

1 Introduction
Recent palaeoecological studies have underlined the importance of multi-proxy programs in order to improve the accuracy of investigations concerning past environment and climate reconstructions. This allows evaluation of a maximum of ecological factors before postulating climatic models. Nevertheless, classical long sequences from mid-Europe are still poorly known from a multidisciplinary point of view. This is the case of Les Échets (France), formerly investigated by pollen analysts. New coring operations carried out at Les Échets (Fig. 1), close to the sites that yielded the pollen sequences analyzed by de Beaulieu and Reille [2,3], provide the opportunity to reconstruct climatic and environmental changes for the last 140,000 years from several multiproxies (pollen, diatoms, ostracodes, Coleoptera, magnetism, tephra, organic sedimentology). The present study is part of this program and focuses on chironomid assemblages.
Geographic location of the Les Échets palaeolake.
Fig. 1. Localisation géographique du paléolac des Échets.
Chironomids (Insecta: Diptera) are one of the most diverse aquatic insect families, with about 650 species distributed in France [31]. Like Coleoptera [12], chironomids respond rapidly to environmental changes, as a result of their short generation times and the dispersive capacity of the winged adults. Moreover, chironomids are well preserved in Quaternary lake sediments [23], and are now acknowledged as an essential component in most multi-proxy palaeoenvironmental studies [7]. Subfossil chironomids are used to reconstruct: (i) palaeotemperatures [32]; (ii) lake eutrophication and nutrients assessing [33]; (iii) dissolved oxygen [42], and (iv) water-level changes [25].
Up to now, few chironomid analyses have been performed on the whole Eemian Interglacial. This paper provides the first continuous record of chironomid assemblages, covering the Last Interglacial (oxygen isotope stage OIS 5e) to the Early Würmian Pleniglacial (OIS 4). Our study is focused on two objectives: (i) to reconstruct past environmental and climate changes from chironomid assemblage dynamics, and (ii) to compare chironomid results with other Eemian palaeoecological data from mid-Europe. This preliminary study aims to disentangle the climatic signal from more local factors on chironomid assemblages, a necessary step for further quantitative temperature reconstruction based on chironomid–temperature inference models.
2 Materials and methods
A 10-cm-wide piston corer was used to extract twenty-nine 1.5-m-long, 43.84-m-spanning segments; 31 sediment samples weighing 150 to 500 g were collected from 39.1 m to 30.5 m, a section corresponding to the Last Interglacial and the lower part of the Würm Glacial (Fig. 2). The sediment below 39.1 m (upper part of the Riss) was sterile.

Relative abundances (%) of chironomid taxa versus depth (cm). Dating of the present sequence is made by comparison with the pollen stratigraphy from the same core (EC1).
Fig. 2. Diagramme des abondances relatives (%) des taxons de chironomes en fonction de la profondeur (cm). Le calage chronologique de la séquence analysée est effectué par comparaison avec la stratigraphie pollinique de la même carotte (EC1).
Extraction methods follow Gandouin et al. [14], identification methods follow Rossaro [46], Wiederholm [52], Schmid [48], Rieradevall and Brooks [44], and Brooks [6]. The relative abundance of each chironomid taxon for each sample is presented as a percentage diagram (Fig. 2). A stratigraphically constrained sum-of-squares cluster analysis (CONISS) [16] for percentage data was performed to highlight major changes in chironomid assemblage composition throughout the sequence. A correspondence analysis (CA) was performed on the n-by-p matrix of percentages to produce an ordination of the p taxa and of the n samples. Rare taxa were removed from the analysis. CA sample scores reflect the relative position of each sample (characterized by a specific faunal assemblage) among all samples in response to one or more environmental gradients. These scores give an estimate of the overall compositional change between two adjacent samples. The CA and its corresponding graphical plots (Fig. 3) were performed in ADE-4 [49].
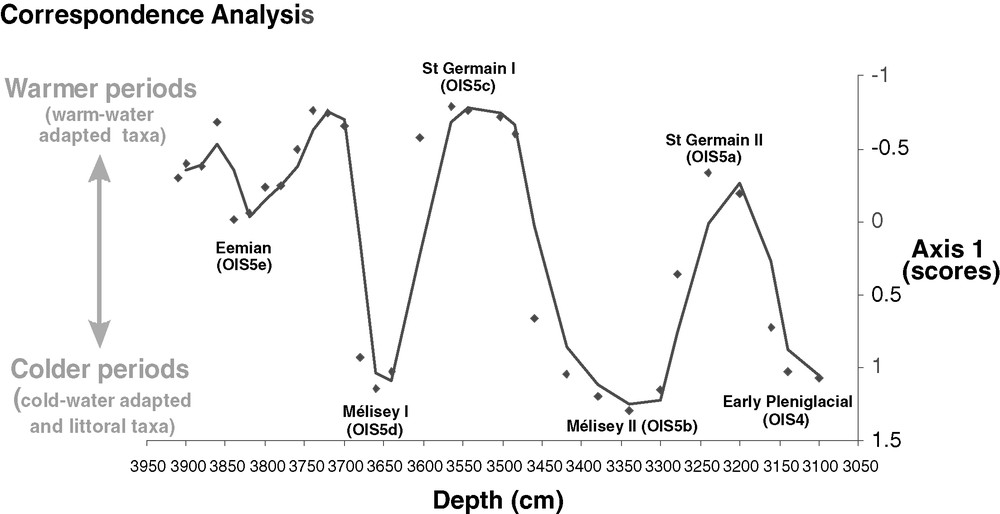
Evolution as a function of depth of the ordination scores on axis 1 of the correspondence analysis (eigenvalue = 0.52, n = 31 samples et p = 42 taxa; rare taxa have been downweighted).
Fig. 3. Évolution en fonction de la profondeur des scores sur l’axe 1 de l’analyse factorielle des correspondances (valeur propre = 0,52, n = 31 échantillons et p = 42 taxons ; les taxons rares ont été exclus de l’analyse).
3 Chironomid assemblages
A total of 42 taxa were identified in the 31 samples. A mean number of 92 head capsules per sample were counted, totalling 2841 head capsules. Eight biozones (Ech-ch1 to Ech-ch8) were identified by stratigraphically constrained cluster analysis (CONISS) (Fig. 2). The ecological classification of chironomid taxa (Fig. 2 and Appendix) and the CA (Fig. 3) allow us to establish that Ech-ch1, Ech-ch2, Ech-ch3, Ech-ch5 and Ech-ch7 are mainly characterised by deep-water, hypoxia-tolerant and warmwater genera (such as Chironomus), and that Ech-ch4, Ech-ch6 and Ech-ch8 are characterised by littoral and coldwater-adapted taxa (such as Microtendipes, Polypedilum and Tanytarsus lugens grp.).
In biozone Ech-ch1 (3910–3860 cm), the increase and the dominance of warmwater-adapted Chironomus, coupled with the decrease of the coldwater-adapted Tanytarsus lugens gp, could be explained by a rise in summer lake-water temperature. Moreover, it also suggests increased hypoxia [4] in the deeper part of the lake, induced by a rise in the water level and/or increased organic matter deposition [37]. However, waters must have remained sufficiently oxygenated to allow the persistence of T. lugens gp, which is known as an indicator of well-oxygenated conditions, and is dominant in oligo to mesotrophic lakes from the Swiss Alps [33]. The co-occurrence of T. lugens gp and Chironomus has been described by Brooks [6] in the Regenmoos Lake towards the end of the Younger Dryas. T. lugens gp usually occurs in deep waters of cold oligotrophic lakes. This could indicate cold profundal water during this period, which allows T. lugens gp to survive. The gradual increase in Chironomus may be a response to a progressive eutrophication of the lake throughout Ech-1.
The biozone Ech-ch2 (3840–3780 cm) is marked by a dramatic drop in the percentages of the profundal and hypoxia-tolerant Chironomus [19], coupled with the appearance and the increase of littoral taxa, such as Dicrotendipes, Polypedilum, Cladopelma lateralis types, and Cladotanytarsus mancus gp. This could be explained by (i) a lowering of the lake level: the reduced water column induces a better availability of the oxygen, which allows the littoral taxa to proliferate (littoral margins are also favourable to aquatic macrophyte development, which are suitable for colonisation by Dicrotendipes [22]); (ii) the fact that Chironomus is also known as a warmwater-adapted taxon, whose decrease could be explained by a lowering of summer temperatures. However, no coldwater-adapted taxon reappears over the considered period. It is therefore likely that in this case chironomids did not respond directly to a thermal climatic change, but rather to a decrease in water level.
In biozone Ech-ch3 (3760–3700 cm) again, chironomids provide evidence of a rise in water level, rather than of a climatic change. The deeper waters are marked by a strong hypoxia, as shown by Chironomus. At such a low oxygen concentration, the predator Procladius, which has a high oxy-regulatory capacity, is also able to survive [4].
The biozone Ech-ch4 (3680–3640 cm) is marked by a sharp drop in the percentages of the warmwater-adapted Chironomus, by the reappearance of coldwater-adapted taxa, such as Tanytarsus lugens gp [8,32] and Eukiefferiella devonica gp [47], and by the dominance of littoral and non-typical stenothermous taxa (such as Microtendipes). Such assemblages have been detected in cold phases of Late Glacial sequences from the Swiss Alps [6], and therefore could suggest a cooling of the climate. However, such indicators should be taken with caution, since such associations (with dominance of Microtendipes) may be related to climatic aridity [37], and probably also to low water levels, easily warmed up by daily insolation. Hence, bathymetric changes seem again to be a major factor controlling the distribution of chironomids at Les Échets. Significant percentages of psammophilous taxa, such as Cladotanytarsus mancus gp and Cladopelma lateralis type, indicate the presence of coarse sediment on submerged banks, induced by either erosion process, aeolian deposits, or local input of sediment by a tributary. The presence of Psectrocladius sordidellus-type may also be linked to a pH change [5]. P. sordidellus-type could also be associated with high levels in humic material [29], probably associated with a more open landscape in the catchment of the lake.
The biozone Ech-ch5 (3604–3484 cm) is marked by the complete dominance of Chironomus. Similar to the end of the Eemian (Ech-ch3), this dominance probably shows a rise in water level with strong hypoxia of the deeper waters. Increase in summer temperatures and enrichment in organic matter, which logically occur during this period of climatic warming, facilitate severe oxygen depletion in the water column.
The biozone Ech-ch6 (3460–3280 cm) is marked by the return of a fauna similar to that of Ech-ch4. However, the better representation of the coldwater-adapted Tanytarsus lugens gp at the beginning of this zone may indicate much cooler summer temperatures than during Ech-ch4. The high percentages of littoral taxa such as Microtendipes are characteristic of a lowering of the water level, resulting probably from lower precipitation. The strong increase of Polypedilum throughout Ech-ch6 could imply a progressive enrichment in nutrients and organic matter in the lake, but the water remained probably sufficiently oxygenated to fulfil the ecological requirements of this taxon [37]. The Polypedilum increase could be induced too by warmer summer water temperature, which may be furthered by probable climate improvement at the end of the zone. Conversely, Polypedilum are also often associated with submerged macrophytes [9], which may be present in cool conditions.
In biozone Ech-ch7 (3240–3200 cm), faunal assemblages differ markedly from those of Ech-ch5, because of a lower abundance of the dominant Chironomus taxon and a significant representation of Tanytarsini (Tanytarsini unidentified + coldwater-adapted Tanytarsus lugens gp). This could be explained by more oxygenated water during Ech-ch7, and probable lower summer temperatures.
The biozone Ech-ch8 (3160–3100 cm) is marked by the strong dominance of Tanytarsini, Tanytarsus, and the coldwater-adapted Tanytarsus lugens gp. Although Tanytarsini and Tanytarsus include eurytopic species, they are often associated with oligotrophic, cold subarctic, and Arctic-Alpine lakes [24], and suggest Pleniglacial cold climate conditions. The abundance of littoral chironomid taxa, such as Cladotanytarsus mancus gp, during the Early Pleniglacial (OIS 4) could be easily explained by a fall in the lake level induced by climate aridity, the latter resulting from the extension of the European ice sheets [34].
4 Comparison with other mid-European palaeoenvironmental and palaeoclimatical data
At the beginning of the sequence, the warming inferred from chironomid assemblages is in agreement with the hypothesis of an Early Eemian thermal optimum, as suggested by pollen data [11,18]. Based on Coleopteran data from la Grande Pile [41], summer temperatures range from 9 to 23 °C at the onset of the Eemian, then range from 14 to 24 °C until the end of the Eemian, which is also consistent with the dominance of Chironomus at Les Échets, a species-rich genus that can be distributed along a temperature gradient from 8° to 19 °C in the Swiss Alps [21] and probably higher than 14 °C, according to a dataset from northern Sweden [30]. Moreover, the increase in lake level may be correlated with high regional rates of precipitation [11] (Fig. 4), also evidenced in Massif Central [45] and in the northern Alps [27].

Comparison between the chironomid biozones from Les Échets with (A) terrestrial climatostratigraphy units from western Europe [18] and several pollen sequences from France: (B) Ribains [45], (C) Les Échets [11], and (D) in the Northern Alps [27]. The shaded areas correspond to low lake-water levels identified from chironomid assemblages.
Fig. 4. Comparaison entre les biozones de chironomides des Échets avec (A) les unités climatostratigraphiques pour l’Europe de l’Ouest [18] et des séquences polliniques françaises : (B) Ribains [45], (C) Les Échets [11] et (D) Alpes du Nord [27]. Les zones en grisé correspondent à de bas niveaux lacustres indiqués par les assemblages de chironomes.
In the second part of the Eemian (Ech-ch2, Ech-ch3), the thermal climatic signal is difficult to detect, since chironomid assemblages seem to be more controlled by lake water levels than by summer thermal changes. Cheddadi et al. [11], Rioual et al. [45], and Klotz et al. [27] describe a decrease in precipitation in the middle part of the Eemian period, which could be correlated with the low water level suggested by chironomids at Les Échets (Ech-ch2).
In the Massif Central and the Alps, climate conditions became colder from the middle part of the Eemian [11,27,45], and a moderate climate oscillation influencing only winter temperatures is possible, whereas oxygen isotope data from the GRIP ice core [17] suggested a major mid-Eemian cooling. Unfortunately, the chironomids are only slightly influenced by winter temperature, since their climate-sensitive stage is in spring and summer, periods of emergence and reproduction of the imago [1]. Nevertheless, the absence of coldwater-adapted chironomids seems to be inconsistent with a major mid-Eemian cooling.
During the Mélisey I, the decrease in regional precipitation suggested by a low water level and littoral chironomids is confirmed by pollen data on French and Swiss sites [27,28] and over western Europe [18,45]. Concerning summer temperatures, chironomids have clearly responded to an important cooling, already reconstructed in central Europe from pollen data [28], with a decrease in summer temperature to 14 °C. This cooling was not recorded at Ribains [45] and Grande Pile [41], due to the absence of modern pollen analogues and of really cold-adapted Coleopteran assemblages, respectively.
The climatic improvement of the St Germain I interstadial is characterized by chironomid assemblages comparable to those obtained at the end of the Eemian. This is in agreement with the value of 18.5 °C reconstructed from pollen at Les Échets [28] and with temperatures between 13–23 °C reconstructed from Coleoptera at Grande Pile [41]. The precipitation rise detected by chironomids is also consistent with other regional data [27,28].
During the Mélisey II stadial, the climate cooling is clearly revealed by the chironomid assemblages. Moreover, the better representation of the cold-adapted Tanytarsus lugens gp suggests that summer temperatures were lower than during Mélisey I. This disagrees with the pollen-based temperature reconstruction [28] suggesting summer temperatures probably 1 °C higher in Mélisey II than in Mélisey I. Chironomid temperature inference models and multidisciplinary studies are needed to resolve this discrepancy. It is also possible that the better representation of coldwater-adapted taxa could have been caused by an increase in continentality [28] marked by late springs.
The climatic improvement of the St Germain II interstadial is clearly marked by the dominance of Chironomus. At Les Échets and other mid-European sites, pollen data suggest that summer temperatures were respectively around 19.7 °C, and 16–17.7 °C [28]. At Grande Pile, summer temperatures reconstructed from Coleoptera range between 15 and 19 °C [41].
In northwestern Europe, thermal conditions during the St Germain II are still questionable. Up to now, summer temperatures during this interstadial were known to be similar to those prevailing during the St Germain I [50]. Recent pollen data from Central Europe [28] suggest that the summer optimum in St Germain II was about 0.7 to 1.2 °C warmer than the optimum in a more continental St Germain I, whereas chironomids suggest the contrary (Fig. 3), with summer temperatures in St Germain II colder than in St Germain I. This latter hypothesis could explain the lower percentages in thermophilous forest (Quercus and Carpinus) and the reestablishement of conifer forest with Picea and Pinus observed in the French sequences at the end of the interstadial [18].
Coldwater-adapted chironomids from Early Pleniglacial are consistent with pollen reconstructed temperatures ranging between 9 and 15 °C [28]. These figures are also similar with those reconstructed from Coleopteran data at Grande Pile, estimated to 10–13 °C [41], and temperatures of around 10 °C reconstructed for northwestern Europe [26,50].
5 Conclusion
Past lake-level changes and summer temperatures from the Early Eemian (OIS 5e) to the Early Pleniglacial (OIS 4) at Les Échets have been reconstructed from subfossil Chironomids. The lake-level fluctuations have been correlated with global humidity patterns throughout mid-Europe. Hence, the role of lake depth fluctuations on assemblages must be taken into consideration in future high-resolution quantitative temperature reconstructions. This improved resolution is now required to identify short and abrupt climatic events, such as Montaigu event [28,41,45]. Further quantitative temperature reconstructions inferred from chironomids should also focus on ecological and thermal differences within both stadial (OIS 5d and OIS 5b) and interstadial (OIS 5c and OIS 5a) periods. However, this original and independent approach confirms that subfossil chironomids already represent an effective palaeoecological and palaeoclimatical tool.
Acknowledgements
The study is part of a large multiproxy palaeoecological program supported by ANDRA, CNRS, NSF, the Lund University and IMEP.
Appendix
Ecological classification of chironomid taxa from Les Échets according to published data. * Einfeldia could be also found in cold Swiss Alps: it has been described at Leysin (Canton of Vaud) during the Younger Dryas [6] as a littoral taxon living in the soft sediment tending towards the eutrophic. ** Stictochironomus is also a cold taxon in the Swiss Alps [20] and can be found in profundal soft sediments or littoral sandy sediments in mesotrophic and oligotrophic lakes [53].
Classification écologique des principaux taxons de chironomides des Échets selon les travaux scientifiques publiés. * Einfeldia peut aussi coloniser les lacs alpins suisses : il a été décrit lors du Dryas récent à Leysin (canton de Vaud) [6] comme un taxon littoral vivant dans les sédiments meubles tendant vers l’eutrophisation. ** Stictochironomus est aussi un taxon des Alpes suisses [20] et peut être trouvé dans les sédiments meubles profonds ou les zones sableuses littorales des lacs oligotrophes à mesotrophes [53].
| Ecological categories | References | Ecological categories | References |
| Warmwater-adapted taxa | Warmwater-adapted and deep-water taxa | ||
| * Einfeldia sp | [51] | Chironomus spp | [6,32,51] |
| Fleuria sp | [52] | ||
| Glyptotendipes spp | [32] | ||
| Synorthocladiussemivirens | [47] | ||
| Coldwater-adapted taxa | Coldwater-adapted and rheophilous taxa | ||
| Tanytarsuslugens gp | [32] | Eukiefferiella spp | [32,52] |
| Krenosmittiaboreoalpina | [10] | Eukiefferielladevonica gp | [47] |
| Eukiefferiellaclaripennis gp | [15] | ||
| Littoral taxa | Littoral and psammophilous taxa | ||
| Dicrotendipes spp | [22,51] | Cladopelmalateralis type | [39] |
| Polypedilum spp and Polypedilum pedestre type | [36,51] | Paratendipesalbimanus | [51] |
| Endochironomus sp | [51] | Pseudochironomusprasinatus | [37] |
| Corynoneuralacustris type | [6,38] | **Stictochironomus sp | [40] |
| Cricotopus/Orthocladius/Paratrichocladius | [20] | Cladotanytarsusmancus gp | [13,36] |
| Microtendipes spp | [6] | ||
| Cryptochironomus sp | [35,51] | ||
| Tanytarsuspallidicornis gp | [43] |