

Abridged English version
1 Introduction
The major shell bed concept constitutes an original approach for correlation at the basin scale. This concept has been initially proposed for the Miocene shell deposits of Maryland [18–20] and since then has been applied to various geographic and stratigraphic examples [11,12,26].
Two kinds of such conserved levels can be recognized. In the first type, the shell beds are linked to high-frequency cycles (5th- to 7th-order sequences) of the fluctuations of the sea level. In the second type, more largely distributed, the major shell beds are situated at the base of the transgressive interval of 3rd-order sequences. In the Paris Basin, the Bathonian brachiopod major beds are the result of the complex interactions between the shell accumulation, the eco-taphonomic features (taphonomic feedback, time averaging), and the shifting of the benthos closely linked to marine fluctuations. The so-called coralligène [25] (FBV) of the Saoura (Fig. 1) furrow (western Algerian Sahara) must be related to this kind of major shell bed. It is dated to the Upper Emsian and can be correlated with similar levels of the Ahnet-Mouydir area and of the Tindouf and Zemmour basins (western Sahara).
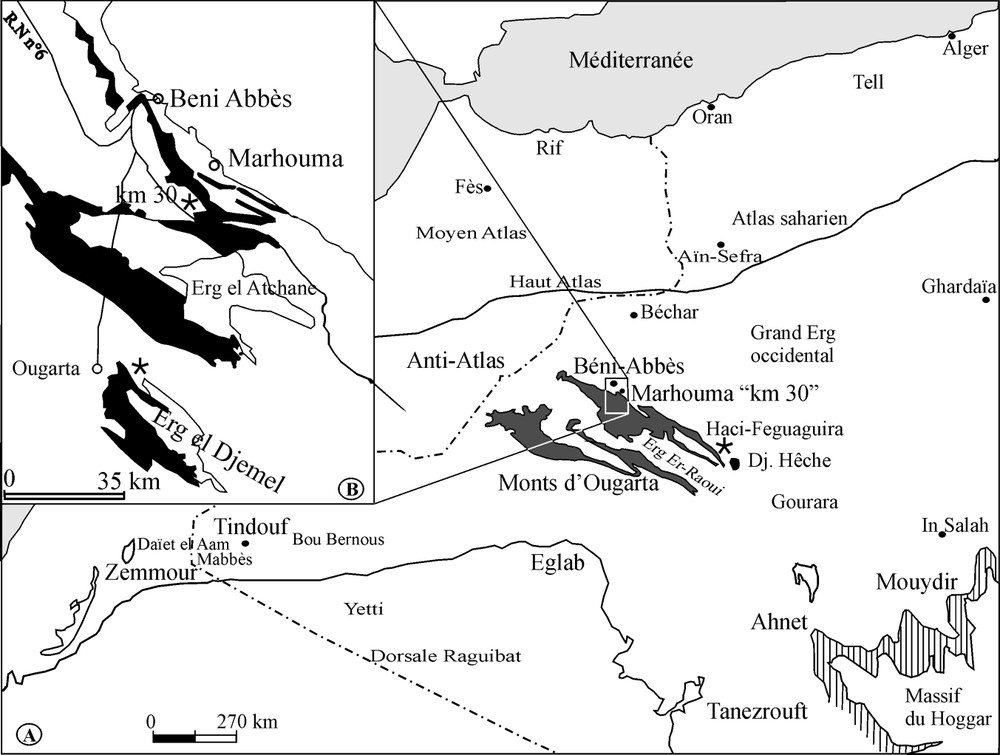
Carte de situation des secteurs étudiés : (A) localisation de la chaîne d'Ougarta (en gris foncé), du Paléozoïque du Tassili (en hachures) et des gisements étudiés (étoiles) ; (B) affleurements dévoniens des environs de Béni-Abbès (en noir).
Sketch location map of the studied areas: (A) Ougarta range (dark grey); Palaeozoic of the Tassili (hachures); (B) Devonian outcrop near Beni-Abbès (in black).
2 Saoura FBV: physical features and environmental significance
2.1 Stratigraphic position
The Saoura FBV is located at the base of the Cheffar el Ahmar formation, which overlies the Oued Teferguenit Formation. Dating [17,27,28] and comparisons with the Morrocan Anti-Atlas [5,22] allow us to place the main part of the FBV into the patulus conodont zone and into the Anarcestes lateseptatus ammonoid zone. The top of the FBV can be ranged into the lowermost Eifelian.
2.2 Facies of the Saoura FBV
It appears as a 12-m thick alternation of crinoidal limestones, marls and fine sandstones in the ‘km 30’ section. The limestones are packstone biomicrites with a shell-supported fabric. The marls yield a poorer and mud-supported fauna. The Cheffar el Ahmar formation is a marl-dominated facies with intercalations of limestones containing ammonoids and tentaculites with substantial decrease of the benthic fauna by comparison with the older levels. At Erg el Djemel, the FBV is only constituted of marls and thin pseudonodular bioclastic limestones. An alternation of clays and sandstones without shell concentration occurs between the lower and the upper part in the Haci-Feguaguira outcrop.
2.3 The brachiopod community
The brachiopods are dominated by articulate groups. The specific diversity is high, with 26 species belonging to 19 genera at ‘km 30’, 35 species and 29 genera at Erg el Djemel and 17 species and 16 genera at Haci-Feguaguira (Table 1). The associated fauna is represented by crinoids (ossicles and fragments), bryozoans, trilobites, ammonoids, orthocones, and scarce gastropods. In the ‘km 30’ section, we have defined three associations: (1) Alatiformia jaekeli association, (2) Athyris (A.) concentrica association and (3) Uncinulus orbignyanus association. At Erg el Djemel, the FBV contains only the second and the third associations. To the southeast, at Haci-Feguaguira, the third association is followed by a fourth association with Warrenella sp. cf. apodecta.
Les associations de brachiopodes dans les secteurs étudiés
Associations of brachiopods in the studied sectors
2.4 Eco-taphonomic features and interpretations
The substrate is either soft (marly levels) or bioclastic with taphonomic feedback [21] attested by several indications of a time of residence on the bottom (encrusting by epizoans, disarticulation, fragmentation) with phases of burial and exhumation (various states of preservation of the shells). No significant transport has intervened because no sorting is evident. Several species are represented by various ontogenetic stages documenting their eudemic nature [9]. Species indicating residence on the bottom (encrusting, fragmentation, disarticulation...) are abundant in the bioclastic levels. This suggests a taphonomic feedback when the rate of sedimentation is weak. This phenomenon causes the smoothing of line time (‘time averaging’) and increases diversity.
The sedimentological features of the Saoura FBV (texture packstone, pseudonodular beds) and the coexistence of benthic shells with nectobenthic goniatites and orthocones as well as the absence of erosion marks (waves or storms) indicate an accumulation in offshore environment, below storm waves. The tetracorallians (solitary and colonial) are abundant at the top of some beds, but their development has been quickly slowed down by the muddy sedimentation.
At Haci-Feguaguira, specific diversity and taphonomic feedback are weaker. The deposits are thicker (67 m) encompassing an episode of clays and sands (25 m) without shell beds.
The elementary cycle of the FBV consists of (i) a calcareous TST (transgressive system tract) with taphonomic feedback and condensation and (ii) a marly HST (high system tract). The whole of these cycles constitutes a major shell beds and marks the base of a transgressive interval. The correlation of constitutive brachiopod associations shows a disposition in onlap (Fig. 2) from the north (km 30) towards the south (Erg el Djemel) and to the southeast (Haci-Feguaguira). The association 3 with U. orbignyanus marks the maximum range of the FBV. The geographical extent of the FBV in all the Saoura region (350 km × 80 km) underlines its transgressive nature.
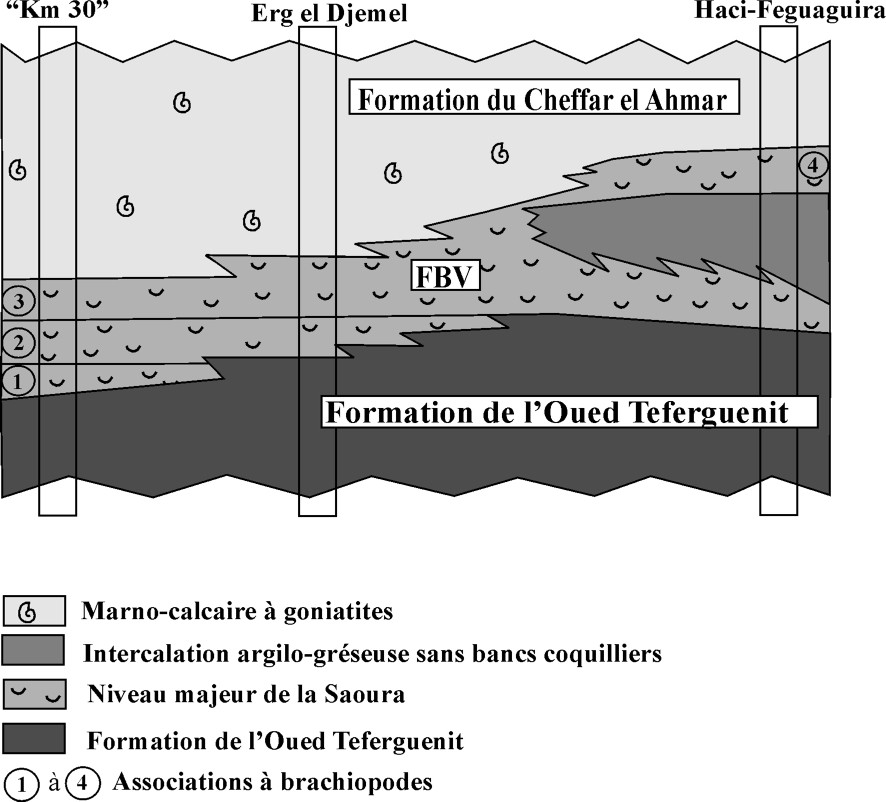
Corrélation des peuplements à brachiopodes et leur disposition en onlap dans le niveau majeur de la Saoura (les épaisseurs ne sont pas respectées).
Correlation of the brachiopod communities and their onlap distribution in the Saoura major level (without thickness scale).
2.5 Main characteristics of the major shell levels (Saoura FBV; Paris Basin Dogger and Maryland Miocene)
The following characteristic elements can be inferred from a comparison between the major levels of the Miocene of Maryland, the Dogger of Paris basin and the FBV of Saoura:
- – variation of the faunal concentration from ‘shell-supported’ to ‘mud-supported’ or ‘matrix-supported’;
- – significant specific richness (‘time-averaging’) and association of relatively varied groups of macrobenthos;
- – phenomena of taphonomic feedback with benthic fauna in various states of preservation (encrusting, dislocation, fragmentation, tapping...) testifying a time of residence on the bottom before the final burial;
- – great geographical extent that allows correlations at basin scale;
- – sequential position at the base of transgressive intervals or related to the maximum flooding period.
3 Conclusion: significance of the Saoura FBV in regional sedimentary dynamics
In Ahnet, the FBV is constituted by ferruginous oolitic limestones with Paraspirifer cultrijugatus [10]. In Mouydir, it changes in two main brachiopod levels (ferruginous conglomerates with Arduspirifer arduenensis and shell coquinas with Spirifer speciosus). They yield also a rich associated benthic fauna (crinoids, tabulates, trilobites, bryozoans). In the Tindouf basin, two levels have been described (Daïet el Aam and North-West Mabes [15]): the first with P. cultrijugatus, the second with Calceola sandalina (Lower Eifelian). At Bou-Bernous [15], a first level with Arduspirifer intermedius precedes beds with Pugnax pugnoides beds. Westward, the levels 74 and 78 of the Zemmour can be correlated to the FBV of the Saoura. Consequently, a correlation between the Saoura FBV and equivalent beds can be established all over the western and central Sahara.
These correlations are complicated, because these levels can be divided into two (Haci-Feguaguira) or three episodes (Tindouf). However, the FBV and its equivalents characterize a transgressive phase, beginning at the end of the Emsian and ending during the Earliest Eifelian, which is coeval with a change in the sedimentary mode. The siliciclastic inputs slow drastically or stop and the goniatite-bearing marl-limestone alternation spreads all over the western and central Sahara.
In conclusion, we can distinguish two kinds of major shell deposits: second-order major levels are developed within one basin alone [12,19] and first-order major shell deposits that are spread over several basins. The Saoura FBV is the single example so far known.
1 Introduction
Le concept des niveaux coquilliers (shell beds) ne cesse de se développer depuis la modélisation des « niveaux coquilliers majeurs » (= major shell deposits) du Miocène du Maryland [18–20]. Divers exemples ont été décrits en Nouvelle Zélande [26], au Japon [23], en Inde [11] et en France [12]. Il constitue une approche originale pour la détermination de la dynamique biosédimentaire, le repérage des cycles transgressifs–régressifs et les corrélations pour la reconstitution des bassins.
On distingue les niveaux coquilliers des séquences de dépôt du 5e au 7e ordre, liées aux fluctuations eustatiques de très haute fréquence (20, 40 et 100 ka), et les « niveaux majeurs », de grande extension géographique, liés aux cycles de 3e ordre (0,5 à 5 Ma). Ce concept a été appliqué aux niveaux à brachiopodes du Bathonien–Callovien du bassin de Paris [12,13], confirmant les condensations et la complexité des caractères éco-taphonomiques. Le rôle prépondérant de la rétroaction taphonomique [21] dans la genèse de ces repères est établi. Les déplacements horizontaux (recrutement) et les relais verticaux de la faune benthique suivent de près les cycles de variation du niveau marin relatif en adoptant des stratégies adaptatives (type r–K) et des innovations évolutives [14].
Une application et une illustration des caractéristiques d'un niveau majeur sont présentées ici à partir de l'étude du niveau à faune benthique variée (FBV) (= « coralligène » [25]) de l'Emsien supérieur de la Saoura (Fig. 1), dans le Sahara sud-oranais. Des assises contemporaines, riches en brachiopodes, tiges de crinoïdes et polypiers solitaires, peuvent lui être corrélées dans l'Ahnet et le Mouydir [10,24] au sud-est, dans le bassin de Tindouf [15] au sud-ouest et dans le Zemmour noir [32], à l'ouest. Cette vaste extension géographique permet une corrélation dans toute la partie occidentale de la plate-forme saharienne.
2 Caractéristiques physiques et signification environnementale du FBV de la Saoura
2.1 Position stratigraphique du FBV de la Saoura
La nomenclature lithostratigraphique adoptée est celle de Boumendjel et al. [3]. Le FBV de la Saoura, épais de 12 m, constitue la base de la formation du Cheffar el Ahmar, qui surmonte les grès fins et les calcaires bioclastiques du sommet de la formation de l'oued Teferguenit. Les datations proposées [17,27], ainsi que les résultats des travaux dans l'Anti-Atlas [4,22], permettent de bien situer biostratigraphiquement le FBV. Il se situe dans la zone à Patulus des conodontes (extrême sommet de l'Emsien) et dans la zone à Anarcestes lateseptatus des goniatites. Il se peut que le sommet du FBV atteigne la base de l'Eifélien, puisque au-dessus d'une combe de 3,50 m, qui surmonte le FBV, une barre calcaire a fourni Fidelites fidelis, Werneroceras (W.) rupachense et Anarcestes lateseptatus, ce qui est une association eifélienne aussi bien au Maroc qu'en Bohême [6]. La présence de Paraspirifer cultrijugatus dans le sommet du FBV s'accorde avec cette attribution, étant donné la distribution de ce marqueur aussi bien dans l'Eifel [31] que dans les Ardennes [5].
2.2 Faciès du FBV de la Saoura
Le FBV de la Saoura est constitué d'une alternance irrégulière d'encrinites, de marnes et de grès fins d'aspect ruiniforme. À l'Erg el Djemel, le FBV comprend une alternance de marnes grises et de calcaires bioclastiques, en bancs minces et pseudonoduleux. Dans la coupe de Haci-Feguaguira, le FBV se divise en deux unités séparées par une alternance argilo-gréseuse, sans concentration de faune.
Le faciès calcaire, de texture packstone, est constitué d'encrinites, parfois argileuses, contenant divers débris de coquilles benthiques et un ciment micritique. Ces bancs calcaires pseudonoduleux (bioturbation) contiennent une macrofaune benthique variée : fragments de pédoncules et d'articles de crinoïdes, brachiopodes, tétracoralliaires tabulés et rugueux, trilobites, orthocères, bryozoaires et quelques rares gastéropodes. On n'observe, ni relief, ni trame squelettique [28]. Les coquilles sont concentrées par endroits dans les calcaires (shell-supported) et quelques concrétions ferrugineuses (2 à 5 mm de diamètre) sont occasionnellement rencontrées.
Les intercalations marneuses, de couleur grise ou verte, livrent une faune moins concentrée (mud-supported) et moins riche. Elles admettent de minces passées de grès à grain fin à moyen. Elles sont bioturbées et leur ciment, carbonaté à la base, devient progressivement argileux vers le sommet, ce qui leur confère un aspect ruiniforme. Elles contiennent des trilobites et des terriers horizontaux ou obliques.
Au-dessus, l'essentiel de la formation du Cheffar el Ahmar est dominé par une alternance marno-calcaire à goniatites et tentaculites. Les niveaux calcaires correspondent à des biocalcilutites de texture mudstone à wackestone, à rare faune benthique.
2.3 Les peuplements de brachiopodes
La diversité spécifique est élevée, avec 26 espèces réparties en 19 genres au « km 30 », 35 espèces représentant 29 genres à l'Erg el Djemel et, enfin, à Haci-Feguaguira, 17 espèces représentées par 16 genres (Tableau 1). Une première liste des taxons a été précédemment publiée [3,25]. Les groupes associés sont représentés par des crinoïdes (articles et fragments de pédoncules), des tétracoralliaires (solitaires et coloniaux, [25,29]), des bryozoaires, des trilobites [3,25], des orthocères, des goniatites [17,25] et quelques rares gastéropodes.
L'absence totale des stromatopores semblent expliquer, en partie, l'absence de bioconstruction, alors que leur association avec les madréporaires aboutit aux biohermes et biostromes, tels ceux des Ardennes [5], du Massif armoricain, de Bohême et de plusieurs secteurs de la plate-forme canadienne [7].
Dans le secteur du « km 30 », nous avons défini trois associations : (1) association à Alatiformia jaeckeli, (2) association à Athyris (A.) concentrica et (3) association à Uncinulus orbignyanus. À l'Erg el Djemel, le FBV ne contient que les associations 2 et 3. Vers le sud-est, à Haci-Feguaguira, l'association 3 est suivie par une quatrième association, à Warrenella sp. cf. apodecta (Tableau 1).
2.4 Caractères éco-taphonomiques et interprétations
Le FBV de la Saoura montre une alternance de substrats meubles (niveaux marneux) et durs (bioclastiques), à rétroaction taphonomique attestée par divers indices témoignant d'un temps de résidence sur le fond (encroûtements par épizoaires, taraudages, désarticulation, fragmentation, agglutination). Des phases d'enfouissement et d'exhumation rendent compte des différents états de préservation des coquilles. Il ne semble pas qu'il y ait eu un transport important, puisque aucun tri n'est apparent. Plusieurs espèces sont représentées par différents stades ontogéniques attestant le caractère eudémique (entités paléobiologiques dont les représentants ont été récoltés dans un milieu où ils ont vécu et se sont reproduits [9]) de ces brachiopodes. Certains exemplaires (Galeatagypa pelagicus ou encore Sieberella costata), représentés uniquement par des exemplaires adultes, pourraient être adémiques (les milieux de vie sont distincts des milieux de reproduction).
Les espèces comme Atrypa (Planatrypa) squamifera et Oglu semirobis pullulent surtout dans les faciès boueux (soft-bottom) ; cette association est connue ailleurs : en Pologne dans les monts de Sainte-Croix [2], en Belgique [16], en France dans le Massif armoricain [8], souvent dans des niveaux marneux. Cette faune des faciès boueux contient peu d'exemplaires encroûtés.
Des espèces telles que Sieberella sieberi, S. costata, Uncinulus orbignyanus, Janius irbitensis et Arduspirifer gr extensus-maturus sont plutôt liées aux substrats durs (bioclastiques). Les espèces présentant des indices de résidence sur le fond (encroûtement, fragmentation, désarticulation...) sont abondantes dans ces niveaux bioclastiques, ce qui suggère la récurrence de la rétroaction taphonomique lorsque le taux de sédimentation est faible. Ce phénomène provoque le lissage des lignes-temps (time averaging), augmentant la diversité spécifique (niveaux calcaires), et des condensations de type intra-habitat, puisque l'environnement sédimentaire est le même.
Le scénario que nous envisageons pour chaque cycle de séquence-dépôt se résume ainsi :
- – phase de ralentissement de la sédimentation (quelques concrétions ferrugineuses) ;
- – apports latéraux épars d'articles et de fragments de pédoncules de crinoïdes par des courants de fonds « latents », non érosifs, assurant les apports nutritifs et une oxygénation du fond ; la colonisation par les organismes pionniers intervient sur un fond boueux (soft-bottom), suivie par plusieurs générations (time-averaging) qui développent un substrat dur par accumulation des coquilles et rétroaction taphonomique (brachiopodes et crinoïdes) ; l'épifaune robuste (madréporaires) s'installe ensuite ;
- – déclenchement de la sédimentation boueuse qui bloque la rétroaction taphonomique et provoque l'étouffement des madréporaires ; une partie de la faune benthique prolifère, car elle tolère une certaine turbidité (A. (P.) squamifera, A. (P.) collega et O. semiorbis) [2,8,16].
Le recrutement paraît continu ; la diversité des micro-habitats (shelly-bottom et soft-bottom) permet une prolifération d'une macrofaune benthique. Les condensations induites par la rétroaction taphonomique sont fréquentes dans les niveaux calcaires.
Les caractères sédimentologiques (texture packstone) et la coexistence des organismes benthiques avec des coquilles de goniatites et d'orthocères ainsi que l'absence de figures d'érosion (de vagues ou de tempêtes) nous permettent de proposer une mise en place en milieu offshore, avec des courants intermittents qui permettent la concentration des éléments squelettiques. La diversité spécifique s'explique par le time averaging dû au faible taux de sédimentation. Les communautés benthiques ne semblent pas en réelle compétition. Il est probable qu'il s'agissait d'un biotope où des courants, non violents, balayaient le fond marin fournissant une part importante de la matière nutritive à la faune hétérotrophe en plus des « pluies » planctoniques.
Les tétracoralliaires (solitaires et coloniaux) ont colonisé des fonds parsemés de microhabitats, ce qui a induit une instabilité des peuplements. Leur prolifération aux sommets de certains bancs est rapidement freinée par la sédimentation boueuse. Il n'y aurait ni piégeage des sédiments ni, en conséquence, bioconstruction. Les formes solitaires arrivent à se maintenir malgré la turbidité de l'eau. Certains auteurs [7] estiment que des remaniements importants peuvent donner des ensembles à coraux dépourvus de trame squelettique ; l'absence de figures de tempêtes écarte cette hypothèse dans le cas du FBV de la Saoura. La cimentation micritique peut être assurée par la dissolution de la microfaune ou par précipitation directe.
Dans la coupe de Haci-Feguaguira, la diversité spécifique étant plus faible et les indices de rétroaction taphonomique moindres, la condensation est faible. En effet, l'épaisseur est plus importante (67 m avec un épisode d'environ 25 m d'argilo-gréseux sans niveaux coquilliers).
Les modèles établis par divers auteurs sur les niveaux coquilliers [11,12,18,23] s'accordent tous sur la relation entre ces niveaux majeurs et le début du régime transgressif. D'autres niveaux moins complexes se développent lors du maximum d'inondation (maximum flooding zone ou MFZ [11]).
Le cycle élémentaire du FBV est constitué par un TST (cortège transgressif) calcaire à rétroaction taphonomique et condensation et un HST (cortège de haut niveau) marneux. L'ensemble des cycles constitue un niveau majeur à brachiopodes qui marque la base d'un intervalle transgressif (Fig. 2). Les corrélations des associations constitutives montrent en effet une disposition en onlap du nord ( « km 30 ») vers le sud (Erg el Djemel) et le sud-est (Haci-Feguaguira). L'association 3 à U. orbignyanus marque l'extension maximale du FBV. L'extension géographique du FBV dans toute la Saoura et l'Erg el Djemel (350 km × 80 km) souligne bien ce caractère transgressif.
2.5 Caractéristiques principales des niveaux majeurs (FBV de la Saoura ; Dogger du bassin de Paris et Miocène du Maryland)
Une comparaison entre les niveaux majeurs du Miocène du Maryland, du Dogger du bassin de Paris et du FBV de la Saoura fait ressortir les éléments caractéristiques suivants :
- – niveaux coquilliers riches en faune benthique permettant de les distinguer par rapport aux unités lithologiques encaissantes ; la concentration varie du shell-supported au mud-supported ou matrix-supported ;
- – richesse spécifique importante (time averaging) et association de groupes de macrobenthos relativement variés ;
- – phénomènes de rétroaction taphonomique avec différents états de préservation de la faune benthique (encroûtements, désarticulation, fragmentation, taraudage...), témoignant d'un temps de résidence sur le fond avant l'enfouissement définitif ;
- – grande extension géographique permettant des corrélations à l'échelle du bassin ;
- – position séquentielle, à la base des intervalles transgressifs ou en relation avec le maximum d'inondation.
Les variations peuvent concerner les caractères sédimentologiques (niveau d'énergie, figures sédimentaires, intensité de la bioturbation, biseautage...) liés à la juxtaposition d'environnements contigus. La différence essentielle porte sur la nature des peuplements ; en effet, les niveaux majeurs de l'Emsien supérieur et du Dogger sont dominés par les brachiopodes, alors que ceux du Miocène sont riches en mollusques. Cette différence est liée aux changements intervenus dans les peuplements au cours des temps géologiques [1,30]. Les caractères d'ensemble sont identiques : fréquence des niveaux riches en macrofaune benthique à rétroaction taphonomique, time averaging, grande diversité faunique et grande extension géographique.
3 Conclusion : signification du FBV de la Saoura dans la dynamique sédimentaire régionale
Un niveau majeur à brachiopodes est caractérisé par sa continuité latérale et permet une corrélation à l'échelle d'un bassin [12]. Le FBV de la Saoura est représenté dans tous les affleurements du passage Dévonien inférieur–Dévonien moyen de la chaîne d'Ougarta. Les associations de brachiopodes sont finement corrélées et la disposition en onlap vient confirmer la relation entre le régime transgressif et le développement d'un niveau benthique repère. À l'échelle de l'Ouest saharien, les corrélations entre le FBV et les niveaux repères des autres régions sont précises.
Le niveau à Paraspirifer cultrijugatus de l'Ahnet s'est développé dans des calcaires oolithiques ferrugineux [10]. Dans le Mouydir, il existe au moins deux niveaux principaux à brachiopodes (conglomérats ferrugineux à Arduspirifer arduenensis et lumachelle à Spirifer speciosus). Ces niveaux sont aussi riches en faune benthique associée (crinoïdes, tabulés, trilobites, bryozoaires). Dans le bassin de Tindouf, sur la coupe de Daïet el Aam et celle du Nord-Ouest de Mabès [15], on peut déceler deux niveaux : le premier à P. cultrijugatus et le deuxième à Calceola sandalina. Dans la coupe de Bou-Bernous [15], le premier niveau contient Arduspirifer intermedius et le second livre Pugnax pugnoïdes. Dans le Zemmour noir [32], les niveaux 74 et 78 correspondent au FBV de la Saoura.
L'épisode (FBV) caractérise une phase transgressive que nous pouvons suivre sur toute la plate-forme ouest-saharienne. Cette transgression, qui s'échelonne de la fin de l'Emsien jusqu'à l'Eifélien inférieur, accompagne le changement du régime sédimentaire ; en effet, les silico-clastiques cessent dans la Saoura et l'Ahnet après ce niveau majeur et deviennent accessoires dans le bassin de Tindouf et dans le Zemmour noir. La sédimentation marno-calcaire à goniatites se généralise sur tout le Sahara occidental après cet épisode.
Ainsi, un niveau coquillier majeur correspond à un niveau d'extension à l'échelle de tout un bassin [12]. Celui de la plate-forme occidentale du Sahara africain s'étend sur plusieurs bassins. De ce fait, nous pouvons distinguer des niveaux majeurs d'ordre 2 qui se développent dans un seul bassin (exemple des niveaux majeurs du Dogger du bassin de Paris et du Miocène du Maryland) et des niveaux majeurs d'ordre 1 corrélables entre plusieurs bassins. Le FBV de la Saoura constitue le seul exemple de ce type connu jusqu'à ce jour.
Acknowledgments
Ce travail s'inscrit dans le programme 02 MDU 555, Lyon1-Oran Différenciation et caractérisation des stades d'ouverture des bassins intracratoniques d'Algérie. Nous remercions A. Boullier (Besançon) pour ses commentaires constructifs, ainsi que le deuxième rapporteur.