

1 Introduction
Le peuplier noir (Populus nigra L.), espèce dioïque, est présente dans pratiquement toutes les vallées des régions circumméditerranéennes. Son aire s’étend du Royaume-Uni jusqu’au centre de l’Asie 〚1〛. Espèce pionnière, P. nigra profite des perturbations cycliques des rivières et fleuves pour s’implanter sur des sites neufs et transitoires. Il cède par la suite sa place à des espèces de bois durs, dans le cadre de successions végétales 〚2–4〛. Ces rajeunissements périodiques des milieux riverains confèrent aux ripisylves leur extraordinaire diversité biologique et favorisent le développement d’une mosaïque forestière où se juxtaposent des communautés d’âges variés 〚5–7〛. Aujourd’hui, par suite de fortes perturbations anthropiques dans les milieux riverains, on assiste à la disparition de nombreux sites de recrutement et P. nigra se retrouve à la limite de l’extinction dans de nombreux endroits en Europe de l’Ouest 〚8〛, en particulier en Belgique et au Royaume-Uni 〚9, 10〛. Une seconde menace pèse sur P. nigra : un appauvrissement de son stock génétique sous la pression de clones plantés devenus prédominants dans de nombreuses plaines alluviales. Ces menaces ont justifié le lancement d’un programme pour la conservation des ressources génétiques du peuplier noir en Europe 〚11–13〛. En France, les travaux ont porté essentiellement sur les bassins de la Loire et du Rhône 〚14〛, mais aucune étude n’a encore été faite sur la population garonnaise.
Face à ces enjeux se pose donc aujourd’hui la question de la place du peuplier noir dans une politique de gestion et d’aménagement des milieux riverains, sachant que la structure et le fonctionnement actuel des zones riveraines dépendent souvent d’événements historiques passés, dont les effets se prolongent dans le présent 〚15〛. Ce legs du passé recouvre des événements naturels et anthropiques, tels que déboisements, endiguements, introduction de clones ou d’espèces, aménagements divers. Dans le cas de la Garonne, il est admis que la surface occupée par la ripisylve a fortement diminué dans le passé 〚16〛, mais l’ampleur de cette régression et sa localisation dans le temps n’a encore jamais été précisée, si ce n’est sur quelques sites isolés 〚17〛. À l’amont de la confluence du Tarn, les boisements riverains de la Garonne occupent aujourd’hui une largeur moyenne d’environ 530 m sur l’ensemble des deux rives, ce qui correspond à un taux de boisement de 17,3% dans la plaine inondable 〚18〛. Toutefois, près de 90% de ces boisements sont des peupliers plantés, dont les trois quarts sont dominés par les deux clones hybrides Populus x euramericana (Dode) Guinier cv. « I214 » et cv. « I45/51 ». Sur les rives de la Garonne, cette habitude de cloner par bouturage les individus présentant un port intéressant s’est répandue très tôt pour stabiliser les berges, pour gagner du terrain sur le fleuve ou simplement pour produire du bois 〚19〛. Mais les clones cultivés aujourd’hui ne sont plus les mêmes que dans le passé, et leur impact éventuel sur la population locale de peuplier noir a certainement changé au fil du temps. Il est difficile d’obtenir des indications sur les surfaces occupées respectivement par les plantations de peuplier et par la ripisylve, car les premières statistiques agricoles et forestières publiées, au cours du XIXe siècle, à l’échelle des départements ne faisaient pas cette distinction. Cependant, cette information peut être extraite de certaines cartes historiques.
Notre premier objectif a donc été de vérifier si les cartes historiques existantes contenaient une information permettant de reconstituer l’évolution des boisements riverains le long de la Garonne au cours des deux derniers siècles. Pour chaque carte, l’enjeu était d’extraire une information quantitative sur les surfaces occupées respectivement par les peupliers plantés et par la ripisylve. Notre second objectif a été d’évaluer, sur la même période historique, l’impact des deux principales perturbations qui ont pesé sur la population garonnaise de peuplier noir, à savoir la présence massive de clones cultivés et les aménagements du fleuve. Dans le premier cas, il s’agissait d’identifier les principaux clones cultivés en vallée de la Garonne et de préciser leurs caractéristiques, afin d’estimer les risques d’une pollution génétique éventuelle. Dans le second cas, il s’agissait d’estimer les conséquences des modifications de la dynamique fluviale et des conditions hydrologiques du milieu sur la population garonnaise de peupliers noirs, compte tenu des facteurs favorables à leur régénération 〚20〛.
2 Matériels et méthodes
Notre travail s’est concentré, dans la moyenne vallée de la Garonne, entre Toulouse et la confluence du Tarn. Cette section est caractérisée par une plaine inondable de 3 km de large et par de larges méandres. Elle a été qualifiée de « Garonne débordante » par opposition à la section en amont de Toulouse où la plaine est très étroite, encaissée et peu sujette aux submersions, et aussi par opposition à l’aval de la confluence du Tarn, où le lit du fleuve est fortement encaissé, et parfois même endigué dans sa propre plaine 〚21〛. Sur le plan climatique, cette section correspond à la partie la plus sèche du bassin, avec un déficit hydrique prononcé, qui s’étend chaque année de juillet à septembre. La Garonne, au régime pluvio-nival, se singularise par un étiage sévère qui survient en pleine sécheresse climatique estivale. À Toulouse, le module est 200 m3 s–1, le débit objectif d’étiage 52 m3 s–1 et la crue centennale proche de 6500 m3 s–1.
L’évolution des boisements riverains a été suivie sur une période de deux siècles, grâce à six séries de cartes historiques, trouvées dans divers lieux d’archives, et dont la partie commune couvrait une section de 42 km entre Grenade (Haute-Garonne) et Moissac (Tarn-et-Garonne) (Tableau 1). Ces cartes ont été copiées sur les lieux d’archivage, scannées au laboratoire au format A4, puis assemblées dans une base raster géoréférencée (système Lambert III, pixel 5 m × 5 m) à l’aide du logiciel Easi/Pace (PCI-Geomatics, Canada). Dans chaque carte, les boisements spontanés, les plantations de peuplier et le lit mineur ont été identifiés et vectorisés. Dans certaines cartes (1810, 1860 et 1945), les dépôts alluvionnaires le long du fleuve (îles, bancs de sable ou de galets, berges sans végétation) étaient mentionnés. Ils ont également été vectorisés, car ils constituaient des sites potentiels de recrutement de P. nigra. Les informations de la carte de 1980 ont dû être complétées par celles des photos aériennes panchromatiques de 1979. Il convient néanmoins d’analyser et de comparer ces cartes avec une certaine prudence, étant donné la disparité des échelles d’origine et des objectifs qui ont prévalu lors de leur élaboration. Pour lever les incertitudes dues à l’absence de légendes à certaines date, d’autres documents d’archives ont été consultés, en particulier des cartes réalisées à peu d’années d’intervalle sur des secteurs plus restreints. D’autres documents issus des archives nationales, des archives départementales de Gironde, de Tarn-et-Garonne et de Garonne, ainsi que des archives communales de Castelsarrasin, de Moissac et de Verdun-sur-Garonne, ont fourni une information sur les aménagements du fleuve, la gestion locale des boisements, la réglementation en vigueur et les conflits entre riverains et l’administration. Les archives des Eaux et Forêts à Nancy sont également riches en information sur la culture du peuplier en vallée de la Garonne au cours du XXe siècle.
Liste des cartes utilisées pour l’étude historique des boisements de la Garonne entre Toulouse et la confluence du Tarn.
| Type de carte | Année | Échelle | Localisation |
| Carte du cours de la Garonne | 1810 | 1:5000 | Archives départementales de la Gironde |
| Minutes de la carte d’état-major | 1849 | 1:40 000 | Cartothèque de l’Institut géographique national |
| Carte du cours de la Garonne | 1868 | 1:10 000 | Archives départementales de la Haute-Garonne |
| Carte d’état-major type 1889 | 1908 | 1:50 000 | Cartothèque de l’Institut géographique national |
| Carte IGN | 1945 | 1:20 000 | Cartothèque de l’Institut géographique national |
| Carte IGN « série bleue » | 1980 | 1:25 000 | disponible dans le commerce |
3 Résultats
3.1 Évolution des surfaces boisées
Au Moyen Âge, les berges étaient abondamment plantées de petits ormeaux et toute la région de la moyenne Garonne était alors vouée à l’orme, les vallées comme les coteaux 〚19, 22〛. Le peuplier noir, parfois appelé localement « sarrazin », existait à l’état spontané dans les boisements riverains de la Garonne, mais il est peu mentionné, car son bois était peu apprécié 〚23, 24〛. Au XVIIe siècle, pour faciliter la remontée des bateaux, de Bordeaux à Toulouse, les riverains devaient supprimer sur les berges du fleuve tout arbre susceptible de gêner le halage, qui se faisait alors par traction humaine. La largeur minimum du chemin de halage fut fixée à 25 pieds 〚25〛. Colbert améliora les conditions de la navigation sur la Garonne et assura une continuité entre la Méditerranée et Bordeaux avec la construction du canal du Midi, qui relia dès 1681 la Méditerranée à Toulouse. Il créa un corps d’inspecteurs de rivières qui signalèrent souvent le manque d’entretien des berges 〚26〛. La Révolution et les guerres napoléoniennes provoquèrent une baisse des échanges et les consignes d’entretien ont été peu suivies. À la même époque, les ormes ont subi un dépérissement sévère par suite de l’attaque d’insectes 〚19, 27〛. Ce déclin des ormes a sans doute favorisé l’essor des plantations de peupliers.
La première carte historique qui mentionne les boisements riverains date de 1810. La largeur moyenne boisée sur l’ensemble des deux rives correspondait alors a un ruban boisé de 608 m (Figs. 1 et 2). La moitié de ces boisements était occupée par la ripisylve, qui formait un cordon continu le long du fleuve. L’autre moitié était occupée par des plantations de peupliers, situées généralement dans la plaine inondable à l’arrière de ces boisements spontanés. La présence massive de peupliers plantés dans la vallée de la Garonne est donc nettement antérieure à la Révolution française. Les clones dominants étaient alors des peupliers noirs locaux, le peuplier d’Italie et le peuplier de Caroline 〚19〛. Cependant, il n’est pas possible de préciser l’importance relative de chacune de ces catégories.

Évolution des boisements riverains de la Garonne entre 1810 et 1868, sur une section de 42 km à l’amont de la confluence du Tarn.

Évolution de la largeur moyenne des boisements riverains de la Garonne sur une section de 42 km, à l’amont de la confluence du Tarn.
Vers 1849, les officiers topographes ont réalisé au 1:40 000 les minutes de la carte d’état-major sur la vallée de la Garonne, en faisant une distinction entre les boisements spontanés et les plantations. Cette distinction disparaît dans la carte d’état-major, publiée au 1:80 000. En 1849, la largeur moyenne boisée n’avait pratiquement pas changé par rapport à 1810. Toutefois, au cours de cette période, les boisements spontanés ont fortement diminué (–51%) alors que les plantations de peupliers étaient en plein essor (+40%) et occupaient 77% des surfaces boisées, contre 54% auparavant. Cette diminution des boisements spontanés peut s’expliquer par les travaux qui ont été menés vers 1830 dans le lit mineur, sur toute la ligne navigable de Bordeaux à Toulouse : les chemins de halage ont été réparés, les écueils ont disparu et la navigation à l’étiage a été améliorée 〚28〛. Au début du XIXe siècle, l’entretien du chemin de halage était aussi plus strict. Les berges étaient surveillées par des gardes de la navigation, qui infligeaient des amendes aux riverains en cas de négligence. La plantation de jetins de saules était tolérée sur les berges, mais nécessitait une autorisation préalable et une coupe tous les deux ans.
Le canal latéral à la Garonne fut inauguré en 1855 pour faciliter la navigation en toute saison entre Toulouse et Bordeaux. Ce canal, construit sur les terrasses alluviales, prolongeait le canal du Midi et permettait donc de relier Bordeaux à la Méditerranée. Après l’ouverture du canal, le trafic sur la Garonne diminua des deux tiers 〚29〛. En fait, dès 1856, la ligne de chemin de fer Toulouse–Bordeaux allait constituer une véritable concurrence pour le canal latéral, qui aura finalement peu servi. La Garonne a alors été de moins en moins surveillée et entretenue, ce qui favorisa une expansion de la ripisylve, perceptible sur la carte de 1868. Dans le même temps, l’essor des plantations de peupliers se confirmait dans la plaine (Fig. 1). Dans l’arrondissement de Castelsarrasin, il n’y avait « guère d’autre bois de construction que le peuplier 〚...〛, l’honneur et la richesse de nos végétations. 〚...〛 Le peuplier a toujours été avantageusement cultivé sur ces rives, et il y a bien longtemps que l’on a dit de nos paysans qu’ils plantaient ainsi la dot de leur fille lorsque celle-ci naissait » 〚30〛.
Nous n’avons pas retrouvé les minutes de la carte d’état-major révisée en 1908. Dans cette carte, il n’est donc pas possible de distinguer les boisements spontanés des plantations de peuplier (Figs. 2 et 3). En revanche, les surfaces boisées totales étaient pratiquement égales à celles de 1868, soit une largeur moyenne de 761 m. Au cours de la première moitié du XXe siècle, suite à de sévères attaques parasitaires dans les plantations de peuplier, une multitude de clones ont été introduits et plantés, dans une certaine confusion 〚31, 32〛. Beaucoup de planteurs produisaient leurs plants à partir de sujets d’origine inconnue, et d’autres achetaient leurs plants à des pépiniéristes, qui ne disposaient pas de renseignements quant à la variété vendue 〚33〛. Deux clones hybrides ont fini par dominer les plantations garonnaises : le Robusta (mâle) et le Regenerata (femelle). Les attaques parasitaires ont provoqué une baisse massive des plantations. En 1945, les surfaces totales boisées n’atteignaient que 70% de celles de 1908, bien que les boisements spontanés occupassent toujours une surface pratiquement équivalente à celle de 1868 (Figs. 2 et 3). La ripisylve garonnaise a subi une deuxième phase de destruction après 1945. En 35 ans, sa largeur moyenne est passée de 147 à 53 m (soit un recul de 64%), alors que la surface totale boisée restait stable et que la proportion des plantations passait de 72 à 91%. À partir de 1968, deux peupliers hybrides, d’origine italienne, se sont imposés rapidement : le clone femelle I214 et le clone mâle I45/51. Ils sont toujours dominants aujourd’hui.

Évolution des boisements riverains de la Garonne entre 1909 et 1980, sur une section de 42 km à l’amont de la confluence du Tarn.
3.2 Introduction de clones cultivés
L’introduction de cultivars et clones plantés s’est fait par vagues successives avec, à chaque époque, un véritable engouement pour quelques clones seulement (Fig. 4).
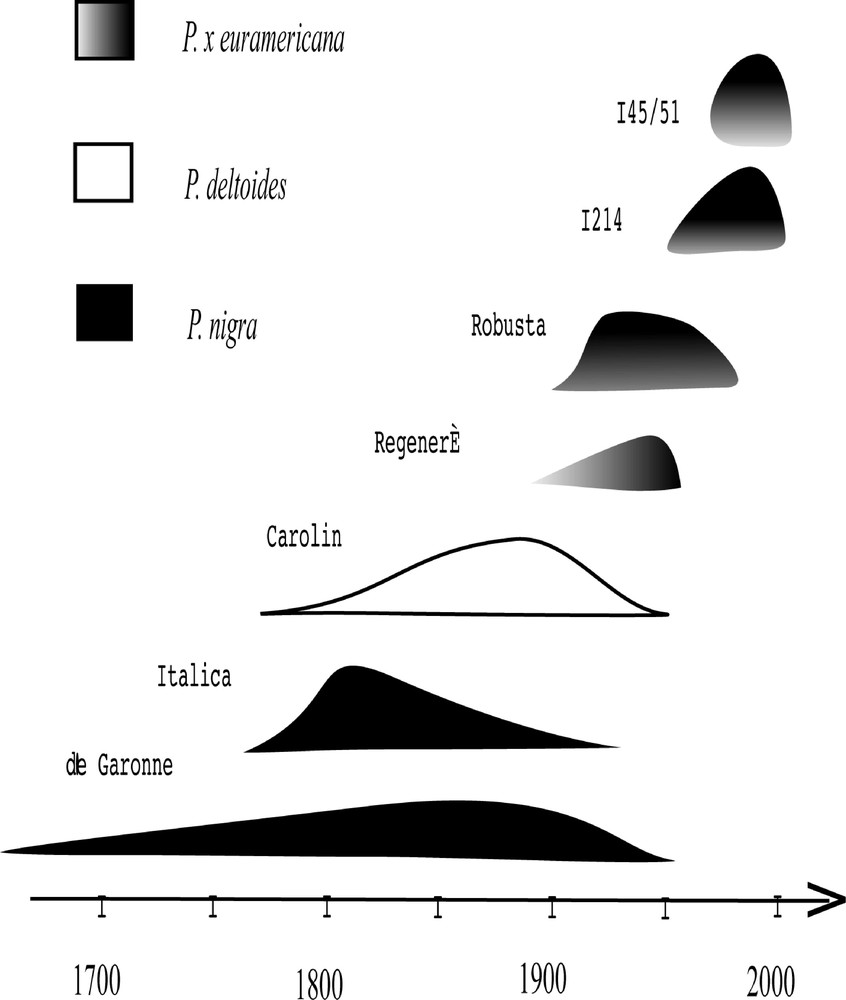
Évolution des principaux clones de peupliers cultivés en moyenne vallée de Garonne.
– Les peupliers « de Garonne » (Populus nigra L. cv. « de Garonne ») ne constituaient pas un cultivar unique, mais un ensemble complexe de clones génétiquement proches les uns des autres. Ils résulteraient de croisements spontanés entre les peupliers noirs issus de populations locales et, sans doute aussi, de croisements avec le peuplier d’Italie 〚23〛. Trois têtes de clones ont été officiellement reconnues et diffusées 〚34〛 : le cultivar « Blanc de Garonne Seilh » (mâle), le cultivar « Vert de Garonne » (femelle) et le cultivar « Sarrazin de Seilh » (femelle).
– Le peuplier d’Italie (Populus nigra L. cv. « italica »), mâle, introduit en France dès 1745, a été qualifié de « prodige » et de « trésor de l’Agriculture » 〚35〛. La diffusion en vallée de Garonne s’est faite à partir la pépinière royale de Montauban vers 1770 〚36〛. Au moment de son introduction en France, il était déjà cultivé depuis longtemps en Italie. Il serait originaire d’Iran ou d’Afghanistan 〚23, 37〛. Par sa présence massive, il a profondément transformé l’aspect et l’économie de la région garonnaise 〚24〛.
– Le peuplier « Carolin » (Populus deltoïdes Marsch ssp. angulata cv. « carolin ») est un clone mâle importé au XVIIIe siècle de Caroline du Sud, l’aire la plus méridionale du peuplier noir américain (black cottonwood) P. deltoides. En vallée de Garonne, il supplanta le peuplier d’Italie dans la deuxième moitié du XIXe siècle 〚19, 23〛.
– Le clone Robusta, Populus x euramericana (Dode) Guinier cv. « robusta », diffusé dès 1895, serait issu du croisement d’un peuplier mâle P. nigra cv. « plantierensis », voisin du peuplier d’Italie, et d’un peuplier femelle américain, P. deltoïdes Marsch ssp. angulata. L’arrêt des plantations massives de Carolin, très sensible aux parasites Dothichiza et Cystospora, a coïncidé presque exactement avec l’essor de celles de Robusta, vers 1920 〚34〛. En 1968, 72% des peupliers plantés dans le Tarn-et-Garonne étaient encore du Robusta 〚38〛. Actuellement, les pépinières de Midi-Pyrénées n’en produisent plus qu’un millier de plan par an.
– Le clone Regenerata (Populus x euramericana (Dode) Guinier cv. « regenerata »), remarqué dès 1814 dans la banlieue parisienne, n’a été diffusé massivement que vers 1940–1950 en vallée de Garonne 〚34〛. Il s’agirait d’un hybride secondaire, résultant du croisement de deux hybrides euraméricains (cv. « marilandica » et cv. « serotina »). La dénomination de peupliers « régénérés » a été très utilisée et englobe sans doute plusieurs hybrides très proches. Ces peupliers femelles étaient réputés avoir une faible fécondité, avec des chatons précocement caducs. Ils ne sont plus cultivés en vallée de Garonne.
– Le clone I214 (Populus x euramericana (Dode) Guinier cv. « I214 ») serait un hybride naturel, repéré et expérimenté dans la vallée du Pô dès 1929, en raison de sa résistance à la tavelure. Il résulterait d’un croisement entre un P. deltoides ssp. angulata ou ssp. monilifera et un peuplier noir européen. Ce clone a régné sur la plus grande partie de la populiculture mondiale de 1950 à 1970 〚39〛. En vallée de Garonne, il a détrôné le clone Robusta vers 1980 〚38〛. Cependant, sa forte sensibilité à Marssonina est la cause de sa régression actuelle.
– Le clone I45/51 (Populus x euramericana (Dode) Guinier cv. « I45/51 ») serait lui aussi un hybride naturel entre un peuplier femelle, P. deltoïdes ssp. angulata, et un hybride mâle, voisin du Robusta. Il a été repéré et diffusé à partir de l’Italie au début des années cinquante. Sa moindre sensibilité à Marssonina et son port rectiligne, qui rappelle le Robusta, en a fait un arbre très apprécié en vallée de Garonne, où il a supplanté le I214 à partir de la fin des années 1980 〚40〛. Il est cependant sensible aux rouilles à Melampsora et son bois est assez fragile, ce qui explique le fait que le clone I214 ait repris sur lui la première place au cours de ces dernières années.
3.3 Perturbation de la dynamique fluviale
Le lit de la Garonne, entre Toulouse et la confluence du Tarn, est resté pratiquement intact jusqu’au début du XIXe siècle. Seuls les abords immédiats du fleuve étaient périodiquement entretenus, pour maintenir un chemin de halage. Les travaux dans le lit sont toujours restés très limités. Ce n’est qu’après 1945 que de grands méandres ont été raccourcis puis, vers 1970, qu’un important programme de fixation du lit a été lancé. Des cordons d’enrochement et des épis ont été construits en alternance d’une rive concave à l’autre et dans les extrados des méandres 〚21〛. Près de 50% des berges du secteur étudié ont ainsi été aménagées. L’extraction massive de granulats dans le lit mineur a démarré à la même époque. Elle a duré une vingtaine d’année, jusqu’à leur interdiction vers 1994. Entre Toulouse et la confluence du Tarn, 33 sites d’extractions ont été inventoriés 〚41〛.
Ailleurs dans le bassin de la Garonne, d’autres aménagements ont perturbé le régime du fleuve. Dès 1862, une partie de l’eau de la Neste, affluent pyrénéen de la Garonne, avait été détournée par un canal vers les rivières gasconnes qui, déconnectées des Pyrénées par le plateau de Lannemezan, subissaient chaque été un assèchement chronique. En 1955, le débit potentiel du canal de la Neste était porté de 7 à 14 m3 s–1. Cette eau ne passe donc plus à Toulouse, en particulier durant l’étiage estival. Le fleuve est ainsi privé de l’équivalent du tiers de son débit objectif d’étiage. La gestion du soutien d’étiage est devenue l’une des préoccupations majeures du Schéma directeur d’aménagement et de gestion des eaux du bassin 〚42〛. Des accords sont négociés avec EDF pour le déstockage partiel des réserves d’eau des grands barrages hydroélectriques, qui totalisent environ 360 × 106 m3 dans les massifs pyrénéens, en amont de la confluence du Tarn. Au cours des vingt dernières années, des barrages supplémentaires et spécifiques pour le soutien d’étiage ont été construits dans les Pyrénées et dans le piémont 〚43〛. Ils totalisent près de 200 × 106 m3.
Les premières conséquences de ces aménagements ont été signalées dès la fin des années quatre-vingts 〚21〛 : stabilisation du lit d’étiage et du lit mineur, diminution de la fréquence et de la durée des submersions dans la plaine alluviale, disparition de certains biotopes, assèchement de bras morts et abaissement du niveau d’étiage. Le lit du fleuve, figé dans ses enrochements, s’est enfoncé verticalement. L’essentiel de cette incision (1,50 m) s’est produit entre 1960 et 1985 〚41, 44〛. En amont de Toulouse, onze barrages hydroélectriques dans le lit du fleuve empêchent la recharge sédimentaire naturelle du secteur aval 〚41, 45〛. Désormais, en de nombreux endroits, les sédiments quaternaires dans lesquels se déplaçaient les méandres ont disparu et la molasse tertiaire est visible en période d’étiage.
L’analyse des cartes historiques (Figs. 1 et 3) nous révèle que la sinuosité du lit principal, mesurée sur 42 km, a fluctué, en deux siècles, entre 1,30 et 1,38 (à l’exception de 1,27 dans la carte de 1908, où le lit du fleuve est particulièrement simplifié et donc peu fiable). Les sinuosités les plus faibles s’observent en 1849 et en 1980, après deux phases d’aménagement du lit. Les sinuosités les plus fortes s’observent en 1868 et en 1945, ce qui témoigne de la réversibilité des phénomènes d’érosion–sédimentation dans la plaine alluviale jusqu’au milieu du XXe siècle. Cette réversibilité n’est plus possible, dans la mesure où le lit mineur a été figé vers 1970. Sur cette section de 42 km, on constate aussi que la longueur cumulée des chenaux secondaires est restée pratiquement égale à celle du chenal principal jusqu’en 1945, soit 54 à 58 km. Les aménagements récents, après 1970, et la mise en culture d’une partie de la ripisylve ont provoqué l’assèchement d’environ 27 km de chenaux secondaires (l’équivalent de la moitié de la longueur du lit principal). Dans le même temps, la surface des sites potentiels de recrutement de P. nigra a diminué des deux tiers entre 1945 et 1980, alors qu’elle était restée pratiquement la même entre 1810 et 1945.
4 Discussion
4.1 Assèchement et simplification du milieu
Depuis trente ans, le fleuve s’écoule dans un lit mineur figé, fortement incisé. En moyenne, la ripisylve n’existe plus que sur une cinquantaine de mètre de large. Elle est moins inondée que dans le passé. Le schéma classique des successions végétales a profondément changé : le milieu est plus sec et la mosaïque forestière, où se juxtaposaient des communautés d’arbres d’âges variés, tend à se simplifier. La ripisylve adulte, encore visible aujourd’hui, est la relique d’une situation antérieure à la chenalisation. Elle n’a réussi à survivre que dans la partie du domaine public fluvial qui n’a jamais été amodiée. Ailleurs, elle a été détruite et remplacée par des cultures ou par des peupleraies. Là où elle existe encore, elle a souvent été perturbée par l’extraction de granulats. L’état sanitaire des boisements spontanés s’est également dégradé. Des phénomènes de dépérissement ont été signalés 〚17〛. Même dans les plantations, les fréquentes ruptures d’alimentation hydrique au niveau des racines fragilisent les clones I214 et I45/51, qui dominent depuis les années 70 et qui ne sont plus vraiment adaptés à la situation garonnaise. Ils subissent de virulentes attaques parasitaires. Aujourd’hui, la plasticité d’un clone pour l’eau est une caractéristique très recherchée et des essais sont menés pour proposer de nouveaux hybrides mieux adaptés. Le remplacement des peupliers par d’autres espèces a également été suggéré 〚46〛.
Sur les berges de la Garonne, la régénération du peuplier et surtout du saule est devenue difficile au cours des dernières décennies 〚20, 47〛. Trois raisons peuvent être invoquées : diminution des surfaces potentielles de recrutement, non-renouvellement périodique de ces surfaces et assèchement du milieu. Les surfaces potentielles de recrutement se limitent aujourd’hui à la partie concave des méandres stabilisés ou à quelques petites îles temporaires. Dans la mesure ou les méandres ne peuvent plus migrer, même partiellement, le rajeunissement de ces surfaces n’est possible que sur place, par suite d’un décapage provoqué par une forte crue. Avant 1970, lorsque les méandres pouvaient encore migrer, de nouvelles surfaces étaient exondées régulièrement et devenaient potentiellement disponibles pour le recrutement. Actuellement, en début d’été, on trouve encore sur les berges des densités de jeunes plantules de peuplier pouvant aller jusqu’à 400 individus m–2. Toutefois, au début de l’été, si le taux de déclin du débit fleuve et du niveau de la nappe qui lui est assujettie est plus fort que le taux de croissance des racines, les chances de survie des plantules sont très faibles 〚48–51〛. Une plantule qui réussit à survivre durant le premier été peut aussi être arrachée par une forte crue d’hiver ou de printemps. Finalement, malgré l’abondance des graines dispersées par un individu, près de 25 millions 〚52〛, il est fréquent qu’aucun recrutement ne soit possible certaines années sur certains sites.
Il est certain qu’au fil des siècles, le maintien des écosystèmes riverains n’a jamais été considéré comme une priorité par les gestionnaires ou les aménageurs. Même dans les années récentes, la gestion quantitative de la ressource en eau, surtout en période d’étiage, est restée marquée par l’émergence d’une priorité, l’irrigation de terres agricoles sur des surfaces qui ont triplé en 30 ans. La loi sur l’eau de 1992 a imposé une approche plus collective et équilibrée de cette ressource et une meilleure gestion de la diversité des milieux riverains. Protéger les écosystèmes aquatiques et les zones humides, valoriser le potentiel piscicole du bassin sont devenus de nouveaux objectifs 〚42〛. Toutefois, cela ne signifie pas un retour à une dynamique fluviale antérieure aux aménagements hydroélectriques ou à la chenalisation.
4.2 Pollution génétique peu probable
Depuis plus de deux siècles et demi, la population garonnaise de P. nigra est entourée de plantations de peupliers dont le matériel génétique est particulièrement restreint. De plus, mis à part le croisement réputé impossible entre un P. deltoides mâle et une P. nigra femelle 〚53〛, il semble que toutes les autres formes d’hybridation entre P. nigra, P. deltoides et leurs hybrides soient possibles (Tableau 2). Toutefois, plusieurs arguments permettent de rejeter l’hypothèse d’une pollution génétique massive.
Possibilités de croisements entre peupliers, d’après 〚1, 8〛. Plusieurs clones interaméricains (hybrides entre P. deltoides et P. trichocarpa) sont actuellement testés en vallée de Garonne.
| ♂ | ♂ | ♂ | ♂ | ||
| P. nigra | P. deltoides | P. x euramericana | interaméricains | ||
| ♀ | P. nigra | oui | non | oui | ? |
| ♀ | P. deltoides | oui | oui | oui | ? |
| ♀ | P. x euramericana | oui | oui | oui | oui |
| ♀ | interaméricains | oui | ? | oui | oui |
Le peuplier noir d’Italie (mâle) a pu provoquer jusqu’au début du XXe siècle une lente dérive génétique des populations locales de P. nigra par croisements spontanés avec des femelles P. nigra réparties dans ces populations. Cette dérive a pu être renforcée par des croisements avec les cultivars femelles locaux (par exemple P. nigra cv « sarrazin de Seilh ») qui, eux-mêmes, avaient déjà comme parent le peuplier d’Italie. Il est donc possible que celui-ci ait imposé au fil des générations dans la population spontanée garonnaise des gènes qui lui sont spécifiques. Les cultivars « de Garonne » mâles, dont le père serait le peuplier d’Italie, ont pu eux aussi fertiliser des femelles des populations locales P. nigra ou des cultivars plantés, et donc contribuer à accélérer cette dérive. Il serait donc important de vérifier, à l’aide de marqueurs génétiques, dans quelle mesure des gènes spécifiques du peuplier d’Italie ont pu être fixés dans la population actuelle et s’il est responsable d’une baisse de la diversité génétique. Toutefois, on peut remarquer que le peuplier d’Italie n’est plus cultivé à grande échelle depuis près d’un siècle et qu’il ne joue plus aujourd’hui qu’un rôle d’ornementation ou de brise-vent. Son port « pyramidal » caractéristique ne semble pas s’être imposé dans les populations spontanées locales. S’il y a eu un jour dérive génétique, celle-ci a pu s’estomper lentement depuis l’arrêt des plantations des peupliers d’Italie.
Le Carolin (P. deltoides mâle), contrairement au peuplier d’Italie et malgré sa présence massive durant près d’un siècle et demi, n’a sans doute eu aucune influence sur les populations spontanées de peupliers noirs garonnais, dans la mesure où les croisements ne sont pas possibles avec P. nigra (femelle). En revanche, des hybridations spontanées entre des peupliers noirs mâles et les deux clones euraméricains femelles dominants (Regenerata et I214), bien que réputés peu fertiles, ont sans doute pu se produire 〚34〛. Les hybridations spontanées faisant intervenir d’autres clones femelles P. deltoides sont restées sans doute très limitées, dans la mesure ou ces clones n’ont jamais été massivement présents en vallée de Garonne.
Des études récentes ont montré que les peupliers femelles seraient fertilisés par un nombre limité de mâles, situés dans leur voisinage immédiat 〚54〛. Or, dans la mesure ou les individus P. nigra femelles sont répartis de façon diffuse au sein de la ripisylve, la probabilité pour que ces individus soient effectivement fertilisés par des hybrides mâles (comme Robusta ou I45/51) est très faible, si l’éloignement est effectivement un facteur limitant. Seuls les individus P. nigra femelles situés en bordures proches des plantations auraient une chance d’être fertilisés par des clones hybrides mâles (Robusta, I45/51).
L’éloignement entre le point de diffusion des graines et les sites potentiels de recrutement constitue un autre facteur susceptible de réduire les risques d’introgression. En effet, si la dispersion des graines peut s’étendre sur des kilomètres, il semble que la grande majorité se dépose à quelques centaines de mètres de l’arbre mère, tout au plus 〚14, 55〛. Or, les sites potentiels de recrutement sont pratiquement limités aux abords immédiats du fleuve et de ses annexes. Ils sont plus rares à proximité des plantations.
Si l’introgression n’est possible que par l’intermédiaire des plantules hybrides qui réussissent à s’installer, on peut supposer que la capacité d’établissement des plantules hybrides est plus faible que celle des plantules P. nigra, étant donné le matériel génétique plus restreint dont elles sont issues. En effet, les quatre hybrides euraméricains les plus répandus dans les plantations au cours du XXe siècle (Robusta, Regenerata, I214 et I45/51) sont fortement marqués génétiquement par la sous-espèce P. deltoides ssp. angulata et par P. nigra cv. « italica ».
Les fragments d’arbres dispersés par l’eau, le vent et les dépôts alluvionnaires peuvent aussi être considérés comme des points de départ potentiels pour de nouveaux individus (en particulier les fragments issus de clones cultivés et périodiquement élagués). Cependant, en milieu naturel, la proportion d’individus issus d’une multiplication végétative semble assez faible 〚14, 56〛. Ce mode de propagation apparaîtrait plutôt comme un moyen, complémentaire à la reproduction sexuée, permettant de renforcer ou de maintenir localement un génotype particulièrement adapté à certains sites ou de résister aux perturbations hydrologiques.
5 Conclusion
Les boisements riverains de la Garonne ont perdu très tôt leur caractère « naturel ». L’homme est souvent intervenu pour essayer de stabiliser les berges par la végétation et, plus rarement, par des enrochements. Il a favorisé certaines espèces : les ormes au Moyen Âge, puis les peupliers et les saules. Cette étude a montré, sur une section de 42 km, que l’essentiel de la ripisylve garonnaise avait été détruit en deux phases distinctes : une première phase vers 1830 (disparition de 598 ha), et une seconde phase dans les années 1970 (disparition de 395 ha). Aujourd’hui, il ne reste plus que 221 ha, soit 19% de la surface de 1810. En d’autres termes, la largeur moyenne de ces boisements spontanés, qui était de 277 m en 1810, n’est plus que de 53 m aujourd’hui. Ces informations ont été extraites de cartes historiques, dont les échelles et les objectifs cartographiques étaient très différents. Il convient donc de comparer les résultats issus de ces cartes avec une certaine prudence. Néanmoins, cette séquence cartographique donne, pour la première fois, une indication quantitative sur l’évolution des surfaces boisées dans la plaine alluviale de la Garonne au cours des deux derniers siècles et permet de distinguer la ripisylve des plantations de peupliers. Les deux phases de destruction de la ripisylve ont eu des conséquences très différentes. Lors de la première phase, au début du XIXe siècle, les plantations de peupliers se sont substituées à une part importante de la ripisylve, mais les aménagements dans le lit du fleuve sont restés très légers. Il n’y a pas eu d’assèchement systématique des bras secondaires et des annexes fluviales. De fait, on constate que la dynamique du fleuve est restée pratiquement intacte entre le milieu du XIXe et celui du XXe siècles. Les mécanismes d’érosion et sédimentation ont conservé leur réversibilité, les surfaces d’atterrissements sont restées équivalentes et la largeur moyenne de la ripisylve a fluctué entre 135 et 168 m. En revanche, lors de la seconde phase de destruction de la ripisylve, au cours des dernières décennies du XXe siècle, des aménagements hydrauliques lourds et une exploitation massive des granulats ont brisé la dynamique naturelle du fleuve. Celui-ci coule désormais dans un chenal figé, qui s’est incisé verticalement. L’alimentation hydrique des arbres est fortement perturbée en période d’étiage 〚57〛. La destruction de la ripisylve a concerné aussi bien les terrains privés que le domaine public fluvial, par suite de fréquentes amodiations. Les boisements spontanés adultes sont aujourd’hui morcelés et apparaissent comme des reliques d’une situation antérieure à la chenalisation. Dans ce nouveau milieu contraignant, P. nigra, espèce pionnière, est toujours présent et se régénère plus facilement que les saules, mais sur des surfaces qui, en 1980, ne représentaient plus que 38% de ce qu’elles étaient en 1945. Les sites de recrutement se limitent désormais aux berges convexes des méandres et à quelques îles, sur lesquelles les successions végétales sont interrompues plus souvent que par le passé.
On peut donc se demander si la population garonnaise de peupliers noirs va pouvoir conserver son potentiel génétique intact ou si, au contraire, elle va subir une dérive génétique. Cette dernière pourrait, en effet, provenir de la présence massive de cultivars et de clones plantés ou, plus récemment d’une modification radicale du milieu et d’une nécessaire adaptation à des conditions hydrologiques plus sèches ainsi qu’à une eau riche en azote. Dans cette étude, nous avons montré que plusieurs arguments pouvaient être invoqués pour rejeter l’hypothèse d’une introgression massive ou d’une dérive génétique forte. Ceci ne signifie pas que cette dérive n’est pas en train d’opérer 〚58〛. Seules des analyses génétiques permettront de répondre avec précision à cette question et d’indiquer l’ampleur des modifications, si elles existent. Il sera intéressant d’évaluer aussi dans quelle proportion les individus P. nigra qui réussissent à s’établir au sein de la ripisylve garonnaise sont issus de graines ou, au contraire, se sont établis par multiplication végétative à la suite de la chute d’un arbre ou d’une branche ou à partir de fragments de racines. En effet, la multiplication végétative est réputée plus facile et plus fréquente chez P. nigra que chez P. deltoides ou P. trichocarpa ; les modifications récentes du régime hydrologique de la Garonne pourraient favoriser ce mode de multiplication, au détriment de la reproduction par graine.
Remerciements
Cette étude a été réalisée grâce à un financement du ministère de l’Aménagement du Territoire et de l’Environnement (DGAD/SRAE/94174 – convention CNRS n° 9502). Elle a pu se poursuivre dans le cadre des deux projets européens : FLOoodplain Biodiversity And Restoration, ENV4-CT96-0317 (FLOBAR) et EVK1-CT-1999-00031 (FLOBAR2).
Abridged version
The black poplar (Populus nigra L.), a pioneer tree species typically found growing along the banks of rivers in Europe, is near extinction in many locations as a direct consequence of a loss in natural habitat. A second threat weighs heavily on P. nigra: the widespread planting of poplar clones in numerous floodplains has the potential to weaken native genetic stocks. In France, research on black poplar has essentially focussed on populations situated in the Loire and Rhone Rivers basins, with no study to date taking into account populations present throughout the Garonne River valley. In the case of the Garonne, while it has been observed that the surface area occupied by riparian woodland has dramatically diminished through the course of time, the scale of woodland loss and historical timing of this loss has never been described. One main aim of this study was, therefore, to follow the evolution of floodplain woodland coverage along the Garonne River, using a series of historical maps as records of change. For each map, quantitative information was sought on the respective surface areas occupied by planted poplars and the natural riparian woodland. A second key objective in this study was to evaluate, over the same historical period, the impact of two major sources of disturbance to black poplar populations along the Garonne River: the widespread introduction of poplar cultivars and shifts in the intensity of river management activities.
The existence of six historical maps covering a 42-km section of the Garonne River between Grenade (Haute-Garonne) and Moissac (Tarn-et-Garonne) allowed an investigation into changes in riparian woodland coverage over a period of two centuries. The maps were scanned, compiled in a raster database and georeferenced. The natural riparian woodland, poplar plantations, and active alluvial zones in each map were identified and digitised. Other documents, originating from a number of different archival sources, provided more detailed information about the historical transitions in poplar cultivation throughout the Garonne valley. The results revealed that the vast majority of the Garonne riparian woodland, which occupied almost 1200 ha in 1810 over a 42-km stretch, was destroyed during two distinct periods: a first wave around 1830 (disappearance of 598 ha) and a second wave in the 1970s (disappearance of 395 ha). Today, only 221 ha remain; that is, only 19% of the woodland that existed in 1810. Viewed from another perspective, the width of the natural riparian woodland bordering the Garonne River has receded from a bandwidth of 277 m in 1810 to 53 m today. During the first wave of destruction, in the early 19th century, the natural riparian woodland was replaced largely by poplar plantations, with only limited changes detected to the riverbed from river management activities. There is little evidence of a systematic drying-out of side arms and other fluvial connections during that period and the natural river dynamics appear, in fact, to have remained almost intact from the beginning of the 19th century until the middle of the 20th century. During this period, patterns of erosion and sedimentation remained reversible, with the surface areas of denuded areas remaining constant and the width of the riparian woodland fluctuating between 135 and 168 m. During the second wave of large-scale riparian woodland loss, around 1970, an intensification in river management activities and heavy exploitation of gravel resulted in a radical breakdown of the natural river dynamics, with a vertical incision of about 1.5 m. The river has since flowed in a constrained and increasingly incised channel, such that during periods of low flow the hydrological links to trees is under intense strain. Under these new and constrained environmental conditions, P. nigra is nevertheless still present and capable of regeneration on surface areas that represent only 38% of what they were in 1945.
It is debateable whether the genetic potential of the Garonne black poplar populations has remained intact or, on the contrary has succumbed to a loss in genetic variety. This latter possibility could in fact have developed through mixing with the dense stands of poplar cultivars in the Garonne valley and, more recently, from the radical changes of the hydrological conditions. This new situation may have favoured genotypes that are better adapted to drier conditions and nitrogen-enriched water. The introduction of distinct cultivars and wide scale planting of these clones has occurred in successive waves with the heavy dominance of only a small number of clonal types such as Italica, Carolin, and hybrids such as Robusta, Regenerata, I 45/51 and I 214. In this study, we present a number of different arguments rejecting the hypotheses that there is widespread introgression or a significant decline in genetic diversity among the natural populations. This does not signify, however, that a decline is not presently underway. It will only be possible to identify and define the extent of any decline in genetic diversity in a reliable manner through the genetic screening of populations.