

Abridged English version
The study of abundant ammonite faunas collected in the eastern Paris Basin (France) in nine deep bore-holes and two open wells made by the ‘Agence nationale pour la gestion des déchets radioctifs’ (ANDRA) near Bure (Meuse Department) has allowed the following to be deduced:
- – a precise bio-chronostratigraphic calibration (Fig. 1) of the Callovian–Oxfordian clay rich facies of the ‘Argiles de la Woëvre’, ‘Terrain à chailles’ and ‘Marnes des Éparges’ formations;
- – the correlation between the boreholes and wells by means of the ammonite biostratigraphic units, which often have the precision of a faunal horizon, very frequently a sub-biozone precision and sometimes even have the precision of a biozone (Figs. 2, 3);
- – to ascertain the outstanding disparity of the sedimentary registration of each of the biostratigraphic units in the lithologic succession;
- – finally, to undertake a palaeoecological and palaeobiogeographical investigation into the ammonite faunal associations in order to highlight their palaeoenvironmental and palaeogeographical applications.
- – the position of the interpositum horizon [55], the uppermost Middle Callovian sub-unit (Coronatum Zone, Grossouvrei Subzone) and its importance for the localization of the Middle Callovian–Upper Callovian boundary;
- – the respective importance of the paucicostatum faunal horizon [32], the uppermost sub-unit of the Upper Callovian (Lamberti Zone, Lamberti Subzone) and the thuouxensis faunal horizon, the lowermost sub-unit of the Lower Oxfordian (Mariae Zone, Scarburgense Subzone) for the precise localization of the Callovian–Oxfordian boundary [28–30];
- – the validity of the woodhamense faunal horizon based on the morphological variability of the corresponding species-index that corroborates the stratigraphic situation of this sub-unit above the scarburgense horizon in the Scarburgense Subzone, Mariae Zone of the Lower Oxfordian [28–30];
- – the consequences of the morphological variability of the successive species Cardioceras praemartini, C. alphacordatum, C. praecordatum and their selection as species-index for the three faunal horizons that subdivide the Praecordatum Subzone, Mariae Zone, Lower Oxfordian [28–30];
- – the difficulties that arise when trying to recognize the Cordatum Zone and the Plicatilis Zone because of the scarcity, if not the total lack of ammonites that are characteristic of these units, which have consequences on the location of the Lower Oxfordian–Middle Oxfordian boundary.
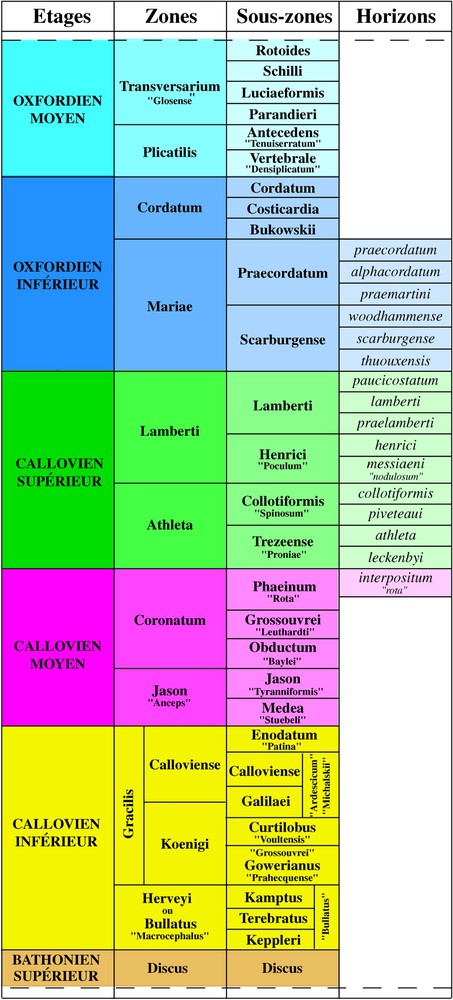
Biozonation d'ammonites du Callovien inférieur p.p. à l'Oxfordien moyen p.p. (modifié d'après [7,54]). Échelle composite vis-à-vis des provinces biogéographiques subtéthysienne et subboréale ; le nom d'unité biostratigraphique utilisé dans le texte est en grand caractère, quelle que soit la province paléobiogéographique ; son équivalent dans l'autre province est entre guillemets.
Ammonites biozonation of the Early Callovian p.p. to Middle Oxfordian p.p. (modified from [7,54]). Composite scale mixing Subtethyan and Subboreal biostratigraphic units; the name of the unit used in the text is in normal font, whatever the palaeobiogeographic province concerned; its equivalent in the other province is between brackets.
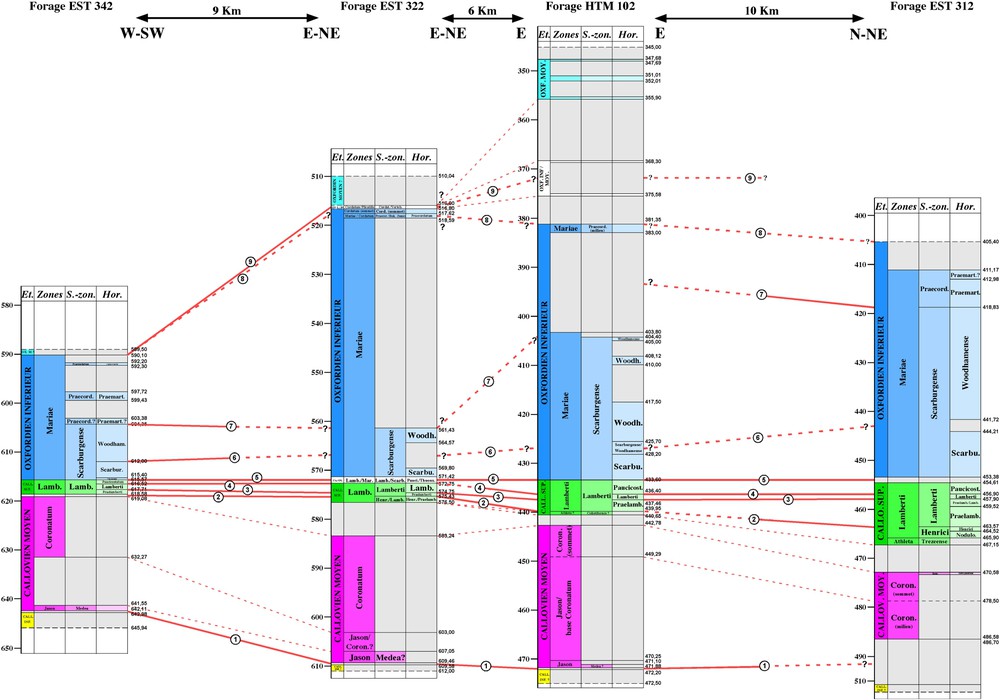
Transect WSW–ENE de corrélation entre les forages EST 342, EST 322, HTM 102 et EST 312. La limite Callovien–Oxfordien est prise comme repère horizontal. Principales lignes de corrélations (traits pleins et traits pointillés épais) : (1) base de la sous-zone à Medea (limite Callovien inférieur–Callovien moyen) ; (2) base de l'horizon à praelamberti ; (3) base de l'horizon à lamberti ; (4) base de l'horizon à paucicostatum ; (5) base de la sous-zone à Scarburgense (limite Callovien–Oxfordien) ; (6) base de l'horizon à woodhamense ; (7) sommet de la sous-zone à Scarburgense ; (8) sommet de la zone à Mariae ; (9) limite Oxfordien inférieur–Oxfordien moyen probable. Les traits pointillés fins ne sont pas des lignes de corrélations mais seulement des lignes repères.
WSW–ENE correlation profile between EST 342, EST 322, HTM 102 and EST 312 boreholes. The Callovian–Oxfordian boundary is chosen as a horizontal datum line. Main correlation lines (thick continuous and dotted lines): (1) base of the Medea Subzone (Lower Callovian–Middle Callovian boundary); (2) base of the praelamberti horizon ; (3) base of the lamberti horizon; (4) base of the paucicostatum horizon; (5) base of the Scarburgense Subzone (Callovian–Oxfordian boundary); (6) base of the woodhamense horizon; (7) top of the Scarburgense Subzone; (8) top of the Mariae Zone; (9) probable Lower Oxfordian–Middle Oxfordian boundary. The thin dotted lines are not correlation lines, but only marker lines.
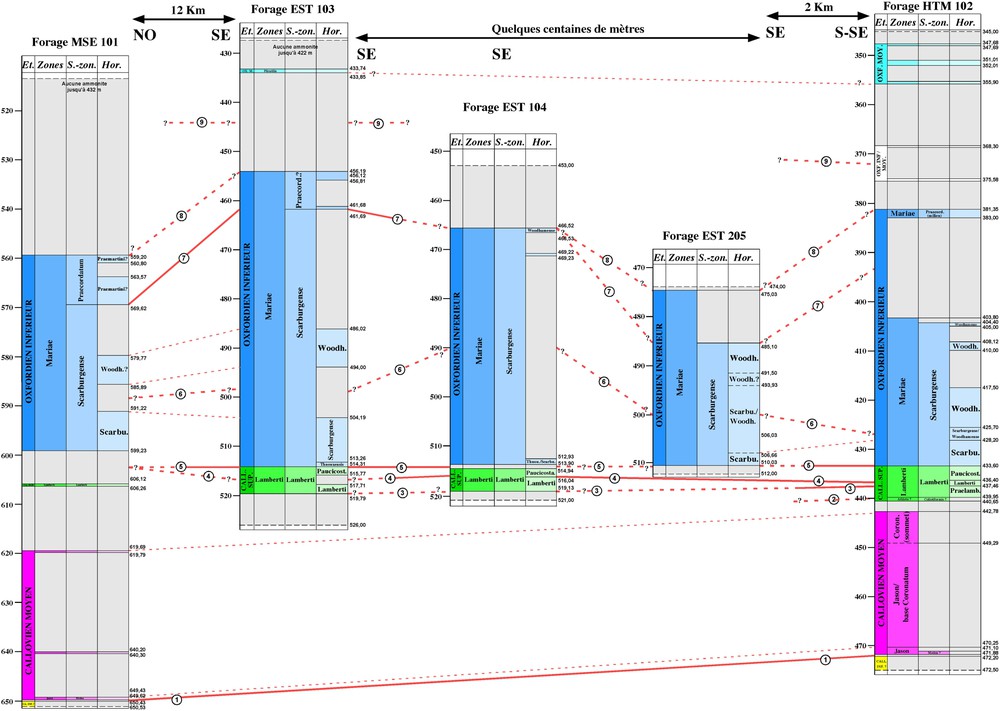
Transect NNW–SSE de corrélation entre les forages MSE 101, EST 103, EST 104, EST 205 et HTM 102 (même légende que la Fig. 2). Principales lignes de corrélations (traits pleins et traits pointillés épais) : (1) base de la sous-zone à Medea (limite Callovien inférieur–Callovien moyen) ; (2) base de l'horizon à praelamberti ; (3) base de l'horizon à lamberti ; (4) base de l'horizon à paucicostatum ; (5) base de la sous-zone à Scarburgense (limite Callovien–Oxfordien) ; (6) base de l'horizon à woodhamense ; (7) sommet de la sous-zone à Scarburgense ; (8) sommet de la zone à Mariae ; (9) limite Oxfordien inférieur–Oxfordien moyen probable. Les traits pointillés fins ne sont pas des lignes de corrélations mais seulement des lignes repères.
NNW–SSE correlation profile between MSE 101, EST 103, EST 104, EST 205, and HTM 102 boreholes. Main correlation lines (thick continuous and dotted lines): (1) base of the Medea Subzone (Lower Callovian–Middle Callovian boundary); (2) base of the praelamberti horizon ; (3) base of the lamberti horizon; (4) base of the paucicostatum horizon; (5) base of the Scarburgense Subzone (Callovian–Oxfordian boundary); (6) base of the woodhamense horizon; (7) top of the Scarburgense Subzone; (8) top of the Mariae Zone; (9) probable Lower Oxfordian–Middle Oxfordian boundary. The thin dotted lines are not correlation lines, but only marker lines.
The correlations between bore-holes and wells, and comparison with nearby outcrops [12–14,16,19,20,36,49], underline that this part of the North Peri-Tethyan intra-cratonic platforms that belong to the Paris Basin has been affected from the Middle Callovian to the beginning of the Middle Oxfordian by an unceasing subsidence, which varied in both space and time. The subsidence seems to be stronger near the centre of the basin than near its margins, with a maximum subsidence taking place during the Lower Oxfordian, Mariae Zone, Scarburgense Subzone, woodhamense horizon.
The Middle Callovian shows a southwest to northeast thickness variation from respectively 25 to 45 m (Figs. 2 and 3). However, the ammonite zones are differently positioned in the marly-clayey deposits. The Jason Zone may be absent or is reduced to few decimetres of marls; the Medea Subzone is the only sub-unit recognized. Conversely, the Coronatum Zone is well developed and varies from 22 to 45 m, although none of its sub-zones can be recognized. This is very different when compared with the outcrops located about 30 km more to the southwest, where the Middle Callovian is reduced to just a few metres or decimetres of marly limestones, sometimes with ferruginous oolites.
The whole Upper Callovian is reduced to 3 m in the eastern borehole (Figs. 2 and 3). Although, it reaches 12 m in the western borehole; gaps and faunal concentrations are frequent. The Athleta Zone can only be recognized in the northeast, preserved in a section of marls and marly limestones; in outcrops it may be either marly or included in the uppermost beds of a ferruginous oolite facies. The thickness of the Lamberti Zone varies from 3 m in the southwest to 12 m in the northeast. Within the Lamberti Zone, the Henrici Subzone is always reduced to few decimetres of clays and only recognized in the central and eastern part of the study area. The Lamberti subzone is well developed; the three faunal horizons praelamberti, lamberti and paucicostatum are frequently recognized. In the nearest southwestward outcrops, the Lamberti Zone is 6 m thick and marly-clayey, but it cannot be subdivided. More to the southwest, it is reduced to a few decimetres of marly limestones, with ferruginous oolites.
The Lower Oxfordian shows the maximum development of the clay deposits (Figs. 2 and 3). Once more, the minimum thickness is registered in the southwestern part of the study area and the maximum one in the northeastern part. The Mariae Zone is the more developed biostratigraphic unit; it varies from 25 m to 67 m. Within it, the Scarburgense Subzone is always thicker than the Praecordatum Subzone. The thuouxensis horizon is generally reduced (0.10 to 1 m) and is sometimes not recognized. The scarburgense horizon is thicker (3 to 20 m) and always present. The woodhamense horizon is the best developed, with an estimated maximum thickness of 32 to 52 m in the centre of the study area. The Praecordatum Subzone is not well documented; only the praemartini horizon is recognized. The total thickness of this horizon varies from 8 to 15 m. In the outcrops, the Mariae Zone is lesser developed, but its sub-zones and horizons are all recognized and proportionally developed. The Cordatum Zone is always reduced to some decimetres or meters of marls and marly limestones; its real thickness cannot be properly measured and the sub-zones and horizons cannot be easily recognized.
The Middle Oxfordian is always badly documented and its sedimentary development cannot be well measured in the whole boreholes. The ammonites are very rare and almost non-characteristic of any kind of a biostratigraphic unit; the Vertebrale Subzone and the Plicatilis Zone are the only ones recognized in some boreholes.
The palaeoecological interpretation of the ammonite faunal associations in the eastern part of the Paris Basin indicates a medium palaeodepth (lower offshore), certainly a little more marked near the centre of the ‘basin’ than on its extremities. If not considering the subsidence effects, this palaeodepth does not seem dissimilar to that of the southeastern basin of France (Subalpine areas) [21]. In comparison with the Paris Basin, the proximity of the former with the Tethyan oceanic domain allows us to deduce that variations of the ammonite assemblages may be related to the different mode of life of the collected taxa [22–24,33,35,56]. Especially, if compared to the Ammonitina taxa, the selective distribution of the Phylloceratina and Lytoceratina is characteristic of open-sea distal platforms [38,40,41].
From a palaeobiogeographical point of view, the Callovian–Oxfordian deposits of the eastern Paris Basin have recorded the two boreal southward ammonite invasions that characterize the Middle–Upper Jurassic transition: firstly, the Kosmoceratidae during the Middle–Upper Callovian, and, secondly, the Cardioceratidae [5,15,52] during the Upper Callovian and Lower Oxfordian. These two invasions gave rise to heterogeneous biogeographic areas, the so-called Subboreal province and Sub-Mediterranean province, which are in a geographically intermediate location, between the northern Boreal province and the southern Tethyan Province [25,50]. Considered as faunal transition areas, the Subboreal and Sub-Mediterranean provinces have composite ammonite associations and their space and time extensions and ranges are variable.
1 Introduction
Parmi les études entreprises depuis 1990 par l'Andra pour l'implantation d'un site de stockage de déchets radioactifs dans l'Est du bassin de Paris, les forages effectués sur le revers des côtes de Meuse, entre Joinville (vallée de la Marne) et Gondrecourt-le-Château (vallée de l'Orbain) ont traversé les « argiles de la Woëvre » ou « argiles callovo-oxfordiennes » (Callovien moyen p.p. à Oxfordien inférieur p.p.). Par rapport aux forages pétroliers, ils ont l'originalité d'avoir carotté les séries argileuses avec un taux élevé de récupération, livrant ainsi une abondante faune d'ammonites, alors que ces assises étaient délaissées au seul profit des unités calcaires susceptibles de renfermer des hydrocarbures.
Cet ensemble argileux repose sur une formation strictement carbonatée (Bathonien terminal–Callovien inférieur p.p.), dont les membres sont rangés sous le terme commode, mais quelque peu désuet, de « Dalle nacrée » [27,53] ou « Dogger calcaire ». Le toit de cette formation est marqué par une surface durcie et/ou perforée, une discontinuité d'extension géographique et d'importance stratonomique majeures à l'échelle du bassin de Paris.
Au-dessus des « argiles de la Woëvre » viennent les « terrains à Chailles » (Oxfordien inférieur p.p.), formation encore assez argileuse, mais avec intercalations plus carbonatées et nodules siliceux, et les « marnes des Éparges » (Oxfordien moyen p.p.), représentant le « faciès argovien » des auteurs [31,37]. Enfin, encore au-dessus, les séries redeviennent strictement carbonatées avec des calcaires variés dont des formations à coraux (Oxfordien moyen p.p. et supérieur), les faciès « glypticien » et « rauracien » des auteurs [8,26].
Quatre forages sont localisés sur le site même de l'usine de stockage, à 2 km au sud du village de Bure et à mi-distance entre Saudron et Mandres-en-Barrois : EST 103, EST 104, EST 204 et EST 205. Aux très nombreux fossiles des forages, il faut ajouter ceux récoltés sur les verses lors du creusement du puits principal d'accès (PPA), sur l'emplacement de EST 204, et du puits auxiliaire (PAX), sur celui de EST 205. Cinq autres forages répartis autour du site ont aussi fourni un abondant matériel : HTM 102, 2 km au sud–sud-est, près de Cirfontaines-en-Ormois (Haute-Marne) ; MSE 101, à 11,5 km au nord–nord-ouest, entre Morley et Couvertpuis (Meuse) ; EST 322 et EST 342, à l'ouest–sud-ouest, respectivement à 8,5 km, près de Montreuil-sur-Thonnance (Haute-Marne), et 20 km, à Nomécourt (Haute-Marne), au-delà de Joinville et de la vallée de la Marne ; EST 312, à 1 km vers le nord-est, aux abords de Demanges-aux-Eaux (Meuse).
2 But de l'étude et méthode appliquée
Une recherche systématique des ammonites des séries argileuses a été effectuée entre le toit de la « Dalle nacrée » et le mur des « terrains à Chailles ». Son but était de replacer toutes les données physico-chimiques fournies par ailleurs (lithologie, diagraphie, géochimie, susceptibilité magnétique, etc.) dans un cadre temporel fondé sur des données biochronologiques, afin de les corréler le plus précisément possible.
Les carottes ont été « disséquées » sur une longueur cumulée de 790 m pour 9 forages, auxquels s'ajoutent les verses des puits. Les ammonites collectées, matériel fondamental en biostratigraphie du Jurassique, accompagnées de lamellibranches et de brachiopodes, a impliqué un délitage centimétrique des carottes afin d'obtenir le maximum d'informations.
Cette collecte de macrofossiles, banale sur le terrain, mais trop rarement utilisée pour les forages, a permis de constituer une base de données paléontologiques de plusieurs centaines de spécimens. Grâce aux cotes de profondeur et aux logs de forages, chaque fossile, en fonction de sa préservation totale ou partielle, a été positionné au centimètre près dans la pile sédimentaire, puis déterminé le plus précisément possible au niveau du genre, du sous-genre et de l'espèce, voire de la sous-espèce ou du morphe.
Fondées sur les apparitions, disparitions, concomitances et abondance des divers taxons répertoriés, les procédures opératoires de la biochronologie ont été appliquées [51]. La recherche dans les successions des taxons caractéristiques a permis de reconnaître et délimiter très précisément les unités biostratigraphiques (zone, sous-zone et horizon), afin de déterminer l'âge de la pile sédimentaire, en référence à une échelle biozonale d'ammonites.
Selon les forages et les faunes recueillies, toutes les unités biostratigraphiques ne sont pas reconnues. Ceci n'implique pas forcément leur absence, mais montre que la précision de datation (zone, sous-zone, horizon) et les possibilités de corrélation varient en conséquence. De plus, il s'est avéré que des intervalles sédimentaires, heureusement le plus souvent peu épais, n'ont pas fourni d'ammonites caractéristiques, ou étaient non fossilifères. Dans ce cas, en fonction du niveau de précision des unités biostratigraphiques les encadrant, ces intervalles sont affectés d'un degré d'incertitude qui intervient évidemment dans la précision des datations et des corrélations.
Enfin, pour avoir une vision plus générale du secteur concerné du bassin de Paris, à ces données nouvelles et fondamentales, sont venues s'ajouter les connaissances régionales existantes acquises uniquement à partir d'affleurements [11–17,20,36,43,49,58].
Au bilan, l'apport des faunes d'ammonites des forages Andra concerne les thèmes suivants :
- – le calibrage biochronostratigraphique de la série argileuse callovo-oxfordienne ;
- – les intérêts paléoécologique et paléobiogéographique des associations fauniques ;
- – leurs applications paléoenvironnementales et paléogéographiques.
3 Biostratigraphie, biochronologie et calibrage de la série argileuse callovo-oxfordienne
3.1 Choix de l'échelle biostratigraphique de référence
L'échelle biozonale d'ammonites utilisée résulte d'un consensus international pour l'Europe occidentale (Fig. 1). Constamment affinée [45], elle est fondée sur de très nombreuses données réparties sur l'ensemble des régions nord-ouest péri-téthysiennes. Pour la France, elle a été établie par le Groupe français d'étude du Jurassique pour tout le système, dont les étages Callovien [53] et Oxfordien [7] intéressent la pile sédimentaire traversée par les forages. Concernant le Callovien inférieur, qui correspond aux calcaires de la « Dalle nacrée », aucune ammonite n'a été récoltée dans les forages ; ce sont les données des affleurements qui ont été transposées aux forages.
3.2 La limite Callovien moyen–Callovien supérieur et l'horizon à interpositum
Les ammonites sont abondantes dès la base des « argiles de la Woëvre » qui débutent au Callovien moyen, voire localement dès l'extrême sommet du Callovien inférieur. Les Kosmoceratidae, Pachyceratidae et Hecticoceratidae sont bien représentés, accompagnés de rares Perisphinctidae et Reineckeidae. Cette faune confirme l'existence en France de l'horizon à interpositum qui souligne la limite Callovien moyen–Callovien supérieur. Cette unité est caractérisée par les premiers Kosmoceras s.str. à ornementation de côtes externes fasciculées ; son équivalent en Angleterre est l'horizon à phaeinum [6]. Fondé sur la sous-espèce Kosmoceras zugium interpositum (Buckman), ce biohorizon [55] individualise le sommet de la sous-zone à Grossouvrei (zone à Coronatum, Callovien moyen), où ce taxon est associé aux derniers et rares individus de l'espèce indice Erymnoceras coronatum (Bruguière). Reconnu dans EST 312 (cote 470,08 m), cet horizon existe dans le Jura, le Nord Bourgogne et le Sud Champagne [19].
3.3 Le Callovien supérieur : zone à Athleta et zone à Lamberti
La rareté ou l'absence d'ammonites, ainsi que des paléoenvironnements éliminant les brachiopodes, engendre des intervalles d'incertitude qui rendent difficile la localisation de la limite Callovien moyen–Callovien supérieur (zone à Coronatum–zone à Athleta). Dans la plupart des forages et dans les rares affleurements proches, la zone à Athleta n'est pas confirmée.
Dans HTM 102, la zone à Coronatum est reconnue dans l'intervalle 449,29 à 442,78 m ; elle est suivie par la zone à Lamberti, sous-zone à Lamberti, horizon à praelamberti, entre 439,95 m et 437,46 m ; la zone à Athleta (indices possibles entre 439,95 et 440,65 m) n'est pas confirmée. La limite Callovien moyen–Callovien supérieur serait entre 442,78 et 440,65 m, dans la première unité d'argilites silteuses des « argiles de la Woëvre ». Les associations palynologiques recensées entre 440 et 460 m (Jan du Chêne, données inédites), rapportées à un intervalle compréhensif zone à Coronatum-zone à Athleta-zone à Lamberti, ne permettent pas de corrélations fines avec les biozones d'ammonites.
Cette limite est comprise dans un mince intervalle d'incertitude, entre 619,08 et 618,58 m dans EST 342, entre des niveaux bien datés de la zone à Coronatum et de la zone à Lamberti, sous-zone à Lamberti, horizon à praelamberti.
Dans MSE 101, en l'absence d'ammonites, cette même limite serait au-dessus de la cote 619,69 m, près du sommet de la première unité d'argilites silteuses finement bioclastiques à filaments. Les rares indices du Callovien moyen sont en dessous de cette cote et le Callovien supérieur, zone à Lamberti, n'est certain qu'entre 606,12 et 606,26 m. La zone à Athleta n'a pas été caractérisée.
Un intervalle conséquent, entre 583,24 et 576,50 m, sans ammonites caractéristiques, ne permet pas de repérer le passage Callovien moyen–Callovien supérieur dans EST 322, alors que respectivement, en dessous et au-dessus de ces cotes, la zone à Coronatum et la zone à Lamberti sont bien présentes. Pour cette dernière, la distinction entre sous-zone à Henrici (horizon à henrici) et sous-zone à Lamberti (horizon à praelamberti) est délicate.
Il en est de même dans EST 312, avec un intervalle d'incertitude entre 470,58 m, toit de l'horizon à interpositum, sous-zone à Grossouvrei, zone à Coronatum et 467,15 m, mur probable de la zone à Athleta. En effet, malgré une faune peu caractéristique qui indiquerait la sous-zone à Trezeense, la zone à Athleta occuperait l'intervalle 467,15 à 465,90 m. L'horizon à nodulosum, sous-zone à Henrici, zone à Lamberti débuterait au-dessus de cette dernière cote.
Enfin, cette limite n'a été reconnue, ni dans EST 103, EST 104, EST 204 et EST 205 où la base de la série argileuse n'a pas été carottée, ni plus au sud–sud-ouest, à Humberville et Saint-Blin [12,36], où les assises correspondantes n'affleurent pas.
3.4 La limite Callovien–Oxfordien
3.4.1 L'horizon à paucicostatum (toit du Callovien supérieur ; zone à Lamberti, sous-zone à Lamberti)
La biozonation du Callovien supérieur et de l'Oxfordien inférieur est fondée sur des observations provenant du vaste domaine cratonique nord-ouest péritéthysien. Les données les plus complètes sont issues de sections couvrant totalement ou partiellement cet intervalle chronostratigraphique. Elles sont échelonnées sur un transect NW–SE, depuis l'île de Skye (Écosse), jusqu'au bassin du Sud-Est (France), en passant par le Sud de l'Angleterre (Weymouth, Dorset), le Boulonnais, l'Ardenne, le bassin de Paris et le Jura. Les recoupements et corrélations entre ces gisements et le rôle joué par plusieurs familles d'ammonites ont permis d'élaborer une échelle atteignant une précision de l'ordre de l'horizon.
La sous-famille des Cardioceratinae est le groupe référent, car il permet des corrélations entre les provinces boréale, subboréale et subméditerranéenne, qui s'étagent du nord vers le sud sur les régions ouest-européennes. Deux autres sous-familles ont permis d'établir des échelles parallèles : les Peltoceratinae [3,4] et les Hecticoceratinae [28–30]. Parfois, ces dernières donnent des précisions stratigraphiques supérieures à celle des Cardioceratinae pour les unités définies à la limite Callovien–Oxfordien et pour les deux premiers horizons de l'Oxfordien inférieur.
Dans les forages, la disparition des genres Kosmoceras, Distichoceras et Orionoides souligne bien le toit du Callovien supérieur avec l'horizon à paucicostatum, fondé sur l'espèce Cardioceras paucicostatum Lange. Ce taxon est associé à des Hecticoceras et des Peltoceratinae strictement calloviens. Primitivement considérée comme la première sous-unité biostratigraphique de l'Oxfordien [32], un accord international en fait désormais l'horizon terminal du Callovien [29,53]. Bien que toujours peu développé en épaisseur dans la série argileuse (1,50 à 2,80 m), cet horizon a été reconnu sans ambiguïté dans la presque totalité des forages : EST 342 (616,52 à 615,57 m), HTM 102 (436,40 à 433,60 m), EST 103 (515,77 à 514,31 m), EST 104 (516,04 à 514,94 m) et EST 312 (456,90 à 454,61 m). Par manque de taxons caractéristiques entre 572,75 et 571,42 m dans EST 322, il ne peut être clairement séparé de l'horizon à thuouxensis, horizon de base de l'Oxfordien qui lui succède. Un intervalle d'incertitude (606,12 à 599,23 m) ne permet pas de positionner l'horizon à paucicostatum dans MSE 101, ainsi que dans la partie inférieure des forages EST 204 et EST 205 qui n'atteignent pas ces niveaux.
3.4.2 L'horizon à thuouxensis (base de l'Oxfordien inférieur ; zone à Mariae, sous-zone à Scarburgense)
L'horizon à thuouxensis, première unité de l'Oxfordien [30] a été reconnu. Primitivement dénommé horizon à elisabethae [7,29], il était fondé sur Peltoceratoides elisabethae Jeannet ; la position stratigraphique de ce taxon s'étant révélée erronée, Hecticoceras (Brightia) thuouxensis Fortwengler, Marchand et Bonnot, espèce abondante et caractéristique de cet intervalle dans le bassin du Sud-Est, a été choisie comme nouvelle espèce indice.
Cet horizon est très fossilifère dans EST 342 (615,57 à 615,40 m) et EST 103 (514,31 à 513,26 m), avec de nombreux représentants de l'espèce indice identiques à ceux du bassin du Sud-Est. Les deux horizons limites du Callovien et de l'Oxfordien n'ont pu être distingués dans EST 322, entre 572,75 et 571,42 m ; dans EST 104, entre 514,94 et 513,90 m, les horizons de base de l'Oxfordien, horizon à thuouxensis et horizon à scarburgense, n'ont pu être dissociés. L'horizon à thuouxensis n'a pas été détecté dans MSE 101, HTM 102, EST 204, EST 205 et EST 312.
3.4.3 La limite Callovien–Oxfordien
La caractérisation ou la non-reconnaissance de l'un ou l'autre, voire des deux horizons, sous-zones ou zones frontières entre le Callovien et l'Oxfordien, conditionne le positionnement précis de la limite entre ces deux étages. Dans certains forages, l'absence d'ammonites dans un intervalle sédimentaire plus ou moins important conduit à une certaine incertitude ; cependant, cette limite est toujours localisée vers le tiers inférieur des « argiles de la Woëvre ». Elle n'a été reconnue, ni à Saint-Blin, ni à Humberville, où les assises correspondantes n'affleurent pas.
Elle est très précisément placée à 615,57 m dans EST 342 et 514,31 m dans EST 103, dans ce dernier à la base d'une passée marneuse à microfilaments ; ces cotes sont encadrées par les deux horizons limites, horizon à paucicostatum et horizon à thuouxensis, respectivement pour le Callovien et l'Oxfordien. Elle est à 433,60 m dans HTM 102, au toit d'un épisode plus silteux ; en dessous de cette cote, les ammonites illustrent parfaitement l'horizon à paucicostatum. Au-dessus, elles dénotent la présence de l'horizon à scarburgense, l'horizon à thuouxensis n'étant pas caractérisé. La limite Callovien–Oxfordien est entre 514,94 et 513,90 m dans EST 104, mince intervalle sans ammonites mais précédé et suivi d'assises marneuses à microfilaments renfermant respectivement des faunes de l'horizon à paucicostatum et des horizons à thuouxensis et scarburgense. Elle est entre 572,75 et 571,42 m dans EST 322, intervalle où il est impossible de séparer les faunes de l'horizon à paucicostatum de celles de l'horizon à thuouxensis ; une situation identique existe entre 454,61 et 453,38 m dans EST 312. Le plus important intervalle d'incertitude, au toit d'une passée marneuse bioclastique à microfilaments, concerne le forage MSE 101 ; des faunes de la sous-zone à Lamberti entre 606,26 et 606,12 m, et de la sous-zone à Scarburgense, horizon à scarburgense à partir de 599,23 m, encadrent une succession sédimentaire sans ammonites caractéristiques où se trouverait cette limite. Quant au forage EST 205, la limite Callovien–Oxfordien serait en dessous de la cote 510,00 m, à partir de laquelle l'horizon à scarburgense est reconnu. Le forage EST 204 n'intéresse pas les unités basales de l'Oxfordien.
Des datations par la palynologie (Jan du Chêne, données inédites) montrent d'importantes divergences et une précision nettement moindre, comparées à celles des ammonites. Le passage Callovien–Oxfordien est placé vers 415 m dans HTM 102, soit plus de 18 m au-dessus de la limite donnée par les ammonites, et entre 560 et 600 m dans MSE 101, c'est-à-dire dans un intervalle sédimentaire de 40 m, contre moins de 7 m avec les ammonites.
3.5 La succession horizon à scarburgense–horizon à woodhamense (Oxfordien inférieur, zone à Scarburgense, sous-zone à Scarburgense)
Fossilifère et bien développée en épaisseur de sédiments, la sous-zone à Scarburgense (base de l'Oxfordien inférieur ; zone à Mariae) et ses horizons ont été reconnus et étudiés avec précision dans presque tous les forages. Toutes les observations concernant la succession des horizons confirment sans équivoque la réalité d'un horizon à woodhamense venant au-dessus de l'horizon à scarburgense, lui-même au-dessus de l'horizon à thuouxensis.
L'espèce Cardioceras woodhamense Arkell, telle qu'elle a été figurée par son créateur, se caractérise par une carène perlée dès les tours internes, alors que cette morphologie n'existe pas chez l'holotype de Cardioceras scarburgense (Young et Bird). Dans les forages, C. woodhamense apparaît sans exception au-dessus des faunes de l'horizon à scarburgense. Cette succession « scarburgense–woodhamense » existe dans le Sud de l'Angleterre (Weymouth, Dorset) [10], le Boulonnais [57], le bassin du Sud-Est [29,30] et le Jura (travaux en cours).
Dans les forages, l'abondance des Cardioceratinae dans l'horizon à woodhamense a permis de reconnaître deux morphologies successives au sein de l'espèce C. woodhamense : une morphologie dite ancienne, dans les niveaux inférieurs de cet horizon, et une morphologie, dite récente, dans les niveaux supérieurs. Chez les plus anciennes, l'ontogenèse individuelle montre dans les tours les plus internes une morphologie typiquement « scarburgense » à carène non encore perlée, qui passe progressivement vers 15 à 20 mm de diamètre à la morphologie « woodhamense » à carène perlée ; au-delà de 40 mm, les côtes secondaires tendent à devenir ventralement proverses. Chez les plus récentes, la morphologie « woodhamense » apparaît plus tôt, vers 10 à 15 mm, et les côtes secondaires sont proverses précocement, vers 25 à 30 mm. Toutefois, la variabilité ontogénétique individuelle et le dimorphisme microconque–macroconque assez marqué chez ces ammonites doivent être pris en compte. Pour une application biochonologique, ces différences ne sont donc fiables que sur des populations d'effectif suffisant, ce qui n'est pas le cas de celles des forages. Les observations faites sur les spécimens de l'Est du bassin de Paris confirment cependant celles en cours sur des faunes jurassiennes, collectées en affleurement et nettement plus abondantes. Dans un avenir proche, il sera sans doute possible de mieux cerner les limites de ces deux entités taxonomiques à des fins d'utilisation biostratigraphique.
3.6 La succession horizon à praemartini–horizon à alphacordatum (Oxfordien inférieur ; zone à Mariae, sous-zone à Praecordatum)
La sous-zone à Praecordatum (zone à Mariae) est subdivisée en trois : horizon à praemartini, horizon à alphacordatum et horizon à praecordatum. L'horizon à praemartini [29] a pour espèce-indice Cardioceras praemartini Spath, dont la morphologie, par rapport à C. woodhamense, montre des côtes fines et ventralement proverses qui apparaissent très tôt dans l'ontogenèse, vers 20 mm de diamètre. Cette espèce est dite « évoluée », d'autant que son ornementation fine est associée à une carène ventrale en cours d'individualisation, vers 20 à 25 mm.
Au-dessus, l'horizon à alphacordatum [29] est fondé sur Cardioceras alphacordatum Spath, espèce où les stades ontogéniques précoces « scarburgense » et « woodhamense », ont disparu des tours internes et médians. Ils sont remplacés par un stade « praemartini » avant 15 mm de diamètre, tandis que chez certains individus des tubercules latéraux apparaissent clairement sur les côtes primaires ; corrélativement, les tours deviennent plus quadrangulaires avec des renflements latéroventraux. Ces caractéristiques morphologiques se renforcent dans l'horizon à praecordatum [44], espèce indice Cardioceras praecordatum Douvillé, qui annonce la morphologie des espèces indices de la biozone à Cordatum (Oxfordien inférieur).
Le premier stade (horizon à praemartini) de ce chronocline morphologique est bien illustré dans plusieurs forages : entre 418,83 et 411,17 m dans EST 312, entre 604,35 et 597,72 m dans EST 342 et entre 569,62 et 559,20 m dans MSE 101. Dans les autres, des intercalations plus carbonatées dans la série argileuse font que les ammonites deviennent plus rares, et les interprétations biochronologiques plus difficiles. L'horizon à alphacordatum n'est reconnu que dans EST 342 (entre 592,30 et 592,20 m). Dans HTM 102, la partie moyenne de la sous-zone à Praecordatum est entre 383,00 et 381,35 m, mais la faune peu caractéristique ne permet pas de préciser l'horizon. La sous-zone à Praecordatum existe dans EST 103 entre 461,69 et 446,80 m, mais sans possibilité d'en différencier les horizons. Quant à l'horizon à praecordatum, il n'est vraiment mis en évidence que dans EST 322, entre 518,59 et 517,62 m, donnant à cet intervalle valeur de transition avec la sous-zone à Bukowskii de la zone à Cordatum. La base de EST 204, entre 463,69 m et 450,56 m, n'a fourni que des ammonites peu caractéristiques, mais les quelques fragments collectés plaident en faveur de la présence de la sous-zone à Praecordatum. Les ammonites collectées dans les verses du puits auxiliaire, creusé à l'emplacement du forage EST 205, ont révélé la présence de l'horizon à praemartini à 484,71 m, 481,49 m et 478,46 m, ce qui est cohérent avec les observations issues du forage ; l'horizon à alphacordatum a été reconnu à 476,58 m et 474,24 m, et l'horizon à praecordatum à 464,87 m, complétant ainsi les données du forage. Le forage EST 104 n'a pas de faune plus jeune que l'horizon à woodhamense.
3.7 Difficultés de localisation de la limite Oxfordien inférieur–Oxfordien moyen
Des intercalations calcaires se développent vers le sommet de la série argileuse et les ammonites deviennent très rares et souvent peu caractéristiques. La reconnaissance des unités biostratigraphiques s'avère ainsi délicate au passage Oxfordien inférieur–Oxfordien moyen, placé entre la zone à Cordatum, sous-zone à Cordatum et la zone à Plicatilis, sous-zone à Vertebrale. Des intervalles d'incertitude s'étendent sur des épaisseurs sédimentaires plurimétriques à pluridécamétriques et la précision de la datation est le plus souvent de l'ordre du sous-étage ou de la zone, rarement de la sous-zone, jamais de l'horizon. De plus, dans la zone à Plicatilis, la faune est beaucoup plus homogène et banale. Elle est dominée par les Perisphinctidae, délicats à déterminer car souvent écrasés, et les groupes accompagnants sont rares et assez peu informatifs.
Un niveau de bentonite existe dans les forages HTM 102, EST 103, EST 104 [47] et dans le puits principal. Il a été retrouvé en affleurement à Saint-Blin [13] et dans le bassin du Sud-Est [46,48] où il est daté par ammonites du début de l'Oxfordien moyen, zone à Plicatilis, sous-zone à Vertebrale. Cet événement volcanique, instantané à l'échelle des temps géologiques, est donc un excellent élément de corrélation et de datation.
À noter enfin, qu'à défaut d'ammonites dans les forages HTM 102, MSE 101 et EST 103, les brachiopodes et les associations palynologiques permettent de proposer un positionnement de la limite Oxfordien inférieur–Oxfordien moyen.
La zone à Cordatum est difficile à reconnaître et les sous-zones qui la composent sont inégalement mises en évidence, laissant soupçonner des condensations, voire des lacunes. Elle n'est certaine que dans EST 322 et dans le puits auxiliaire (EST 205). L'unité inférieure, sous-zone à Bukowskii, n'est reconnue que dans EST 322 et la supérieure, sous-zone à Cordatum, que dans EST 322, le puits principal (EST 204) et le puits auxiliaire (EST 205). Quant à l'unité moyenne, la sous-zone à Costicardia, elle n'a jamais été mise en évidence. Cette dernière, reconnue en Angleterre (Yorkshire et Dorset) [1], probablement en Pologne [39], n'a été prouvée en France que dans le Jura (travaux en cours). Dans le bassin du Sud-Est, sa présence est probable, mais elle n'a pas encore été prouvée par ammonites. D'après leurs caractéristiques paléonvironnementales, ces régions du Sud-Est sont sans doute assez profondes ou plus ouvertes, alors que l'Est du bassin de Paris est sans doute moins profond ou moins ouvert. Généralement, quand cette sous-zone est mise en évidence, elle correspond à une sédimentation réduite et peu fossilifère qui caractériserait un épisode de bas niveau marin relatif sur une bordure de plate-forme distale.
La zone à Plicatilis est présente dans EST 103 et EST 322, ainsi que dans les puits principal et auxiliaire. Dans tous les cas, seule la sous-zone à Vertebrale a été mise en évidence (EST 322 et puits auxiliaire-EST 205). Au-dessus, les faciès deviennent de plus en plus carbonatés et dépourvus, ou presque, d'ammonites : la sous-zone à Antecedens n'a pas été reconnue.
La limite Oxfordien inférieur–Oxfordien moyen peut être positionnée entre 375,58 m et 368,30 m dans HTM 102, cotes auxquelles ont été trouvées quelques ammonites peu caractéristiques mais généralement présentes dans ces deux sous-étages. Elle est en accord avec les dernières ammonites certaines de l'Oxfordien inférieur, zone à Mariae, sous-zone à Praecordatum à 381,35 m et les premières ammonites certaines de l'Oxfordien moyen à 355,90 m. Les associations palynologiques à 371 m et 352 m de la transition Oxfordien inférieur–Oxfordien moyen (Jan du Chêne, données inédites), et la bentonite à 351,42 m sont aussi en accord avec ce positionnement.
Entre 516,80 m et 516,00 m dans EST 322, un intervalle avec une association d'ammonites de la zone à Cordatum, sous-zone à Cordatum et de la zone à Plicatilis, sous-zone à Vertebrale assure le passage entre Oxfordien inférieur et moyen ; ce dernier est reconnu jusqu'à la cote 510,04 m.
Dans EST 342, l'Oxfordien moyen débuterait à la cote 590,10 m, immédiatement au-dessus des dernières ammonites de la zone à Mariae. L'horizon à praecordatum et la zone à Cordatum n'ont pas été reconnus, mais il est impossible de conclure à leur absence, compte tenu des faunes trop peu caractéristiques au-dessus de l'horizon à alphacordatum, présent entre 592,30 m et 592,20 m.
Un ensemble d'arguments concordants conduit à placer la limite Oxfordien inférieur–Oxfordien moyen au-dessus de 456,12–456,19 m dans EST 103, dernières ammonites de l'Oxfordien inférieur (zone à Mariae, sous-zone à Praecordatum). Les premières ammonites de l'Oxfordien moyen (zone à Plicatilis) sont nettement plus haut dans le forage, à 433,74–433,85 m, et la bentonite est entre 429,30 et 429,40 m. Les données de EST 104, proche de EST 103, sont compatibles. Les dernières ammonites de l'Oxfordien inférieur (zone à Mariae, sous-zone à Scarburgense, horizon à woodhamense) sont à 466,52 m. Elles sont suivies d'un épais intervalle sans faune caractéristique et la bentonite est entre 428,97 et 429,05 m.
Dans MSE 101, il n'y a aucune ammonite caractéristique entre 559,20 m, derniers indices de l'Oxfordien inférieur (zone à Mariae, sous-zone à Praecordatum, horizon à praemartini) et 432,00 m. La limite pourrait se situer entre 523 m et 518 m, fondée sur la présence de brachiopodes de l'Oxfordien moyen (Thurmanella obtrita, Gallienithyris cf. maltonensis) dans cet intervalle (Garcia, données inédites), et d'associations palynologiques caractéristiques de la transition Oxfordien inférieur–Oxfordien moyen vers 535 m (Jan du Chêne ; données inédites). La bentonite n'a pas été retrouvée dans ce forage.
Des ammonites de l'Oxfordien moyen, zone à Plicatilis ont été récoltées dans le puits principal (EST 204) entre 431,47 et 413,03 m ; la bentonite est à 427,00 m. La limite Oxfordien inférieur–Oxfordien moyen serait donc comprise entre cette dernière cote et les ammonites de la zone à Mariae, sous-zone à Praecordatum (463,69 à 450,56 m). Dans le puits auxiliaire (EST 205), quelques brachiopodes à 442,87 m seraient d'âge Oxfordien moyen, zone à Plicatilis, sous-zone à Vertebrale ; ils viennent peu au-dessus (445,77 m) d'une ammonite très probablement de l'Oxfordien inférieur, zone à Cordatum, sous-zone à Cordatum, elle-même au-dessus d'ammonites de la zone à Mariae, sous-zone à Praecordatum, horizon à praecordatum, entre 470,41 et 468,05 m.
À la cote 405,40 m, les échantillons d'ammonites les plus jeunes du forage EST 312 donnent un âge Oxfordien inférieur. L'Oxfordien moyen n'est pas caractérisé.
3.8 Corrélations entre les forages et enregistrement sédimentaire différentiel des unités biostratigraphiques
La précision de datation obtenue dans les forages permet fréquemment des corrélations à la résolution de l'horizon biostratigraphique, le plus souvent à celle de la sous-zone et pratiquement toujours à celle de la zone. Deux transects sont établis :
- – le premier, orienté WSW–ENE concerne EST 342, EST 322, HTM 102 et EST 312 (Fig. 2) ;
- – le second, orienté NNE–SSE, relie MSE 101, EST 103, EST 104, EST 204, EST 205 et HTM 102 (Fig. 3). La limite Callovien–Oxfordien, bien repérée dans la plupart des forages, est choisie comme référence horizontale.
Considérant que les conditions de sédimentation et l'évolution post-sédimentaire de la série argileuse callovo-oxfordienne semblent avoir été relativement homogènes pour tout le secteur considéré, il est possible de relever et de comparer les épaisseurs de chaque unité biochronostratigraphique. Cette procédure semble justifiée par le fait que les phénomènes de compaction ont vraisemblablement été similaires sur l'ensemble du secteur : couverture post-callovo-oxfordienne de même nature et d'épaisseurs voisines, profondeur actuelle du complexe argileux de même ordre de grandeur, fossiles écrasés de manière identique par compression.
3.8.1 Le Callovien moyen
Son épaisseur la plus courante est comprise entre 25 et 30 m, avec un faible gradient d'augmentation OSO–ENE depuis EST 342, vers HTM 102, ainsi qu'en direction du nord, vers MSE 101. En revanche, une forte augmentation à 45 m est enregistrée dans EST 312, forage le plus oriental.
Des deux biozones du Callovien moyen, la zone à Jason est très réduite. Dans les forages où elle a été reconnue, elle n'intéresse que quelques décimètres de sédiments ; au plus, elle pourrait atteindre 2 m dans EST 322. Dans tous les cas, seule la sous-zone à Medea est reconnue.
La zone à Coronatum est nettement plus épaisse ; elle atteint plusieurs dizaines de mètres, avec une nette augmentation d'ouest en est. De 22 m dans EST 342, elle atteint 40–45 m dans EST 312 en passant par des épaisseurs comprises entre 26–34 m à EST 322 et 29–31 m à HTM 102 au plus près du site de Bure. Dans MSE 101, forage le plus septentrional, une épaisseur minimum de 30 m est cohérente avec les autres forages ; cependant, à cause d'incertitudes de datation engendrées par des niveaux sans ammonites caractéristiques, elle pourrait atteindre un maximum de 43 m, épaisseur tout aussi cohérente avec celle observée dans EST 312, forage le plus oriental.
Les épaisseurs et faciès du Callovien moyen des forages diffèrent fortement de celles des affleurements plus au SSO (Latrecey, Bologne-Marault, Humberville et Saint-Blin/Semilly) ; très fossilifères quand elles existent, la zone à Jason et la zone à Coronatum sont réduites à quelques mètres ou décimètres de calcaires argileux ou de marno-calcaires à oolithes ferrugineuses [13,19,20,36].
3.8.2 Le Callovien supérieur
Le Callovien supérieur présente à la fois une forte réduction d'épaisseur et un faible gradient de variation d'épaisseur ouest–est ; des lacunes et des phénomènes de concentration faunique traduisent sans doute une réduction de l'accumulation sédimentaire. Les épaisseurs enregistrées vont de 3 m dans EST 342 à 12 m dans EST 312. Des variations non significatives existent dans le sens nord–sud : 7 m estimés dans MSE 101, 4 et 5 m dans EST 103 et EST 104, 7 m dans HTM 102.
3.8.2.1 La zone à Athleta.
Reconnue dans EST 312 et avec doute dans HTM 102, la zone à Athleta est réduite à quelques alternances décimétriques de marnes et de bancs un peu plus carbonatés au sein de la série argileuse. L'absence de cette biozone dans les autres forages correspond peut-être à une lacune réelle, mais sans doute aussi à la rareté ou à l'absence de faune caractéristique. Quand cette biozone existe, sa stratonomie d'alternances et son faciès marnocalcaire rappellent les calcaires et marnes feuilletées observées en affleurement (Bologne-Marault) [17]. Ailleurs (Humberville, Saint-Blin/Semilly, Latrecey), cette biozone est souvent incluse dans des dépôts condensés à oolithes ferrugineuses [13,20,36].
3.8.2.2 La zone à Lamberti.
Malgré une épaisseur assez faible, la zone à Lamberti présente un gradient d'augmentation ouest–est : de 3 m dans EST 342, elle atteint 12 m dans EST 312. Des épaisseurs intermédiaires sont relevées : 4 m dans EST 322, 5 m dans EST 103, entre 4 et 6 m dans EST 104 et 7 m dans HTM 102 ; par absence d'ammonites caractéristiques, aucune évaluation n'est possible pour les forages MSE 101 et EST 205. Il semble que ce gradient soit associé à une conservation de plus en plus complète des sous-zones et horizons qui subdivisent la zone à Lamberti. À l'ouest, la sous-zone à Henrici n'est pas reconnue dans EST 342 et HTM 102, mais la sous-zone à Lamberti est présente avec ses trois horizons ; à l'est, ses deux sous-zones et ses cinq horizons sont reconnus dans EST 312. Au sein de la zone à Lamberti, la sous-zone à Henrici, quand elle est présente, est toujours réduite à quelques décimètres, alors que la sous-zone à Lamberti est toujours plus épaisse. Dans cette dernière, le développement des trois horizons qui la composent est très variable et ne semble pas suivre de logique : l'horizon à praelamberti est absent dans MSE 101, EST 103 et EST 104 ; l'horizon à lamberti et l'horizon à paucicostatum ont une fréquence de présence plus élevée et leurs épaisseurs cumulées sont souvent complémentaires.
Les alternances de calcaires argileux et marnes, ainsi que les épaisseurs relevées en forage sont compatibles avec les affleurements les plus proches (Humberville, Saint-Blin/Semilly), où la zone à Lamberti, sans possibilité de subdivision, atteint 6 m de développement [12,36]. Plus au sud-ouest, à Latrecey, cette biozone est sous forme de dépôts réduits à oolithes ferrugineuses [17].
3.8.3 L'Oxfordien inférieur
Ce sous-étage présente les taux de sédimentation les plus élevés de toute la série, avec des variations d'épaisseur très significatives et un enregistrement fortement différentiel entre ses diverses unités biostratigraphiques. À titre d'exemple, le forage EST 322 souligne cette grande disparité avec une zone à Mariae dépassant 50 m et une zone à Cordatum comprise entre 1 et 2 m, où la sous-zone à Bukowskii (entre 518,69 et 517,82 m) et la sous-zone à Cordatum (entre 517,62 et 516,80 m) sont seules présentes. De nouveau, un net gradient d'augmentation d'épaisseur est observé d'ouest en est : 26 m dans EST 342, 55 m dans EST 322 et 52 m dans HTM 102. La limite Oxfordien inférieur–Oxfordien moyen n'a pas été reconnue dans EST 312, où toutefois au moins 42 m de sédiments concernent sans doute ce sous-étage.
La zone à Mariae est très bien documentée ; la zone à Cordatum l'est beaucoup moins, soit par rareté ou absence d'ammonites caractéristiques, soit par absence de carottage. Ce manque d'information, en particulier à la limite entre les deux zones du sous-étage entraîne inévitablement des incertitudes sur les épaisseurs de sédiments enregistrés dans les intervalles correspondants.
L'Oxfordien inférieur argileux des forages contraste fortement avec celui des affleurements. Plus au sud-ouest, à Latrecey [17] la totalité du sous-étage est incluse dans une série condensée de marno-calcaires à oolithes ferrugineuses de quelques décimètres d'épaisseur. En revanche, il est faciologiquement assez semblable à celui de l'affleurement de Saint-Blin/Semilly [12], où il atteint 33 m ; toutefois, le développement respectif des unités biostratigraphiques est totalement différent.
3.8.3.1 La zone à Mariae.
Son épaisseur passe de 25 m dans EST 342 à 53–55 m dans EST 322, puis à 52–58 m dans HTM 102 ; cette dernière valeur est compatible avec l'ensemble des observations, malgré des incertitudes sur le positionnement de ses limites dans ce dernier forage. Le maximum d'épaisseur serait enregistré sur le site de Bure (EST 103), où l'extension de cette biozone est de 67 m. Un manque de données ne permet pas d'évaluer les épaisseurs dans EST 312, MSE 101, EST 104 et EST 205. La seule sous-zone à Scarburgense, passe de 11 m d'épaisseur dans EST 342 et EST 322, à 30 m dans HTM 102, puis 36 m dans EST 312. Elle atteint 52 m dans EST 103 et 47 m dans EST 104, confirmant le fort taux de sédimentation de cet intervalle au droit du site de Bure. Dans MSE 101, malgré des incertitudes, la sous-zone à Scarburgense atteint au moins 30 m d'épaisseur.
Dans l'affleurement de Saint-Blin/Semilly [12], la zone à Mariae se développe sur 16 m, donc sensiblement moins que dans les forages ; en son sein, la sous-zone à Scarburgense atteint 11,5 m et la sous-zone à Praecordatum 4,5 m, ce qui est proportionnellement compatible avec les forages. L'horizon à scarburgense (5,5 m) et l'horizon à woodhamense (6 m) ont une extension quasi identique ; l'horizon basal à thuouxensis n'a pas été reconnu et au-dessus, seul l'horizon à praecordatum (4,5 m) a été mis en évidence.
3.8.3.1.1 3.8.3.1.1. La sous-zone à Scarburgense.
L'horizon basal à thuouxensis est généralement très réduit (0,10 à 1 m dans EST 103 et EST 104) ; il n'a pas été reconnu dans tous les autres forages, mais cette absence n'indique peut-être qu'un manque de données fauniques, et peut-être pas une lacune.
L'horizon à scarburgense, présent dans tous les forages, est toujours épais ; malgré quelques incertitudes pour cerner sa limite supérieure, il varie de 3 m (EST 342) à près de 20 m (EST 103), en passant par des valeurs intermédiaires : 5 m dans HTM 102, 5 à 10 m dans EST 322, 9 à 12 m dans EST 312, 8 à 13 m dans MSE 101, 5 à 16 m dans EST 205.
L'horizon à woodhamense est toujours l'unité la plus développée ; quel que soit le forage, il représente plus des deux tiers de l'amplitude sédimentaire de la sous-zone à Scarburgense. La distribution des épaisseurs suit toujours un gradient d'augmentation globalement ouest–est : 8 à 10 m dans EST 342 et EST 322, 22 à 25 m dans HTM 102 et EST 312. Il atteint 20 m dans MSE 101 et EST 205. Dans EST 103, l'horizon à woodhamense n'est formellement reconnu que sur 8 m ; cependant, considérant les cotes respectives de la limite supérieure de la sous-zone à Scarburgense et de la base de l'horizon à scarburgense, son épaisseur réelle serait comprise entre 32 et 52 m, valeurs compatibles avec les données du forage EST 104.
3.8.3.1.2 3.8.3.1.2. La sous-zone à Praecordatum.
Compte tenu des incertitudes concernant sa délimitation et la reconnaissance de ses subdivisions, l'évaluation en épaisseur de sédiments de la sous-zone à Praecordatum est impossible. Cependant, le seul horizon à praemartini, unité la plus fréquemment reconnue, enregistre des taux de sédimentation très élevés, du même ordre de grandeur que ceux des horizons de la sous-zone à Scarburgense. Il atteint au moins 8 m dans EST 312, 10 m dans MSE 101, 13 m dans EST 342 et 15 m dans EST 103, confirmant le développement important de la série argileuse pour le même intervalle de temps, au droit du site de Bure. Malgré un manque de données, et compte tenu du positionnement de la limite zone à Mariae-zone à Cordatum dans EST 322 et HTM 102, cet intervalle est sans doute bien développé dans ces deux derniers forages.
3.8.3.2 La zone à Cordatum.
D'une manière générale, la zone à Cordatum est toujours nettement réduite, avec des épaisseurs qui ne peuvent être évaluées que dans deux forages : quelques décimètres dans EST 322 à quelques mètres dans HTM 102. Les sous-zones qui la composent sont inégalement mises en évidence, laissant soupçonner des condensations, voire des lacunes.
À Saint-Blin/Semilly [12], la zone à Cordatum est beaucoup mieux documentée et montre de fortes différences d'enregistrement vis-à-vis des forages. Elle se développe sur 17 m et ses trois sous-zones sont épaisses et bien caractérisées : sous-zone à Bukowskii, 4 m ; sous-zone à Costicardia, 3 m, sous-zone à Cordatum, 11 m.
3.8.4 L'Oxfordien moyen
Trop incomplètes, aussi bien dans les forages que dans les affleurements, les données sur l'Oxfordien moyen ne permettent pas d'évaluation de l'enregistrement sédimentaire de ce sous-étage, a fortiori de ses subdivisions, dont seule l'unité basale, la sous-zone à Vertebrale de la zone à Plicatilis, a été ponctuellement reconnue.
4 Analyse des peuplements ammonitiques : intérêts paléoécologique et paléobiogéographique ; applications paléoenvironnementale et paléogéographique
4.1 Taphonomie et préservation des coquilles
Les sédiments argileux renfermant les ammonites des forages sont riches en matière organique et le plus souvent fortement bioturbés. La pyrite y est diffuse et les ammonites elles-mêmes ont des tours internes parfois pyritisés. Les coquilles sont généralement de petite taille, mais assez rarement fragmentées. Le plus souvent, ce sont des répliques et des moules internes. Elles sont presque toujours latéralement écrasées, rendant difficile l'observation de la forme de la section des tours ; les lignes de sutures du phragmocône ne sont que rarement visibles.
Les apophyses jugales des Hecticoceratinae, Perisphinctidae, Peltoceratinae et Kosmoceratidae, ou le rostre ventral des Cardioceratidae, qui caractérisent le péristome des microconques adultes, n'ont été que peu retrouvés ; elles n'ont été observées que chez quelques individus des genres Creniceras, Kosmoceras et Orionoides. Étant donné qu'il n'existe pas d'indices d'hydrodynamisme important (rides de vagues et de courants ou figures de tempête), on peut conclure à des dépôts effectués dans un environnement calme et suffisamment profond (), qui correspond à l'offshore inférieur dans la zonation des milieux de sédimentation. L'absence presque généralisée d'apophyses ou de rostres peut alors s'expliquer de trois façons :
- – les microconques retrouvés ne sont jamais adultes, ce qui est statistiquement peu probable, vu le nombre élevé de spécimens récoltés ;
- – ces structures péristoméales étaient peu calcifiées, ce qui aurait facilité leur dissolution dans un milieu géochimiquement assez agressif ;
- – ou enfin, parce que les fossiles des forages sont des répliques ou des moules internes.
4.2 Composition des peuplements : analyse paléoécologique et applications paléoenvironnementales
Sur l'ensemble des forages, plusieurs centaines d'ammonites ont été récoltées ; considérant les trois sous-ordres d'Ammonoidea jurassiques, ce sont à 99% des Ammonitina. Les Phylloceratina représentent le 1% restant ; aucun Lytoceratina n'a été trouvé. Cette composition est certainement le reflet de modes de vie différents de ces trois groupes, ainsi que de certains facteurs paléoenvironnementaux de la bordure est du bassin de Paris au Callovien–Oxfordien.
4.2.1 Les Phylloceratina et Lytoceratina
Au sein des Phylloceratina, le genre Sowerbyceras est totalement absent. Ce constat est important car, en dehors du bassin du Sud-Est et de sa bordure ardéchoise où il abonde et représente, dans certains niveaux du Callovien–Oxfordien, jusqu'à 80% de la faune ammonitique [21,28–30], le genre Sowerbyceras est toujours rare. Il en est de même dans le Jura (travaux en cours) sauf dans sa partie méridionale. Deux explications sont possibles :
- – ce genre est exclusivement téthysien et son extension vers le nord est limitée par des conditions environnementales exclusives aux marges et talus de cette aire océanique ;
- – son absence sur des plates-formes ou dans des bassins intracratoniques comme le bassin de Paris est associée à son mode de vie.
Au sein des Phylloceratina, le genre Sowerbyceras a une morphologie très originale. Sa section est plutôt quadrangulaire, avec des flancs subparallèles, un ventre arrondi et un ombilic largement ouvert ; sa coquille est donc de type platycône. Ses lignes de suture, relativement simples, contrastent avec celles généralement très découpées des autres Phylloceratina. Morphologiquement, les Sowerbyceras se rapprochent donc des Ammonitina platycônes et ils avaient vraisemblablement un mode de vie nectobenthique en domaine de plate-forme distale. Ils n'étaient probablement pas nectopélagiques comme les autres Phylloceratina à coquille sphaerocône à subsphaerocône, plus globuleuse, à ombilic très étroit et sutures très complexes [25,50,56]. Après la mort, vis-à-vis du milieu aquatique, leurs coquilles se comportaient de manière différente de celles des autres Phylloceratina. Celles des Sowerbyceras, au mode de vie plutôt nectobenthique, ne devaient pas pouvoir, ou que très rarement, remonter vers la surface, ce qui réduit, voire exclut une flottaison post-mortem ; leur dispersion vers les faibles profondeurs des plates-formes plus proximales était donc faible, voire impossible. À l'opposé, celles des autres Phylloceratina, au mode de vie plutôt nectopélagique et qui pouvaient mourir entre deux eaux, plus près de la surface, ont eu une dispersion post-mortem un peu plus fréquente et plus vaste.
4.2.2 Les Ammonitina
Deux approches permettent d'utiliser les ammonites pour des reconstitutions paléoenvironnementales : une approche synécologique (analyse des peuplements) et une approche autécologique (analyse des caractères morphologiques en relation avec le milieu). Complémentaires, elles permettent de travailler simultanément selon ces deux concepts. Une question se pose alors : au sein d'une population, les peuplements ammonitiques, c'est-à-dire les pourcentages relatifs des différents genres et/ou espèces (approche synécologique), et par voie de conséquence les pourcentages des différentes morphologies (approche autécologique), sont-ils de bons indicateurs de milieu ?
Les résultats obtenus montrent que les compositions des peuplements ammonitiques ne sont pas aléatoires et s'accordent bien avec les paléoenvironnements déduits des analyses sédimentologiques [22–24,33–35,38,41]. Ainsi, lorsque les paléoprofondeurs sont fortes, ce sont surtout les ammonites serpenticônes et sphaerocônes à cloisons complexes qui abondent ; cela suppose pour ces formes un mode de vie nectopélagique [55]. En revanche, lorsque la paléoprofondeur diminue, on constate que les morphologies platycônes et suboxycônes à cloisons plus simples dominent ; cela suggère alors, pour ces formes, un mode de vie nectobenthique. Dans ce deuxième cas, on ne trouve jamais de Phylloceratina associés, sauf de façon très anecdotique.
Le trop petit nombre de spécimens collectés dans le Callovien moyen des forages (zone à Jason et zone à Coronatum) et la biodiversité restreinte qui en découle, ne permettent pas d'interprétations paléoécologiques ou paléoenvironnementales. Cependant, l'association de Kosmoceratidae, Pseudoperisphinctidae, Hecticoceratidae et rares Pachyceratidae est tout à fait conforme à ce qui est observé dans les affleurements [50].
Au Callovien supérieur, dans les affleurements des bordures du bassin de Paris, les Ammonitina sont relativement diversifiées. À côté des Hecticoceratinae et des Cardioceratinae qui sont majoritaires, les Distichoceratinae, les Kosmoceratinae et les Pseudoperisphinctinae sont aussi présents ; il peut s'y ajouter quelques genres très minoritaires tels que Taramelliceras et Berniceras. Dans les forages, surtout dans la zone à Lamberti, les peuplements sont tout à fait similaires, à l'exclusion des formes rares qui sont proportionnellement trop peu nombreuses pour être représentées.
À la base de l'Oxfordien inférieur, zone à Mariae, sous-zone à Scarburgense, horizon à scarburgense, les Peltoceratinae sont assez bien représentés et parfois même abondants dans une grande partie de l'Europe occidentale ; cette particularité s'observe dans les forages. Au-dessus, dans l'horizon à woodhamense, les Hecticoceratinae et des Cardioceratidae redeviennent dominants, alors que les Perisphinctidae sont toujours moins bien représentés, aussi bien dans les forages qu'à l'affleurement. Dans ces derniers, les Taramelliceratinae sont parfois présents, mais plutôt anecdotiques [30] ; en revanche, on les trouve plus souvent dans la partie moyenne de cet horizon, comme dans le Boulonnais [57] ou le Jura (travaux en cours). Cette caractéristique se retrouve dans les deux forages EST 312 et EST 322 à l'ouest du site de Bure, où existent non seulement des Taramelliceratinae, mais aussi le genre Creniceras, en général plus fréquent dans les régions plus méridionales, sans doute un peu plus ouvertes et plus profondes comme le Jura.
Dès la base de la sous-zone à Praecordatum, dans l'horizon à praemartini, surtout bien reconnu dans les verses des puits, on observe une baisse de l'abondance en spécimens et de la biodiversité. Les Cardioceratinae sont cependant de nouveau assez bien représentés, mais ce sont les Perisphinctidae qui dominent ; les Taramelliceratinae sont redevenus rares. Au-dessus, dans l'horizon à alphacordatum et l'horizon à praecordatum, la rareté des ammonites s'accentue et ne permet pas une approche synécologique, de même que pour la zone à Cordatum où ne coexistent que quelques Perisphinctidae et Cardioceratinae.
À la base de l'Oxfordien moyen, zone à Plicatilis, sous-zone à Vertebrale, comme presque partout en Europe occidentale, la faune est très nettement enrichie en Perisphinctidae. Généralement, cet enrichissement s'accentue encore dans la sous-zone à Antecedens sus-jacente, mais cette dernière n'est pas clairement identifiée dans les forages.
En appliquant aux peuplements ammonitiques des forages et des affleurements de l'Est du bassin de Paris, l'analyse synécologique désormais utilisée [35], la faune apparaît franchement dominée par des ammonites nectobenthiques, caractérisant un environnement ouvert mais de profondeur moyenne ne dépassant sans doute pas 150 m. Les petites différences de composition faunique observées, tel qu'un enrichissement en Taramelliceratinae entre, d'une part, les forages du site de Bure et ceux situés à l'ouest de ce dernier et, d'autre part, ceux localisés plus vers l'est, indiquent un milieu sans doute un peu moins profond dans cette direction.
4.3 Composition des peuplements : analyse paléobiogéographique et applications paléogéographiques
Les peuplements d'ammonites observés dans les forages illustrent parfaitement les deux événements paléobiogéographiques qui perturbent l'équilibre dynamique existant depuis le Bajocien–Bathonien entre les faunes boréales et téthysiennes : (1) l'expansion vers le sud des Kosmoceratinae à la base du Callovien moyen, suivie (2) au sommet du Callovien supérieur et à l'Oxfordien inférieur, par l'expansion des Cardioceratinae. Il se crée alors une aire biogéographique hétérogène, considérée comme un domaine d'interface à la rencontre de deux biotas [25]. En conséquence, sur l'Europe occidentale, et en particulier sur le bassin de Paris, cette interface correspond à ce qui est souvent considéré comme des provinces : « province Subboréale » et « province Subméditerranéenne ». L'individualisation de ces dernières est fondée sur l'importance relative, au sein des peuplements, de taxons septentrionaux (Cardioceratinae et Kosmoceratinae), méridionaux (Hecticoceratinae, Pachyceratidae, Peltoceratinae), ou plus ubiquistes (Perisphinctidae) [2,50]. Elles comportent des faunes mixtes et occupent des secteurs plus ou moins vastes sur les plates-formes intracratoniques nord-péritéthysiennes ; en fonction de cette composition variable, leurs limites et extensions sont fluctuantes au cours du temps.
Au Callovien moyen, les Kosmoceratinae, bien implantés au Callovien inférieur sur l'Angleterre et la bordure nord du bassin de Paris, s'étendent vers le sud dès la zone à Jason ; cet épisode place le bassin de Paris et le Jura sous influence subboréale. Ensuite, dans la zone à Coronatum, les Pachyceratidae plutôt téthysiens dominent et placent cette région sous influence subméditerranéenne. Ces événements sont bien documentés dans EST 342, EST 322, HTM 102, EST 312 et MSE 101 où Kosmoceratinae, puis Pachyceratidae sont accompagnés d'Hecticoceratinae, Perisphinctidae et de rares Reineckeidae. Ces dernières, plus méridionales, voire ubiquistes, sont proportionnellement plus abondantes dans la zone à Coronatum que dans la zone à Jason.
Avec la base du Callovien supérieur, zone à Athleta, les formes méridionales et ubiquistes restent fortement majoritaires face aux formes septentrionales, gardant ainsi un caractère subméditerranéen assez marqué. Un important renouvellement faunique marque la zone à Lamberti qui est le témoin d'une invasion massive des Cardioceratinae boréaux, accompagnés des derniers Kosmoceratinae [5,18,50,52]. Le médiocre enregistrement de la zone à Athleta ne permet pas de vérifier cet intermède dans l'expansion des groupes boréaux. En revanche, l'abondance et la diversité des faunes de la zone à Lamberti dans tous les forages placent indiscutablement l'Est du bassin de Paris dans un contexte subboréal.
À l'Oxfordien inférieur, un gradient de composition faunique reste marqué depuis l'Écosse (île de Skye) jusqu'au bassin du Sud-Est [9], avec des influences boréales au nord et des influences méridionales au sud ; l'Est et le Sud-Est du bassin de Paris restent cependant assez marqués par les influences boréales, avec la présence constante et importante des Cardioceratinae. Dans les forages, les associations d'ammonites ont plus d'affinités avec les faunes du Boulonnais [57] qu'avec celles du Sud et du Sud-Est du bassin de Paris [22]. Par rapport au Jura, les différences sont plus marquées, dans la mesure où il existe, dans cette région, des genres micromorphes inconnus dans le bassin de Paris ou sur ses bordures [42]. Enfin, par rapport au bassin du Sud-Est [41], la différence majeure réside pour ce dernier dans la fréquence, voire l'abondance, des Phylloceratina représentés surtout par le genre Sowerbyceras. Mais, en ce qui concerne les Ammonitina, elles présentent une biodiversité identique à celles du bassin de Paris, y compris en proportions, Phylloceratina exclus. Cette dernière observation, ajoutée au fort taux de sédimentation qui le caractérise, montre bien que le bassin du Sud-Est était un secteur nettement plus subsident que le bassin de Paris, mais où la tranche d'eau n'était pas vraiment beaucoup plus importante. Ceci est confirmé par la mise en évidence de tempestites distales dans certains niveaux des « Terres noires » du bassin du Sud-Est, alors que ces figures sédimentaires n'ont pas été observées dans les forages.
À l'Oxfordien moyen, dans les sondages, comme partout en Europe occidentale, les Cardioceratinae sont de plus en plus rares, alors que les Perisphinctidae deviennent de plus en plus fréquents. Les quelques spécimens récoltés appartiennent effectivement à cette famille, mais la raréfaction des ammonites ne permet pas d'interprétation paléobiogéographique fondée.
5 Conclusion
Les faunes d'ammonites des forages de l'Andra confirment que la succession biostratigraphique du Callovien moyen–supérieur et de l'Oxfordien inférieur–moyen établie pour l'Europe occidentale est fiable et robuste. En fonction de la qualité de préservation et l'abondance des spécimens, tant dans les forages que dans les affleurements de la moitié est du bassin de Paris, la succession des diverses unités biostratigraphiques est vérifiée. La précision de datation est toujours atteinte à hauteur de la zone, fréquemment à celle de la sous-zone, ainsi qu'assez souvent à celle de l'horizon faunique, en particulier au Callovien supérieur et à l'Oxfordien inférieur.
Les peuplements d'ammonites vont dans le sens des résultats de la géologie sédimentaire et de la géodynamique du bassin de Paris. Elles montrent que le secteur le plus occidental concerné par cette étude, en direction du centre du « bassin », apparaît comme celui où la tranche d'eau devait être la plus forte, mais où la sédimentation s'est révélée la moins épaisse du Callovien moyen à la base de l'Oxfordien moyen. Les épaisseurs enregistrées pour cet intervalle stratigraphique témoignent cependant d'une subsidence généralisée, nettement plus forte et plus continue que sur les bordures, en particulier en direction du sud-est. Cette caractéristique semble être une constante depuis le début du Mésozoïque.
Les associations d'ammonites s'intègrent parfaitement dans le contexte paléobiogéographique des plates-formes et bassins intracratoniques nord-péritéthysiens où des influences boréales sont surtout bien marquées à partir du Callovien supérieur, jusqu'au passage Oxfordien inférieur–Oxfordien moyen. Elles confirment la réalité d'aires intermédiaires, dites provinces « subboréale » et « subméditerranéenne », à faunes de composition mixte et d'extension fluctuante dans l'espace et dans le temps, jouant le rôle d'interface entre les provinces « boréale » et « téthysienne ». Elles confirment aussi les liaisons directes existant à cette époque entre les vastes étendues des plates-formes marines péritéthysiennes peu profondes de l'Europe occidentale et, d'une part les mers boréales, et d'autre part la façade océanique téthysienne toutes deux plus profondes. Cependant, les influences méridionales et/ou océaniques, sont toujours moins fortes que les influences septentrionales ; la preuve en est l'absence totale ou la grande rareté de certains taxons, pourtant relativement abondants dans le Sud-Est de la France.