

1 Introduction
The studies based on recent data consider in priority the spatial dimension, the temporal dimension being supposed uniform. The well-known example of the relationship prey–predatory where the population of the prey declines when the predator thrives, then the reverse, according to the classical law of the population dynamics (equations of Lotka–Volterra) supposes that there is no temporal disturbance of climatic type. The paleoenvironmental studies show that, on the contrary, temporal dimension has its own texture, with successions of ice ages and interglacials. Consequently, during a long climatically stable period, even if the competition between species ends up modelling the landscape, this stability is of sufficiently short duration to question periodically equilibria.
Before three millions of years, the studies of marine [23] and terrestrial sediments [9] converge to describe a world with rare glaciations, even if the fluctuations of the Earth’s orbit around the Sun [2] have induced periodical variations in the quantity of energy that our planet has received from the Sun. After 2.6 Myr, these periodical fluctuations intensified and caused the glaciation occurrences that are the major fact of the Quaternary period. These alternations are explained by the theory of Milankovitch, who identified several major cycles in the insolation and then in the climate variations. They are roughly 400, 100, 40, and 20 thousand years [2]. These glaciations left traces in the geological landscapes (moraines), profiles and sedimentary accumulations that many tools made it possible to decode. We will focus on two bioindicators having each one their particular temporal characteristics: pollen and tree rings.
The pollen grains, which are dispersed at more or less long distance according to their morphology and their size (between 5 and 100 m) are an essential part of the reproduction of the higher plants. Their envelope (exine) is made up of sporopollinine, which is a very resistant substance. When they are deposited and accumulated in the sediment, exine is preserved almost indefinitely if it is safe from oxidation. Starting from a sample taken in a sedimentary core, the palynologist identifies the transmitting plants source, making it possible to reconstruct the composition of the surrounding vegetation. We ignore here the problems related to pollen preservation and disturbance of the sediment, and we compare the past data to our knowledge of the present vegetation to imagine which could be the climate having generated these vegetation types according to the (uniformatarian) principle that the same causes induce the same effects [11]. This is not without trap, as it will be seen below.
The tree rings relate to one more recent period, generally the last centuries. The cores taken in the tree stem give time series of annual growth mainly driven by the climate: a thick ring is produced by one climatically favourable year. The notion of favourable must be understood in term of temperature for trees growing in a generally cold climate, and in term of water availability for trees growing in a generally dry climate [5]. Further developments made it possible to analyze the evolution of the tree-ring density within the year, giving access to the seasonal variations [18], and the isotopic composition of the ring, making it possible to define better the climatic signal [12].
Decoding the biological relationships between bioindicators and climate is often based on statistical equations, which allows an quantitative approach, but often neglects robust causal analysis. Conceptual models make it possible to describe in a simplified way complex relationships [10]: they contribute to the understanding of processes, but really do not allow one to predict or analyze tree sensitivity to future pertubations, such as global warming. The approaches of the biogeographic and biogeochemical modelling make it possible to explain variations at a landscape scale by studying processes at the organismal level. These models, although still relatively simplified, allow a predictive approach. They also make it possible to transpose our observations of the past in terms of risks for the future.
2 Conceptual model of species response to environment
The niche theory [1] makes it possible to describe the relationships between the species and their environment. According to [10], one can represent in any environmental space – to simplify, a climatic space with two dimensions – the distribution eras of the species considered (Fig. 1). The potential environment space occupies the whole space described by these two variables, but only the era called realised environmental space really exists in the real world. A given species can survive in a niche defined by the constraints that it is able to support. It is its fundamental niche, but only the intersection between this fundamental niche and the realised environmental space exists in the real world: it is the potential niche. For reasons external to the environmental space considered, for example additional constraints due to the edaphic conditions, this species will not occupy all the fundamental niche, but only one part, called realised niche. This model assumes implicitly that species fitness to its niche is maximised at the center of the niche and falls toward the edges. Some of the difficulties that one can meet in climate reconstruction from bioindicators is that the tolerance of a species to a given climatic range can be estimated only by its realised niche. One may underestimate its plasticity and then underestimate the associated climatic range.
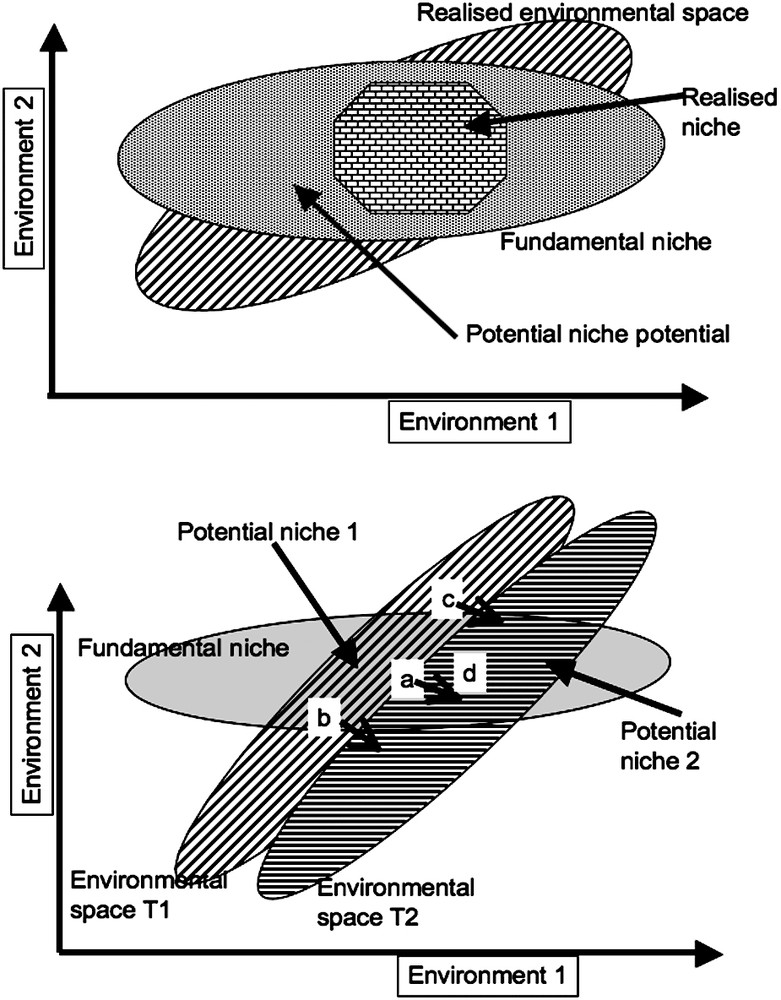
Conceptual model of the interaction between species and environment according to [1,10], see text for explanations.
Modèle conceptuel des interactions entre espèces et environnement selon [1,10], voir texte pour les explications.
The second part of Fig. 1 makes it possible to understand how a species can evolve when the climate changes (T1 changes to T2). Let us assume the environmental position of the population (a) of the given species, well in the center of its fundamental niche. When the climate changes from T1 to T2, it remains around the center of its fundamental niche, and this population is maintained. Because of the change, population (b) is pushed to leave its fundamental niche and thus will die out. Population (c), in environmental space T1, did not exist because it was outside its fundamental niche. The climatic change makes that it is back to its fundamental niche and makes possible colonisation (if other conditions, not taken into account by the model, allow it). Let us consider population (d): it occupies the margin of environmental space T1, which constraints it to conditions of extreme survival, but if the conditions suddenly changed to T2, then this population will be able to bloom: in T1, it will be very sensitive to the climate, but much less in T2. Climatic changes may make it possible that extreme events become normality. The variability is then as much important than mean changes. Another point that is not easily taken into account by this model is the rate of change. If the change is much quicker than the response time of the population, there is little chance that a new equilibrium could be reached. It is a point where past is not completely a key for the future, as the present anthropogenic changes are much quicker than the glacial–interglacial transition.
3 Statistical approach
The first methods of climate reconstructions, devised several decades ago, were directly based on the niche theory [4]. Using floristic atlases and field samples, potential niches of significant species have been defined in the space of two climatic variables (often the summer and winter temperatures). The overlapping of the niches of the species present in a given assemblage provides the desired climatic limits. As the pollen data provide more precise information than the simple presence/absence of species, namely their abundance, the method has evolved to compare past data abundances with those of surface samples. Two samples that are similar are supposed to come from the same vegetation and thus from the same climate. They are called ‘analogues’. The method of the analogues was abundantly used to reconstruct climate over several hundreds of thousands of years [3,7].
The statistical approaches have given very good results, but provided that the past assemblages can have analogues today. Possibly, two species, which have their potential niche overlapping in the realised environmental space, may disjoin under a rather important climatic change. So it will make it impossible to find any modern analogue containing these two species. Another problem is that an external factor may interfere, making the realised niche rather reduced as compared to its potential niche. One will see below an example with the effect of the atmospheric CO2.
4 An extremely cold event: the last glacial maximum (LGM)
Twenty-one thousands years ago, Europe was covered with a herbaceous vegetation instead of the temperate forests that one would see everywhere nowadays if the man had not cleared abundantly. The composition of the pollen assemblages was relatively poor, with a predominance of Poaceae and Artemisia. The modern analogues of this type of vegetation are partially found in the continental steppes (where Artemisia dominates) and partially in the Arctic tundra, where Poaceae dominate. Whereas in the first, the winter temperatures are very low and the summer temperatures relatively high, in the second, summer temperatures are rather low. The seasonal variation of the monthly temperature profile was then different. The duration of the day plays also an important role for Arctic vegetation. From these data, Peyron et al. [15] and Tarasov et al. [20] reconstructed rather cold winters, but with large uncertainties due to the large extent of the potential niches of some species present during this period marked by a strong glacial advance. Even if these inferences agree rather well with the temperature reconstruction in relation to the extension of permafrost [17], it is not certain whether the values reconstructed for sites located close to the Mediterranean Sea were compatible with a sea ice-free all the year or not. Briefly said, is it possible that the realised niche of the taxa concerned is drawn towards cold by other factors?
The first assumption tested is that the increase of climate variability around an average less cold than that inferred by Peyron et al. [15] could explain the disappearance of the forest. Indeed, a larger variability increases the amplitude and frequency of the extremes. The extreme cold events being compensated by extreme warm ones, this would not affect the climate average, but it modifies the vegetation, which could not support an excessive repetition of these cold waves. We consider here the change in high-frequency variance and ignore possible changes in the distribution of rare events. Testing this assumption can be done with a dynamic vegetation model: model LPJ-GUESS [19]. Such a model simulates some characteristics of the vegetation from climatic variables, soil characteristics, and atmospheric concentration of CO2. Several climatic scenarios, reflecting the various assumptions to be tested, are introduced into the model: more or less cold and/or wet climates with different variances [16]. The vegetation simulated for each one of these scenarios was compared with the pollen data. The conclusion was that, to simulate an herbaceous vegetation, it was necessary to lower the temperature and precipitation to levels close to those reconstructed from pollen data, and that the increase in variability alone was not sufficient to explain the disappearance of forest.
The second assumption tested is that the LGM lowering of the atmospheric CO2 to 200 ppmv, whereas presently it is higher than 360 ppmv, was a sufficiently limiting factor to explain the disappearance of the forests. Wu et al. [22] used another model, simpler than LPJ-GUESS, BIOME4, together with pollen data. By a procedure of inversion too complicated to be explained here (see [8,22]), they showed (Fig. 2) that the CO2 could explain the equivalent of a reduction of 10 C of MTCO in western Europe, of 7 C in the Mediterranean area, but no skew is established for the eastern areas. In the same way, one observes an equivalent gradient of systematic skew for annual precipitation, with a maximum of almost 500 mm. The explanation is that, in a low-CO2 environment, the trees are obliged to open their stomata to absorb a sufficient quantity of carbon, leading to a greater water loss by evapotranspiration. The subsequent water stress does not allow forests to maintain. Current abundance in CO2 shifts the realised niche of the LGM vegetation towards a cold and dry continental steppe. The potential niche of the LGM European vegetation was only at the edge of its modern equivalent.

Reconstruction of the mean temperature of the coldest month (MTCO) and of the annual precipitation in Europe and Siberia for the last glacial maximum using two methods: a statistical method (PFT) and an inverse vegetation modeling method using BIOME4 with the LGM level of CO2 (IVM). The reconstructions are grouped into boxes and whiskers representing interquartiles and 90% confidence intervals, calculated on all the sites of the considered region [22].
Reconstruction de la température du mois le plus froid (MTCO) et de la précipitation annuelle en Europe et Sibérie pour le dernier maximum glaciaire à partir de deux méthodes : une méthode statistique (PFT) et une méthode d’inversion de modèle de végétation BIOME4 avec le niveau de CO2 du dernier maximum glaciaire (IVM). Les reconstructions sont groupées en boîtes à moustaches représentant l’intervalle interquartile et l’intervalle de confiance à 90.
5 Droughts in the Mediterranean region and tree-growth response to future changes
The Mediterranean forests are crucial to preserve the high biodiversity that characterizes them and to provide essential ecosystemic services, such as the protection of the soils, the conservation of the water resources and the regulation of the climate. The climatic models simulate, for the 21st century, a significant warming and a decrease of precipitation, with a significant increase of extreme events, which can reduce considerably the productivity of these forests. It is interesting to look in the recent past how those responded to the climate variations. Dendrochronology makes that possible. A set of series of Pinus halepensis annual growth are available for southeastern France [14], for at least one century. Model MAIDEN simulations fit relatively well these growth series [13]. The correlation between simulated series and the average series (Fig. 3) is largely significant () over the period 1963–1964 [6]. Other tests showed that the spatial distribution of the growth across southeastern France was also correctly simulated.
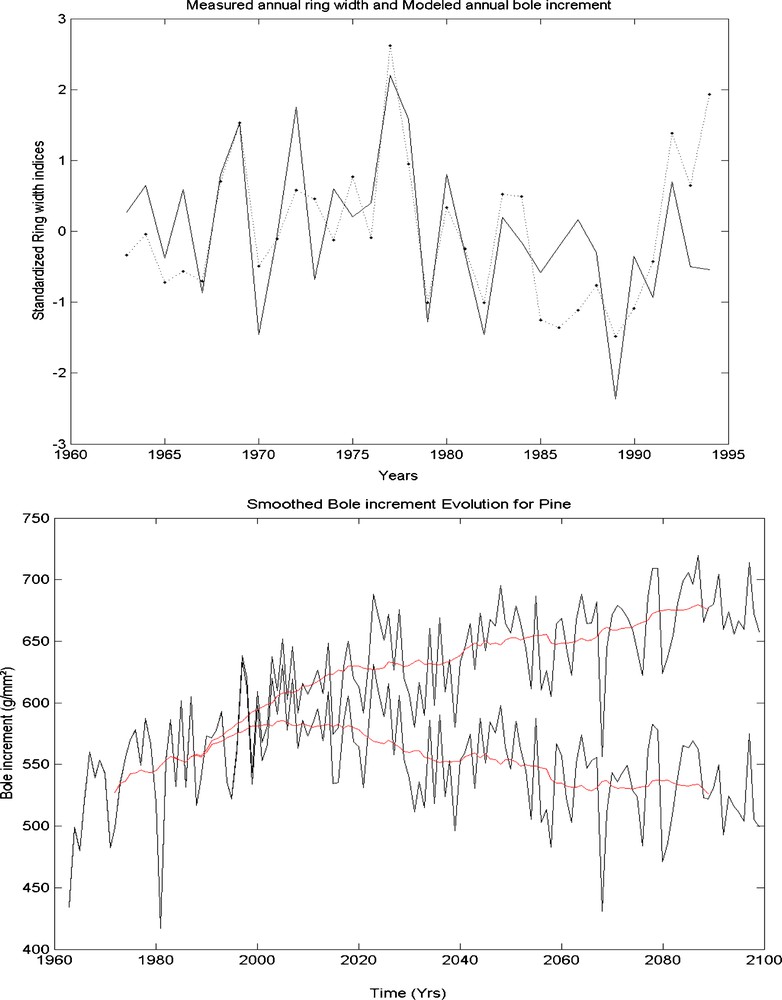
Top: simulation of tree growth for the period 1963–1994 using MAIDEN and comparison with the mean indexed chronology of Aleppo pine of the southeastern France. Bottom: simulation of pine growth for the next century using a climatic scenario of ARPEGE, taking into account the CO2 fertilisation effect (lower curve) or not (upper curve); annual variations are accompanied with moving average.
Partie supérieure: simulation de la croissance des arbres pour la période 1963–1994 à partir du modèle MAIDEN et comparaison avec la chronologie moyenne indicée du pin d’Alep du Sud-Est de la France. Partie inférieure: simulation de la croissance du pin pour le XXIesiècle à partir d’un scénario climatique d’ARPEGE, prenant en compte l’effet de fertilisation du CO2 (courbe supérieure) ou non (courbe inférieure) ; les variations annuelles sont accompagnées d’une courbe lissée par moyenne mobile.
One thus could use a climatic scenario of the model ARPEGE developed by Météo-France as input climate for MAIDEN and to simulate the pine growth until the end of the 21st century (Fig. 3). According to whether one takes into account the climatic change alone or the climatic change together with the fertlisation effect of CO2, one finds a different evolution. In the first case, the growth decreases after the beginning of the 21st century and in the second case, it continues to slightly increase to stabilize at the end of the 21st century. The water-use efficiency is in fact increased by the stomata closing due to more abundant carbon in the atmosphere (the opposite effect than for the LGM). This effect is sufficient to counterbalance the lower water availability induced by precipitation decrease and evaporation increase.
This approach does not take into account the fact that frequent extreme events may damage the trees, which will not be able to recover between two events. So the productivity may decrease rapidly and balance the fertilization effect [21]. These effects on the sanitary state of the tree cannot yet be modelled with mechanistic models. Moreover, this model does not take into account anymore the possible limitations due to lack of nutriments. Here again, we see that if we do not take into account the CO2 concentration, climatic change pushes quickly the trees to the margin of their potential niche, whereas the CO2 fertilisation effect keep them within the niche during several decades.
6 Conclusion
Even if the conceptual and mechanistic models are still simplistic, the most important physiological processes are taken into account and some indications on the population vulnerability can be drawn. To avoid overinterpretation, limits must be clearly stated. In that respect, the use of past data (on several decades or centuries for tree rings; on several millennia for pollen data) increases the chance to have a better insight into the potential niche of the species: larger is the distribution gradient of the species, larger is the chance that the realised niche tends to the potential niche. In other words, past data give us the possibility to calibrate some characteristics of the model used on a larger range of situations and so to avoid some spurious extrapolations.
Interaction between models and data has another power. Interpretation of the data is necessarily based on the knowledge that we have of the modern world. The changes observed in the vegetation data are interpreted as first-order changes in dominant factors (climate or anthropogenic). Second-order changes (or changes in variance) are more difficult to handle. The data–model comparison leads us to consider alternative explanations when there are discrepancies between both approaches. Moreover, models give us tools to test these alternative hypotheses. From this fruitful interaction, we expect a better knowledge of more or less hidden processes and finally a better chance to anticipate future changes.
Acknowledgements
This research has been partly funded by the French GICC-2 program (REFORME), by the European Union project MOTIF, by the European Science Foundation, EUROCLIMATE/DECVEG and the French ANR PICC.