un plus bas niveau relatif au Callovien. Enfin, les signatures palynologique et lithologique monotones des Argiles de la Woëvre semblent révélatrices de conditions paléoenvironnementales peu contrastées, propres à la partie orientale du bassin de Paris à la limite Dogger–Malm.
5 Conclusion
La différenciation du signal palynologique entre les parties centrale et orientale du bassin de Paris à partir de l'Oxfordien pourrait indiquer une superposition de signaux de valeurs globale et régionale. L'appréciation des parts respectives de ces signaux, pour difficile qu'elle soit, devrait aider à comprendre certains des mécanismes qui concourent à la répartition et à la préservation des matières organiques dans les bassins sédimentaires, depuis leur production jusqu'à leur enfouissement, en passant par leurs divers modes de transport. L'existence d'un signal contrasté en géochimie organique à la limite Dogger–Malm, finalement assez concordant avec la nature du contenu palynologique, ainsi que les résultats des analyses d'argiles sur le même intervalle stratigraphique dans le sondage HTM 102 [8], incite à appliquer aux sondages A et B une démarche pluridisciplinaire intégrant la palynologie, la géochimie organique et l'analyse des cortèges argileux. Nous pouvons attendre de cette approche, soit une confirmation, soit une infirmation des distorsions mises en évidence par la palynologie entre le centre et l'Est du bassin de Paris. Tout comme pour la palynologie, nous rechercherons d'éventuelles superpositions de signaux globaux et régionaux dans les enregistrements organiques et sédimentaires. Si la palynologie paraı̂t apte à donner des éléments d'appréciation du transport ou de la production primaire, la géochimie organique pourrait, quant à elle, donner une vision complémentaire de l'origine et de la maturité de la matière organique, aussi bien amorphe que figurée. Par ailleurs, si l'on considère que, par leur petite taille, les minéraux détritiques argileux constituent un analogue minéral de certains éléments organiques également microscopiques, tels que les acritarches, l'analyse des argiles, en précisant les variations des apports terrigènes au cours de l'intervalle Callovien–Oxfordien, pourrait fournir des éléments d'appréciation du transport des particules de petite taille, aussi bien organiques que minérales. Enfin, l'application de méthodes d'analyse du paléoclimat à partir des spores-pollen et des dinoflagellés [1,2,10] sera entreprise, afin de rechercher d'éventuelles traces de modifications du paléoclimat, enregistrées par les flores fossiles et susceptibles d'améliorer notre compréhension des conséquences paléoenvironnementales d'un événement tel que la connexion du proto-Atlantique avec le bassin de Paris, probablement intervenue à la limite Dogger–Malm.
Remerciements
Nous remercions Elf-Aquitaine (désormais TotalFinaElf) et l'Andra de nous avoir autorisé l'accès aux carottes des sondages A, B et HTM 102.