

Abridged English version
The objective of this study is to test the hypothesis of the specificity of the vapour isotopic signature transpired by plant canopy. Isotopic models of atmospheric general circulation assume that when the leaf water content reaches an isotopic steady state, the heavy isotope content of transpiration vapour is identical to that of the precipitation water absorbed by plant roots in deep soil layers. This concerns rainwater that would have escaped by soil evaporation due to fast infiltration in the deep soil layers. Nevertheless, as the roots developed in surface soil layers have access to water subjected to evaporation, it requires the precise determination of the soil depth which is affected by root water uptake. The isotopic methodology (18O, 2H) can allow this determination by comparing isotopic signatures of the soil water profile to that of the water extracted at the level of the first internodes of the plant stem. This comparison can be carried out with the caveat that (i) no isotopic fractionation occurs during root water uptake, (ii) there is a vertical isotopic gradient of water in the soil profile, (iii) the roots are distributed according to soil depth, and (iv) there is a moisture gradient in the soil. To test in temperate regions the hypothesis used in the isotopic models of atmospheric general circulation, an experiment was carried out on a research plot cultivated in maize (Nemours, Seine et Marne, France). During a 24-h period, the flow of evapotranspiration and the isotopic composition of the soil water and of that of the plants (leaves, stems taken at the level of the first internodes) were measured. After sampling, soil and plant water samples were vacuum extracted. The 18O signature of extracted water samples was classically analysed with an isotopic mass spectrometer equipped with multiple collectors after equilibration between water and carbon dioxide. The deuterium content was analysed after extracting the hydrogen of the water by reduction on uranium metal. The root density and soil water content were also measured at the time of sampling. Contrary to what is usually reported in the literature, the water isotopic composition of the first stem internodes of the maize plant evolves through time. During the daily cycle, this composition decreases appreciably between 03 h 00 and 15 h 00, then increases afterwards to reach the initial isotopic content observed during the previous night. Three hypotheses can be invoked to justify the temporal isotopic evolution of the water of the first internodes. The first hypothesis calls upon the double circulation in the stem saps, the ascending xylem sap having an isotopic signature close to that of soil water, while the descending phloem sap is comparatively more enriched in the heavy isotope. However, heavy isotope content of first internode water reached a minimal value when the heavy isotope content of leaf water was highest. The increase in the heavy isotope concentration of the phloem sap should consequently cause that of the stem water, which is not observed here and thus calls for the rejection of this first hypothesis. A second hypothesis takes into account the process of hydraulic lift where water extracted by roots in deep soil layers during the night is released in the upper horizon by shallower roots during the day. Because of the existence of an isotopic gradient in soil water, this insertion should cause a considerable reduction in the isotopic signature of the surface layer soil water, which was not observed here and therefore calls for the rejection of this second hypothesis. A third hypothesis suggests the evolution of water quantity absorbed by the roots during the day in response to the evaporating request evolution. The examination of the diagram δ2H–δ18O shows that the water isotopic signature of the first internodes ranges between that of the soil water of surface layers and that of the deep layers. It is however not possible that all soil water extracted by roots comes from a single horizon. To solve this problem, we considered that soil water extraction is directly proportional to the amount of roots present in each soil horizon, and therefore that the isotopic signature at the first internode can be described by:
| (4) |
This experiment invalidates the classical hypothesis used in isotopic models of general atmospheric circulation in temperate regions. In fact, only half the amount of water vapour transpired by the canopy during the day presents a signature similar to rainwater sampled in deep soil layers.
1 Introduction
Les modèles isotopiques de circulation générale atmosphérique [16,17] et différentes études hydrologiques [20,21] cherchent à prendre en compte l'évapotranspiration réelle pour compléter les bilans hydriques continentaux. Ceci a conduit à admettre l'hypothèse d'un simple recyclage par la transpiration de l'eau prélevée par les racines et donc à négliger l'évaporation de l'eau dans le sol. À l'état isotopique stationnaire de l'eau au sein du réservoir foliaire, la plante agirait comme une simple mèche [19]. Dans ce contexte, la signature de la vapeur issue de la transpiration serait donc identique à celle de la pluie infiltrée dans le sol et prélevée par les racines. Dans un diagramme δ2H–δ18O, la signature isotopique de la vapeur d'eau issue de la transpiration appartiendrait donc à la droite locale des eaux météoriques. L'évaporation provoque une modification très sensible de la signature isotopique des précipitations infiltrées dans les couches superficielles du sol [4,8,18]. De plus, le réseau racinaire est souvent largement développé dans ces mêmes couches superficielles. Ce réseau permet donc à la plante d'être alimentée par une eau concentrée en isotopes lourds par l'évaporation. Enfin, l'extraction racinaire modifie fortement la répartition de l'eau dans les différents horizons du sol et donc modifie sa disponibilité vis-à-vis des couverts végétaux. Cela n'est bien évidemment pas sans conséquence sur les plans agronomique et hydrologique. Les déterminations de l'humidité volumique dans le sol, du potentiel hydrique, de la conductivité hydraulique, de l'architecture racinaire, de la chimie des solutions de sol et de la demande évaporatoire sont des paramètres indispensables à la compréhension des transferts hydriques liés à l'extraction racinaire. Cependant, l'ensemble de ces paramètres peut être difficile, sinon impossible, à acquérir in situ, que ce soit dans le temps ou dans l'espace. Ceci entraîne une difficulté certaine pour l'identification des zones actives d'absorption racinaire de l'eau dans le sol.
La détermination de la profondeur du sol à laquelle l'eau est extraite par les racines reste donc un problème d'actualité. Le traçage isotopique naturel de l'eau peut permettre d'effectuer cette détermination [11,12,22] en raison de l'absence de fractionnement isotopique lors de l'absorption racinaire [25] et de l'existence d'un gradient isotopique de l'eau dans le sol. Ce gradient permet d'individualiser chaque couche de sol à travers sa signature isotopique. La détermination de la zone d'extraction racinaire est réalisée classiquement à partir de la comparaison entre le profil isotopique vertical de l'eau dans le sol et la teneur en isotopes lourds de l'eau, mesurée au niveau du tronc, du collet ou du premier entre-nœud de la tige [2,24]. Ces compartiments peuvent être considérés comme des collecteurs globaux dans lesquels est réalisée l'homogénéisation des différentes signatures de l'eau absorbée par les racines à différentes profondeurs.
Nous chercherons dans le cadre de ce travail, réalisé à l'échelle locale, à tester l'hypothèse d'un simple recyclage par la transpiration de la pluie infiltrée dans les sols, afin d'examiner la possibilité d'une transposition de cette hypothèse à des échelles supérieures. Cette hypothèse sera testée dans le contexte d'un couvert de maïs développé en région tempérée.
2 Matériels et méthodes
L'échantillonnage a été réalisé les 20 et 21 juillet 2000 sur une parcelle expérimentale (1 ha) plantée en maïs (Nemours, Seine-et-Marne, France). Au cours d'une période de 24 h, nous avons prélevé des échantillons de sol (quatre carottages), à différentes profondeurs (de la surface du sol à ), et de végétaux (premier entre-nœud de la tige de maïs et feuilles de plants à une même hauteur de 1,80 m). Le flux d'évapotranspiration a été déterminé à partir de la méthode du rapport de Bowen.
Après prélèvement, l'extraction de l'eau des échantillons de sol et de végétaux est effectuée au laboratoire par distillation sous vide. Les analyses isotopiques sont réalisées classiquement après une équilibration isotopique H2O–CO2 pour l'oxygène 18 [13] ou après réduction de l'eau sur de l'uranium pour le deutérium [5,15]. Les résultats sont exprimés en unités δ (équation (1)), par référence à l'étalon international V-SMOW [14] :
| (1) |
La reproductibilité analytique est de pour l'oxygène 18 et de pour le deutérium.
La densité racinaire a été mesurée par méthode gravitaire après séparation mécanique par dispersion sous jet d'eau de la matière organique et minérale et tamisage à 1 mm.
3 Résultats
3.1 Signature isotopique de l'eau du sol (Fig. 1)
L'eau des couches superficielles du sol présente simultanément les teneurs moyennes en isotopes lourds les plus élevées (, ) et les teneurs en eau les plus faibles (). Puis, en profondeur, les teneurs isotopiques diminuent rapidement pour atteindre une valeur moyenne sensiblement constante à partir de (, ). L'humidité moyenne pondérale augmente fortement jusqu'à une valeur sensiblement constante () à partir de .
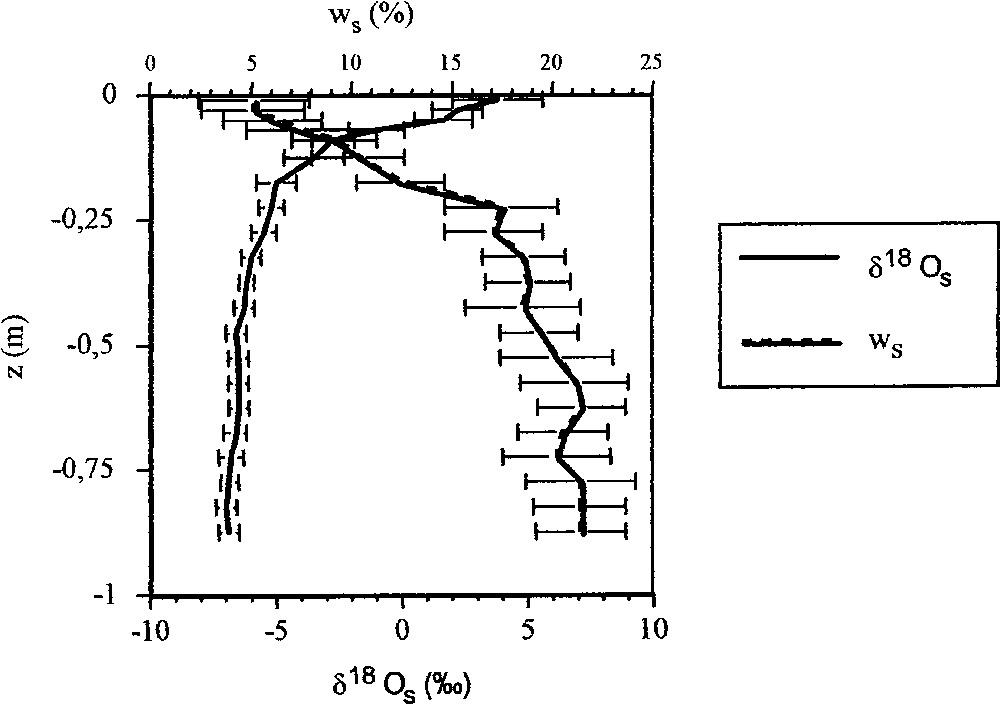
Évolution verticale de l'humidité pondérale moyenne du sol et de la teneur moyenne en oxygène 18 de l'eau extraite à différentes profondeurs de quatre carottes de sol (Nemours, Seine-et-Marne). Les écarts types sont également présentés.
Vertical profile of the average soil water content ( weight percent) and of the average oxygen-18 concentration of the water extracted at different depths form four soil cores (Nemours, Seine-et-Marne, France). Standard deviations of the measurement uncertainties are also given.
L'enrichissement isotopique enregistré à la surface du sol est caractéristique de l'évaporation (Fig. 2), comme le montre la répartition, dans un diagramme δ2H–δ18O, des teneurs isotopiques le long d'une droite dont la pente (3), (4) est caractéristique de ce processus (équation (2)) :
| (2) |

Évolutions, dans un diagramme – des signatures isotopiques de l'eau du sol, du premier entre-nœud et des feuilles des plants de maïs prélevés les 20 ef 21 juillet 2000 à Nemours (Seine-et-Marne). Les évolutions de la signature isotopique de l'eau du premier entre-nœud et de l'eau du sol sont également présentées dans l'encart.
Changes in the isotopic signatures (–) of the soil water, the water at the first internode of the stems of the plants, and of the leaves of the maize collected on 20 and 21 July 2000 at Nemours (Seine-et-Marne). The insert shows the changes in the isotopic signature of the water in the first internode of the stems of the plants and in the soil.
Il est à noter que les teneurs isotopiques de l'eau des couches profondes du sol restent très proches des valeurs qui caractérisent la droite des eaux météoriques ( [9]).
La densité racinaire devient maximale (48 % des racines dans les 15 premiers centimètres) à la profondeur du sol où la signature isotopique de l'eau atteint les valeurs les plus élevées et l'humidité pondérale, les valeurs les plus faibles (Fig. 3).
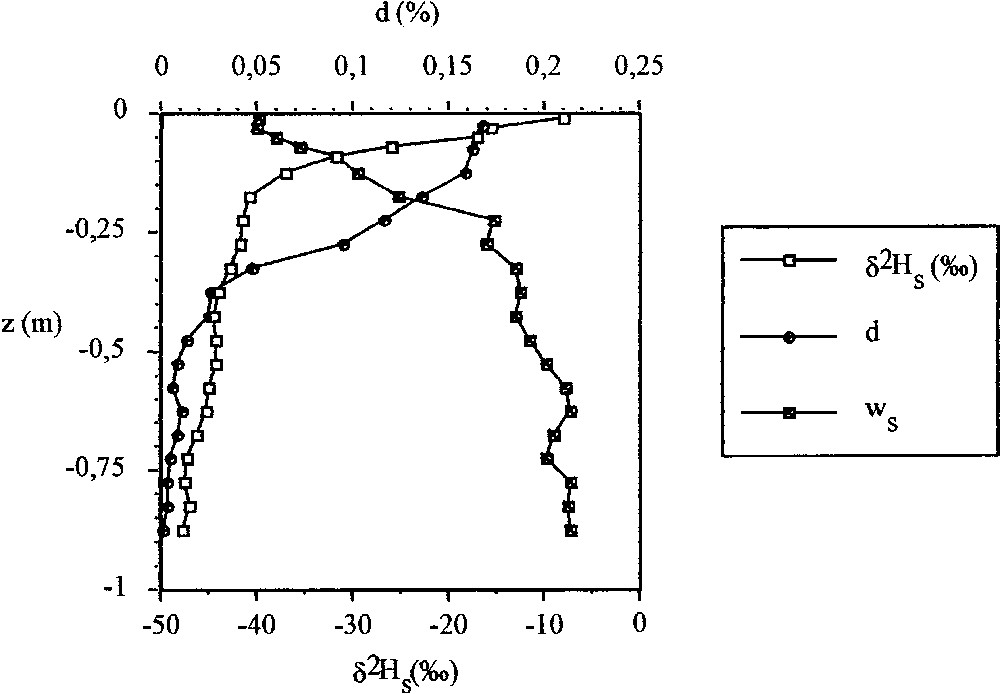
Évolution en fonction de fa profondeur (z) de l'humidité pondérale (w), de la densité racinaire (d) et de la teneur en deutérium de l'eau des échantillons de sol prélevés à Nemours (Seine-et-Marne) les 20 et 21 juillet 2000.
Evolution with depth (z) of the soil water content , the root density (d) and the deuterium concentration of the soil water collected at Nemours (Seine-et-Marne, France) on 20 and 21 July 2000.
Les compositions isotopiques de l'eau du sol, mesurées à une même profondeur, ne montrent pas d'évolution significative au cours du temps.
3.2 Signature isotopique de l'eau des plants de maïs (premier entre-nœud de la tige, feuille, Fig. 4)
La teneur en isotopes lourds de l'eau du premier entre-nœud présente une évolution sensible au cours du temps. Le 20 juillet 2000, au cours de la période 03 h 00–16 h 00, cette teneur évolue de à pour l'oxygène 18 et de à pour le deutérium. Puis elle tend à revenir à l'état initial nocturne, qui est atteint à 02 h 00 le 21 juillet 2000. Dans un diagramme δ2H–δ18O (Fig. 2), les teneurs en isotopes lourds de l'eau de l'entre-nœud appartiennent à la droite d'évaporation définie précédemment (équation (2)).
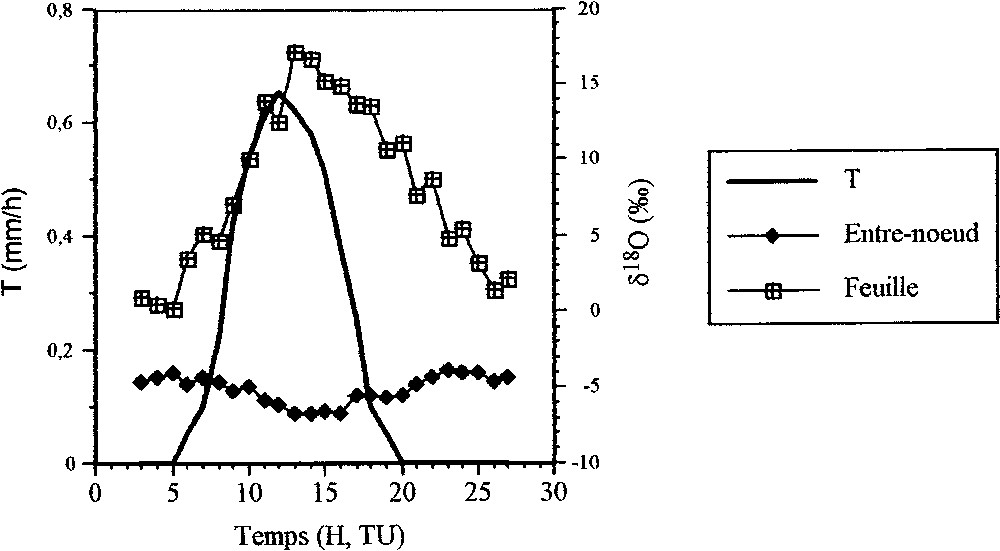
Évolution temporelle du flux de transpiration (T) et des teneurs en oxygène 18 de l'eau du premier entre-nœud et des feuilles des plants de maïs prélevés à Nemours (Seine-et-Marne) les 20 et 21 juillet 2000.
Evolution with time of the transpiration flux (T) and of the oxygen-18 concentration of the water at the first internode of the stems and in the leaves of the maize plants collected at Nemours (Seine-et-Marne, France) on 20 and 21 July 2000.
La composition isotopique de l'eau des feuilles prélevées à la hauteur de 1,80 m présente une évolution temporelle très importante (18,1‰ pour l'oxygène 18 et 36,4‰ pour le deutérium, Fig. 4). Cette évolution reste globalement parallèle à celle du flux d'évapotranspiration (Fig. 4), avec un déphasage d'environ 1 h pour les valeurs maximales. Dans un diagramme δ2H–δ18O, les teneurs isotopiques de l'eau foliaire se répartissent suivant une nouvelle droite d'évaporation (Fig. 2) :
| (3) |
4 Discussion
L'examen du diagramme δ2H–δ18O (Fig. 2) montre que la signature isotopique de l'eau du premier entre-nœud évolue entre celle de l'eau des couches superficielles du sol ( « pôle évaporation ») et celle de l'eau des couches profondes ( « pôle précipitation »). D'une manière générale, les teneurs isotopiques de l'eau du sol et des racines ne peuvent être jugées représentatives du signal d'entrée dans la plante. Ceci est dû à la forte hétérogénéité verticale des signatures isotopiques de l'eau dans le sol et à la difficulté d'accéder à l'ensemble du réseau racinaire in situ. En revanche, au premier entre-nœud, considéré comme collecteur commun à l'ensemble du réseau racinaire, l'homogénéisation des teneurs isotopiques de l'eau prélevée par les racines à différentes profondeurs est réalisée.
Trois hypothèses peuvent permettre d'expliquer une évolution temporelle de la signature de l'eau du premier entre-nœud :
- (a) l'hypothèse du double marquage isotopique lié à la circulation des sèves dans la tige [3] : la composition isotopique des eaux qui circulent dans la tige serait le produit d'un double marquage isotopique, lié d'une part, à l'eau du sol (sève brute) et d'autre part, à la transpiration foliaire (sève élaborée) ;
- (b) l'hypothèse de l'ascenseur hydraulique dans le sol [6,7,10] : l'eau, prélevée la nuit dans les couches profondes et humides du sol, serait réintroduite le jour dans les couches superficielles plus sèches. Vis-à-vis de l'eau des couches superficielles du sol, les racines se conduiraient donc à la fois comme une source et un puits. Le développement de ce stock d'eau temporaire dans les couches superficielles du sol est réputé permettre aux couvertures végétales de renforcer la lutte contre les contraintes hydriques. L'insertion dans les couches superficielles du sol d'une eau prélevée en profondeur devrait donc conduire à une diminution de la teneur en isotopes lourds de l'eau des couches superficielles et donc du signal mesuré au premier entre-nœud. Dans le cadre de cette hypothèse, seules les racines des couches superficielles participeraient à l'extraction de l'eau dans le sol. Cette hypothèse pourrait justifier la répartition des teneurs en isotopes lourds de l'eau du premier entre-nœud le long de la droite d'évaporation (équation (2)) dans un diagramme δ2H–δ18O ;
- (c) l'hypothèse de l'évolution de la quantité d'eau prélevée en profondeur par les racines, en relation avec la demande évaporatoire.
Au cours de la journée, lorsque la quantité d'eau présente dans les couches superficielles du sol devient insuffisante au regard de l'augmentation de la transpiration, les racines mobiliseraient l'eau disponible dans des couches plus profondes. Pendant la nuit, la transpiration nocturne devient très faible : les racines n'ont donc plus à extraire des quantités d'eau importantes en profondeur. Cette évolution simultanée de la quantité d'eau extraite et de la profondeur du sol à laquelle cette eau est prélevée provoquerait donc l'arrivée, au premier entre-nœud des plants de maïs, d'eaux marquées différemment en isotopes lourds. De même, comme dans l'hypothèse précédente, ceci pourrait justifier, dans un diagramme δ2H–δ18O, la position occupée par la signature isotopique de l'eau du premier entre-nœud de la tige des plants de maïs.
La première hypothèse du double marquage isotopique des sèves ne peut être retenue ici. En effet, l'eau des feuilles est concentrée fortement en isotopes lourds au moment où la transpiration atteint les valeurs les plus élevées (Fig. 4). La teneur maximale en isotopes lourds de l'eau foliaire ( pour l'oxygène 18) est atteinte à 15 h 00, heure à laquelle la teneur isotopique de l'eau du premier entre-nœud devient minimale (). L'arrivée de la sève élaborée au premier entre-nœud devrait, au contraire, provoquer une augmentation de sa teneur en isotopes lourds. Or, c'est la situation inverse qui est observée lors de cette expérience, ce qui conduit donc au rejet de cette hypothèse.
L'hypothèse d'une réintroduction par les racines, dans les couches superficielles du sol, d'une eau prélevée en profondeur (et dont le signal est proche de celui des précipitations) ne peut également être retenue. En effet, cette insertion devrait se traduire par une diminution du signal isotopique de l'eau dans ces couches superficielles. Cette situation n'est pas observée au cours de cette expérimentation (Fig. 5), d'où le rejet de cette deuxième hypothèse.

Évolution temporelle de la teneur en oxygène 18 de l'eau prélevée dans le sol à différentes profondeurs (Nemours, Seine-et-Marne, 20 et 21 juillet 2000).
Evolution with time of the oxygen-18 concentration in the soil water at different depths (Nemours, Seine-et-Marne, France, 20–21 July 2000).
La troisième hypothèse prend en compte l'évolution de la quantité d'eau prélevée en profondeur par les racines, en réponse à l'augmentation de la transpiration. Contrairement à ce qui est classiquement réalisé dans la littérature, il semble difficile d'attribuer à une seule profondeur donnée le poids de la totalité de l'extraction racinaire. En revanche, on peut chercher à reconstituer le signal d'entrée dans la plante à partir d'un simple bilan isotopique, réalisé couche de sol par couche de sol (équation (4)) :
| (4) |
Ceci suppose que l'extraction racinaire soit proportionnelle à la quantité de racines présente dans une couche de sol donnée, ce qui constitue bien évidemment une forte hypothèse en raison du degré de dessiccation plus ou moins prononcé du sol.
L'application de l'équation (4) à l'ensemble des profils (isotopes et densité racinaire) conduit à des teneurs calculées ( : , : ) supérieures à celles mesurées tout au long du cycle nycthéméral. L'élimination progressive, à partir de la surface du sol (équation (4)), des quantités d'eau supposées être prélevées par les racines, permet de restituer, dans un diagramme δ2H–δ18O, une évolution satisfaisante de la signature issue de l'extraction racinaire au cours de la journée (Fig. 6). Cette représentation confirme, en particulier, qu'au cours de la période 12 h 00–16 h 00, au moment où la transpiration devient maximale, l'alimentation hydrique de la plante provient essentiellement des couches profondes de sol. Cela se traduit par l'arrivée, à l'entre-nœud, d'une eau marquée en isotopes lourds par les précipitations. C'est donc cette troisième hypothèse que nous retiendrons pour expliquer l'évolution temporelle de la signature isotopique de l'eau du premier entre-nœud des plants de maïs. On peut supposer que le réservoir d'eau foliaire atteigne très rapidement l'état isotopique stationnaire [1,23] et restitue à l'atmosphère la signature isotopique de l'eau de l'entre-nœud. Cela se traduit donc par l'introduction dans l'atmosphère de vapeurs marquées différemment en isotopes lourds, en raison de la variation de la quantité d'eau prélevée en profondeur et de l'évolution de la composition isotopique du mélange qui en résulte au niveau de l'entre-nœud.

Comparaison entre les compositions isotopiques de l'eau mesurées au premier entre-nœud des plants de maïs et calculées à partir de l'équation (4). Les horaires des prélèvements sont indiqués au-dessus du symbole des teneurs mesurées.
Comparison between the isotopic composition of the water at the first internode of the stems and as calculated from Eq. (4). The time of sample collection is given above the symbols for the measurements.
En revanche, cette expérimentation invalide l'hypothèse évoquée précédemment dans le contexte des modèles isotopiques de circulation générale atmosphérique. En effet, si l'on ne retient dans le bilan de masse de la vapeur transpirée que la quantité mesurée durant les heures de la journée où la plante prélève majoritairement de l'eau dans les couches profondes du sol, on constate qu'il n'y a que 55 % de cette vapeur qui présente une signature isotopique identique à celle des précipitations.
5 Conclusion
La détermination de la composition isotopique de l'eau du premier entre-nœud des plants de maïs permet de suivre dans le sol l'évolution spatio-temporelle de la profondeur où l'eau est mobilisée par les racines. Bien évidemment, il ne s'agit encore que d'une approche préliminaire de l'analyse de l'absorption racinaire de l'eau à partir de la méthodologie isotopique. Cette étude est rendue possible par l'existence d'un réseau racinaire développé sur tout le profil du sol analysé, d'un dessèchement du sol dans les couches superficielles, d'un gradient isotopique de l'eau dans le sol en fonction de la profondeur et par l'absence de fractionnement isotopique de l'eau du sol lors de l'extraction racinaire.
Cette expérience infirme, en régions tempérées, l'hypothèse retenue dans les modèles isotopiques de circulation générale atmosphérique. En effet, seule la moitié de la quantité de vapeur d'eau émise par la plante au cours de la journée présente une signature identique à celle de l'eau des précipitations prélevées dans les couches profondes du sol.
Acknowledgments
Cette étude a été réalisée dans le cadre des programmes de recherche CNRS–CNRST (France–Maroc, contrat STU./07) et du PNRH (Insu–Inra–BRGM–Cemagref–Météo France–Cnes–IRD–LCPC–CIRAD–CNRS (SDU, SC, SdV, SPI)]. Nous tenons tout particulièrement à remercier les deux rapporteurs (Roberto Gonfiantini et un rapporteur anonyme) de cet article, ainsi que Ghislain de Marsily, pour leurs remarques particulièrement constructives.