

1 Introduction
Monoculture has become predominant in industrialized agriculture over the last hundred years due to mechanization of work and the increased yield it procures. In this system, a small number of cultivars are grown and the resulting low genetic diversity brings with it severe disadvantages, such as vulnerability to diseases, which necessitate the generalized use of pesticides. Furthermore, breeding programs for disease control are based mainly on specific resistance genes. The efficacy of this type of resistance has often been short-lived, three to five years per resistance gene. The use of single resistance genes in large-scale monocultures is a source of strong selective pressure for pathogen races capable of overcoming the resistance, with the corollary that cultivar resistances have to be renewed constantly. A recent example is the breakdown of the Yr17 resistance to yellow (stripe) rust of wheat in northern Europe [1]. In England and Denmark, cultivars with Yr17 resistance were grown on an increasingly large area over the period from 1994 to 1998, and this was followed by a rise in the frequency of corresponding virulence to 100%. Elsewhere in northern Europe, Yr17 cultivars were grown on a smaller scale during this period. However, virulence for Yr17 was detected in France and Germany in 1997 and had reached a frequency in excess of 70% by 1999. The homogeneity of resistance genes in the crops is responsible for their breakdown. This problem could be mitigated by introducing functional diversity within the crop.
It is apparent that it is not the resistance genes themselves that are inefficient but the way they are managed. To avoid or limit some of the problems of monocultures, diversity needs to be introduced and carefully managed. There are two practical ways of organizing the cultivation of resistant cultivars – in different contiguous fields or within the same field. Cultivar mixtures, made up of cultivars having similar agronomic and technological characteristics but differing for their resistance genes grown in the same field, are the most efficient way to reduce epidemic spread [2–5]. The use of cultivar mixtures is based on the principle of risk spreading. It is usually very difficult to predict which cultivar will be the highest yielding in the next season in any specific location. By choosing several cultivars with high potential, the risk of a large loss due in particular to an epidemic is minimized.
2 Review of cultivar mixtures
2.1 Mechanisms of cultivar mixtures for reducing epidemics
In diverse populations, many genetic and ecological interactions between plants play a role in keeping disease levels low [3,4,6,7]. The principal mechanism by which levels of disease are reduced in cultivar mixtures is the loss of inoculum due to the presence of resistant plants between susceptible ones. In addition, resistance induced by avirulent races can reduce infection by virulent races [5]. A mixture is composed of cultivars that are susceptible to some of the races present in the pathogen population and resistant to others. The distance between host plants possessing the same resistance is greater than in the pure stands. The probability of a spore released from a lesion being deposited on susceptible tissue is reduced in proportion to the density of susceptible plants, i.e., in proportion to the increased distance between susceptible plants. In addition, the presence of resistant plants in the canopy provides a physical barrier against spore dispersal. These two mechanisms, the density of susceptible plants and the barrier effects of resistant plants, are mechanical effects related to the dispersal of the pathogen spores. The magnitude of disease reduction depends therefore on parameters such as the spore dispersal gradient, lesion growth rate, plant size, and the distribution of plant genotypes (groups of plants or random distribution) [8,9].
Furthermore, in cultivar mixtures, the genetic diversity of the pathogen population is greater than in a pure stand and, for a given host component, virulent and avirulent spores coexist. Therefore, a third mechanism comes into play, this time a physiological one, that of induced resistance. This enhances plant resistance, providing protection from subsequent infection by normally virulent races. This protection is based on the stimulation of defence mechanisms and metabolic changes that speed up the process of pathogen recognition. It has been argued that induced resistance should be durable. Overall, induced resistance reduces epidemiologically important parameters such as infection efficiency, lesion growth, and sporulation rate. These effects become even more important at the polycyclic level in the field. While induced resistance can be local or systemic in plants, in the examples of foliar diseases of small grains, induced resistance seems mainly to have localized effects. However, computer simulations indicate that even locally induced resistance can be effective in reducing disease severity in cultivar mixtures at the epidemic level, with efficiency increasing with the number of infection cycles [10]. Estimates made from field studies indicated that induced resistance was responsible for a 24% reduction in powdery mildew on barley cultivar mixtures [11] and a 20–40% reduction in yellow rust in wheat cultivar mixtures (Fig. 1) [12].
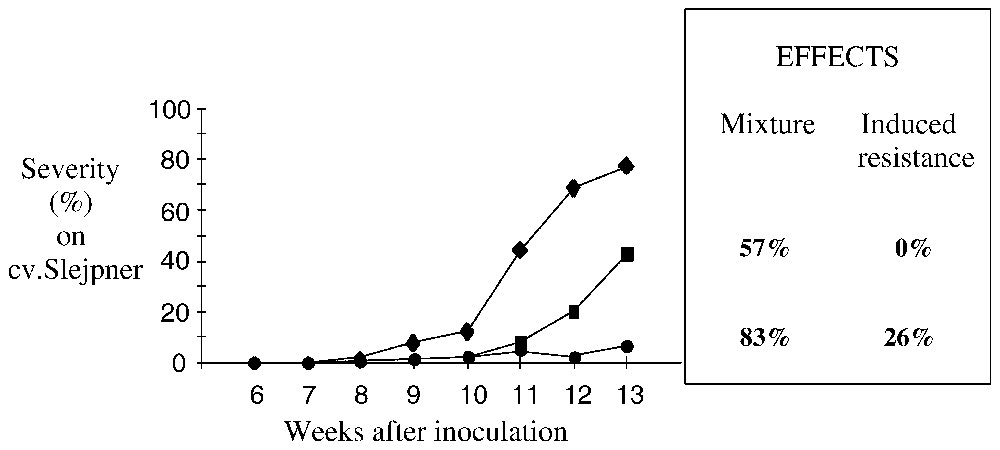
Changes in the severity of wheat yellow rust on field plots, 6–13 weeks after inoculation with the yellow rust isolate J89108 (race 232E137, virulent on Yr9, avirulent on Yr6) and the inducer isolate J89101 (race 45E140, virulent on Yr6, avirulent on Yr9). cv. Slejpner (Yr9) in a pure stand (); cv. Slejpner mixed with the totally resistant cv. Estica (1:2) (■); cv. Slejpner mixed with cv. Arcane (Yr6) (1:2) (●). Each point was the mean of 75 scoring points (15 tillers each) and four replicates. The mixture effect was calculated on the area under the disease progress curves (AUDPC) as the relative AUDPC differences between cv. Slejpner in the mixture and in pure stand. The induced resistance effect was the difference between the cvs. Slejpner:Estica mixture and the cvs. Slejpner:Arcane mixture (de Vallavieille-Pope and Goyeau, unpublished). Masquer
Changes in the severity of wheat yellow rust on field plots, 6–13 weeks after inoculation with the yellow rust isolate J89108 (race 232E137, virulent on Yr9, avirulent on Yr6) and the inducer isolate J89101 (race 45E140, virulent on ... Lire la suite
Cultivar mixtures do not eliminate the pathogen as a fungicide might. Rather, they reduce the rate of disease progress by eliminating large numbers of spores at each cycle of pathogen multiplication. Cultivar mixtures result in a high level of partial resistance due to collective resistance.
In addition to disease resistance diversity, the different cultivars that go into mixtures also react differently to abiotic stresses. Cultivar mixtures compensate for variations in environmental conditions such as drought and freezing and therefore limit the risks of yield loss. Because of the interactions among the components, mixtures provide a buffer against environmental variation [5]. Therefore, not only are cultivar mixtures able to provide control against airborne diseases, enabling fungicide treatment to be delayed or reduced [13], but they also offer greater yield stability.
Very few studies have focused on the harvest quality of mixtures. Compensation can also occur regarding product quality, as demonstrated by the protein content of wheat [14]. Under low nitrogen fertility conditions, a 2:1 cultivar mixture yielded as much as did the high yielding cultivar alone, but with the same high quality as the low yielding cultivar.
2.2 Disease spectrum of efficiency of cultivar mixtures
Cultivar mixtures can limit the spread of pathogens that have an airborne dispersal phase and cause polycyclic epidemics (cereal powdery mildew, rusts, septoria leaf blotch, helminthosporium, and rhynchosporium diseases), even to the extent that the use of fungicides is no longer necessary. In the case of cereal rusts and powdery mildew, most resistance is complete, and mixtures can reduce disease severity by 40 to 80% compared to the mean of the pure stands [4,15]. In the case of septoria leaf blotch, most cultivar resistance to Mycosphaerella graminicola and Phaesphaeria nodorum is partial, and there is a high degree of genetic diversity in the pathogen population [16]. Mixtures are therefore less efficient at reducing disease severity. Cultivar mixtures reduced tritici and nodorum septoria blotch by 6 to 27% in field experiments [17–19]. However, a cultivar mixture composed of one susceptible cultivar and one cultivar with intermediate disease resistance (1:1) did not reduce septoria tritici leaf blotch progression [20]. Wheat cultivar mixtures have been shown to improve the control of a virus disease [21]. However, cultivar mixtures are ineffective against soil-borne diseases such as eyespot due to Pseudocercosporella herpotrichoides [22].
2.3 Yield stability
In general, yield stability is higher in cultivar mixtures than in pure stands. Yield stability of mixtures compared to pure stands has been assessed from experimental small-grain trials. The genotype rankings over several years are higher in the mixtures [23] (Fig. 2), the means are high and the standard errors are lower, and regression coefficients between yields and environmental indexes are low [24]. Yield stability was also assessed by analysing the genotype×environment interaction (Belhaj Fraj, unpublished). On-farm trial environments were characterized by the factors limiting wheat growth and yield of a reference cultivar present in all the mixtures tested. These factors were eyespot, septoria leaf blotch, nitrogen deficiency, water deficit, and topsoil compaction. Regardless of the yield potential of the location, over a two-year period in 15 environments the cultivar mixtures ranked among the most stable cultivars. When yield was limited by water deficit before anthesis, the cultivar mixture was clearly more stable than its components and over-yielded the best cultivar. These results illustrate that heterogeneous populations could result in a better exploitation of environmental resources and could be advantageous under suboptimal conditions [5].
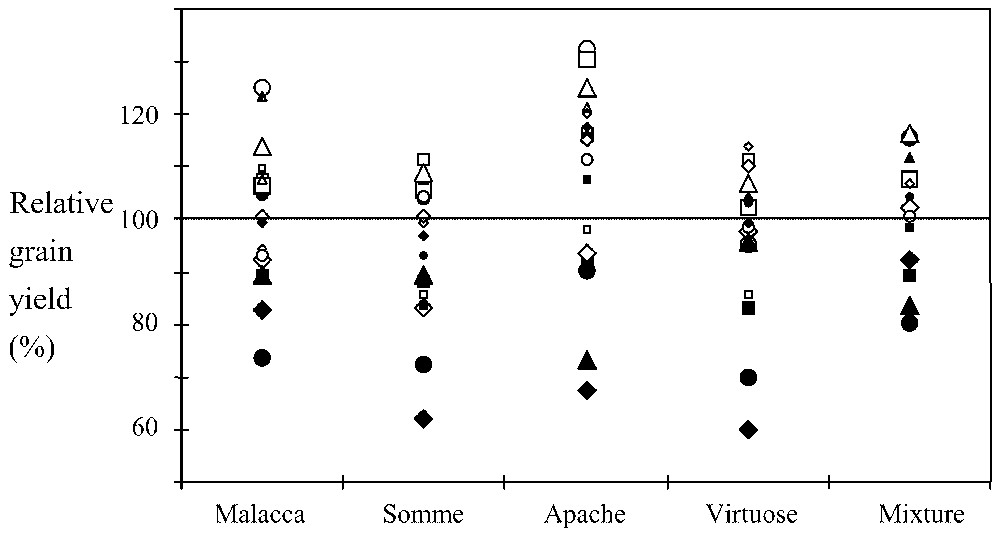
Relative grain yield of four wheat cultivars grown in pure stands and as a mixture in on-farm trials over two consecutive years. The mean yield of the pure stands on 19 locations=100 (Belhaj Fraj et al., unpublished).
2.4 Resistance durability
In the long term, the risk is that the cultivar mixtures will select complex races that are able to develop on most or all of the mixture components. This risk can be reduced by regularly changing the components of the mixtures, as a function of the evolution of the pathogen populations. Several studies have addressed the question of the durability of resistance with the van der Plank concept [25] of stabilizing selection, based on the hypothesis that unnecessary virulence genes will reduce pathogen fitness when infecting a susceptible host [26]. Besides this cost of virulence, other selective forces can influence pathogen evolution in cultivar mixtures and may reduce the rate at which complex races develop. Diversity for aggressiveness among the different isolates of a race could slow down the rate of selection of complex races versus simple races [27]. Simple races always reproduce on the same cultivar whereas complex races are able to grow on several cultivars. During an epidemic in pure stands, the fittest components of the pathogen population on the cultivar are selected; in the mixture the pathogen is exposed to several cultivars and selection is therefore slower. In a field experiment on powdery mildew on wheat grown in pure stands or mixtures, isolates sampled at the end of the epidemic from the mixtures were found to be less aggressive than those from pure stands [28]. Differential adaptation, i.e., adaptation within a race to a specific host genotypic background, may prevent the rapid evolution of complex races in mixtures.
2.5 Selection of cultivars for mixtures
In a diversified crop, agronomic characteristics such as harvest maturity and quality should be as similar as possible, and other traits such as disease resistance should be diversified. At first, diversification was achieved at the level of the gene using multiline varieties, i.e., association of lines differing solely for resistance genes against one or two diseases [2,29]. A current successful application of multilines can be found in the use of the club wheat multiline ‘Rely’ in Washington State in the northwestern USA [30] and of the coffee bush ‘Colombia’ multiline in Colombia. However, multilines are not generally acceptable because breeding is time-consuming and the homogeneity of the lines for many traits other than disease resistances does not provide compensation against abiotic stresses. Therefore, diversification is mainly achieved using cultivar mixtures that can be easily constituted from available commercial cultivars without any further breeding for agronomic uniformity [4].
The aptitude of cultivars to perform in mixtures has been studied with cultivars exhibiting several contrasting characteristics, such as differences in specific and complete resistance genes, grain quality, plant height, and earliness. Such work has usually been carried out after artificial inoculation. General and specific mixing abilities have been analysed for two-cultivar mixtures as compared to pure stands [31], while in practice, cultivar mixtures usually have four to five components rather than two. Knott and Mundt (1990) [32] have suggested that mixing ability estimates derived from two-way mixtures could be used to predict the performance of complex mixtures with more than two components. Recently, a model was developed to predict the mixing abilities of complex mixtures on the basis of results from two-component mixtures and experimental data from wheat plots inoculated with Puccinia striiformis f.sp. tritici, the causal agent of yellow rust. In this specific case, similar predictions were obtained using mixing abilities and the mean of the pure stands, resulting from a low competitive effect compared to the additive effect between the tested cultivars [33].
In practice, cultivars to be mixed are chosen for their homogeneity in grain quality and agronomic characteristics such as stem height and earliness at maturity, as well as complementary resistances to fungal pathogens. Not all mixtures provide equivalent disease protection or yield increases; they can be better than, equal to, or worse than the mean of the components grown in pure stands. In practice, mixtures are constituted of commercial cultivars, each of which is selected primarily for use in monoculture with no consideration of the performance in mixtures. The performance of cultivars in mixtures should be evaluated because this cannot be extrapolated from their performance in pure stands.
In order to reduce the number of cultivar combinations for evaluation, a field experiment was conducted to examine the mixing abilities of 12 bread wheat cultivars currently used for commercial production in France. Two-way mixtures and four-way mixtures were compared over two years. The cultivars chosen were among those most resistant to foliar diseases. Both the two-way mixtures and the four-way mixtures performed better than the mean of the components grown in pure stands for Septoria tritici blotch severity, grain yield components, and grain protein content (Mille, unpublished). Four-way mixtures were particularly efficient (significant reduction of septoria leaf blotch severity and increased grain protein content) compared to the mean of the pure stands, as were the two-way mixtures also, but the four-way mixtures were on no occasion better than the mean of the corresponding six two-way mixtures. The four-way mixture efficiency was higher but close to the estimation based on the two-way mixtures. A more precise prediction of four-way mixture performance was obtained from two-way mixtures than from the pure stands.
One four-way mixture was classified as superior to the other mixtures for all criteria: disease severity, yield, and protein content. In this case, all but one of the corresponding two-way mixtures were better than the mean of the two pure stands for the criteria tested. The overall mean of the six two-way mixtures was not significantly different from the corresponding four-way mixture. For the four-way mixture with the poorest results across the same criteria, the mixture efficiency of the corresponding six two-way mixtures was low or null compared with the two pure stands. However, the performance of the four-way mixture was slightly higher than the mean of the six two-way mixtures and the mean of the four pure stands, indicating a positive effect of competition between the cultivars.
Disease reduction, grain yield, and grain protein content in the four-way mixtures were better correlated to those of two-way mixtures than to those of the pure stands, clearly suggesting that the two-way mixtures predict the four-way mixture performance better than do the pure stands. These results indicate that two-way mixtures can be used for the field evaluation and selection of the most appropriate combinations. It is suggested that binary mixtures should be tested as a first step to eliminate poor combinations. Complex mixtures could be tested in a second step, once the specific behaviour of each component is known.
2.6 Mixtures in practice
A long time has passed since the concept of within-field diversification was promoted by Jensen in 1952 [29], but the practical use of cultivar mixtures is still limited, although used in countries with varied economic systems. In cereal production for animal feed there are few practical objections to growing mixtures, and some farmers are therefore using mixtures in practice, e.g., in Poland and Canada. In the high-quality market such as for malting and baking, there is resistance from end-users. The failure of barley mixtures to achieve commercial acceptance in the UK is largely due to the expectation that malting performance will be uneven [34]. However, mixtures of wheat and barley are used for baking and brewing in Switzerland, Poland, and the USA. In Denmark, mixtures of spring barley and winter barley have been grown since 1979 and 1988, respectively. In the Pacific Northwest of the USA, 20% of wheat acreage is grown in mixtures.
Large-scale observations demonstrate the success of this agricultural practice. The spring barley crop sown in cultivar mixtures in the former East Germany increased from 0 to 350 000 ha during the period 1984–1990, while powdery mildew incidence and fungicide use declined by 80% and malting quality remained high [35]. Recently, rice cultivar mixtures have been planted in China in contiguous fields totalising 800 ha in 1998 and 3300 ha in 1999. Highly susceptible cultivars planted in mixtures with a resistant cultivar had 90% greater yield, and blast severity was 94% lower than when they were grown in pure stands [36]. In this case, harvesting by hand ensured that rice cultivars with different qualities could be separated and kept for their specific markets. The concept has also been applied successfully for the last 20 years on a perennial crop in Colombia, where multilines of coffee are grown over a large area (300 000 ha) to provide protection from coffee leaf rust, caused by Hemileia vastatrix, under climatic conditions conducive to the disease [37].
In France, cultivar mixtures are not yet accepted. An experiment was set up therefore in the north to test whether this cultural practice could be useful for bread wheat. A series of on-farm experiments, totalling 28 environments over 250 ha, tested four different four-cultivar bread wheat mixtures and the associated pure stands over a three-year period. The integrated crop management system aimed at using 30% less input than the system generally practiced in the region. Only one foliar fungicide spray was applied in most of the environments. On average, 6% less Septoria tritici blotch severity was observed in the mixtures compared to the mean of the pure stands; yield was increased by 0.32 t ha−1, and protein content by 0.54% (Belhaj Fraj unpublished). Yield and protein content increased in parallel, and the total amount of the accumulated nitrogen in grains was 12.5 kgN ha−1 higher than the mean of the pure cultivars. Baking quality of the mixtures was equal to that of the mean of the pure stands (Table 1). The performances of two of the mixtures approached that of the best cultivar. This study clearly demonstrates the practical advantages of wheat cultivar mixtures in the region.
Efficiencya of wheat cultivar mixtures compared with the means of the pure stands (%) in on-farm trials (Belhaj Fraj et al., unpublished)
| 2000 | 2001–2002 | |||
| SMRY | SMRO | TMAL | SMAV | |
| Septoria tritici blotch severity | 1.7 | 17.7 | 18.4 | 25.2∗ |
| Grain yield | −1.7 | 7.8 | 2.3 | 5.9∗ |
| Grain protein content | 3.6 | 8.5 | 2.7 | 1.8 |
| Baking quality | −0.3 | 3.4 | −0.5 | 0.1 |
a Decreased disease severity and increased yield and baking quality ratings expressed as the relative differences between the values observed in the mixture and the mean of the pure stands.
∗ Significant mixture efficiencies at P⩽0.05.
Some of the resistance to cultivar mixtures from end-users is related to the requirements of traceability. Newly developed molecular methods for cultivar identification make it possible to detect mixture components in the harvest using DNA tests. Microsatellite markers have been selected to identify bread wheat cultivars commonly grown in France and to estimate the relative proportions of cultivar and hybrid grains in the harvests of four different four-cultivar mixtures planted in equal proportions in on-farm trials [38]. Among the grains harvested, the cultivar proportions ranged from 15 to 37%. No contamination was found from other widely grown cultivars. Hybrids accounted for between 1.3 and 6.3% of the grains produced in the four-cultivar mixtures. This technique enables mixtures to be subjected to similar standards of scrutiny as those currently applied to pure stands.
3 Conclusion
There is an increasing demand for sustainable crop production characterized by reduced inputs of pesticides and chemical fertilizers and increased agricultural biodiversity. In general, low-input crop systems produce more variable yields than in higher-input systems, since the inputs used are insufficient to control all the environmental variations. One way to stabilize crop yields and quality is to make use of plant genetic diversity, growing cultivar mixtures constituted from available commercial cultivars without any further breeding for agronomic uniformity. Although the efficiency of cultivar mixtures is partial, this cultural practice contributes to disease control and can be integrated into other agronomic practices aimed at reducing disease. These practices, used in an integrated crop management system [39], include delaying sowing dates, reducing crop density, and adjusting the quantities of pesticides and fertilizers to crop needs for a target yield.
Future research on cultivar mixtures could usefully include studies on mixing ability by testing binary mixtures to eliminate inappropriate cultivar pairs. Mixtures should be chosen to respond to a specific need of the end-users, e.g., baking, animal feed, and for a specific crop management system (biological, integrated, productive). These experiments should be carried out in the region where the mixtures are intended to be grown, so that the actual climate, soil, and disease pressure conditions are fully taken into account. A longer-term investigation of plant development and inter-plant interactions should be formulated in order to identify the traits that favour compensation between cultivars. Furthermore, functional diversity could be achieved more quickly with the assistance of the new breeding technologies for selecting cultivars with different resistance genes, but similar agronomic traits.
Questions / Answers – Questions / Réponses
Question de Michel Thellier
Lorsqu'on a trouvé un mélange de variétés pour augmenter la résistance, ce résultat est-il robuste, ou bien le parasite s'adapte-t-il rapidement, obligeant à modifier fréquemment le mélange de variétés ?
Réponse de Claude de Vallavieille-Pope
La durabilité des résistances des mélanges variétaux a été montrée dans les cas d'utilisation sur de grandes superficies durant plusieurs années. Plusieurs gènes de résistance d'orge de printemps vis-à-vis de l'oı̈dium qui étaient contournés en Europe dans les monocultures sont demeurés efficaces dans les associations variétales pratiquées en RDA. Durant sept années, les superficies cultivées en mélange ont augmenté jusqu'à 360 000 ha et le gène Mla12, par exemple, efficace et très employé dans les mélanges, était surmonté ailleurs en Europe. De même dans la région ouest des États-Unis, après dix années d'utilisation d'une multilignée de blé, seulement la moitié des résistances aux rouilles jaune et brune des dix composants a été contournée et la protection de la multilignée est demeurée satisfaisante. Par ailleurs, des études théoriques montrant que le coût des virulences inutiles et l'adaptation différentielle des isolats vis-à-vis d'un génotype hôte expliquent le désavantage des races complexes du parasite par rapport aux races simples, se développant sur un seul composant du mélange.
Question de Jean-Louis Bonnemain
La stratégie d'association variétale est intéressante. Peut-on l'appliquer à des cultures autres que les graminées ? Je pense par exemple à la vigne.
Réponse de Claude de Vallavieille-Pope
Il est vrai que cette pratique culturale s'applique particulièrement bien aux céréales : il est possible de renouveler les composants régulièrement et la diversité spatiale entre les individus est élevée. Ce concept s'applique également aux cultures pérennes. Un exemple convaincant est celui des multilignées de caféier pour résister à la rouille orangée. Depuis 20 années, des multilignées de caféier sont cultivées sur 300 000 ha en Colombie, et sont résistantes. La vigne est plus difficile à gérer, étant donné que les possibilités de diversification des greffons est très limitée. Le développement de techniques facilitant l'introduction de différents gènes de résistance dans un même fond génétique devrait permettre de pallier cet inconvénient majeur. L'intérêt d'associations variétales d'autres cultures que les céréales sont étudiées à l'Inra : des associations de pommiers vis-à-vis de la tavelure et de pomme de terre vis-à-vis du mildiou sont généralement moins sévèrement attaquées que la moyenne des cultures pures.
Question de Pierre Devaux
Les industriels sont-ils prêts à utiliser les mélanges variétaux, dont certaines caractéristiques technologiques ne sont peut être pas identiques, à l'heure où la traçabilité est devenue un critère si important ?
Réponse de Claude de Vallavieille-Pope
En effet il reste à franchir la barrière commerciale. Il importe d'associer des variétés qui aient la même destination industrielle. La qualité de l'association sera alors en principe au moins égale à celle de la moyenne des composés purs. Nous avons montré que les outils moléculaires permettent de vérifier la composition de la récolte de l'association variétale et de lever les objections de traçabilité. Dans certains pays, comme les États-Unis, les récoltes de blé sont achetées selon leur qualité, sans impératif de pureté variétale. En France, les variétés sont déclarées à la coopérative, mais, par la suite, plusieurs variétés sont mélangées dans un même silo de stockage. Enfin, c'est tout l'art du meunier ou du brasseur d'associer les variétés en vue de la qualité du produit final. Certains industriels sont prêts à utiliser les mélanges variétaux. Nous avons travaillé avec des meuniers, qui dans le cadre de cultures contractualisées, ont accepté de racheter la récolte de mélanges variétaux de blé.
Question de Gilles Bompeix
Quelle est la place de l'utilisation des mélanges variétaux dans la lutte intégrée ? Quelles sont les causes de la non-extension des techniques décrites par Claude Pope (auxquelles je crois !), économiques (évaluation par les agriculteurs de l'efficacité) ou plus simplement psychologiques (évaluation irrationnelle du risque) ?
Réponse de Claude de Vallavieille-Pope
Nous avons montré que, pour la culture de blé, les mélanges variétaux peuvent être un des composants d'un système de culture de protection intégrée. Les mélanges variétaux contribuent, par la réduction de la sévérité des maladies foliaires, au gain en rendement et en qualité de la récolte et donc, par voie de conséquence, à la réduction d'intrants. Les effets sont modérés et significatifs. Il est ainsi possible d'améliorer la production variétale d'un gain équivalent à celui justifiant d'une inscription variétale, en utilisant des variétés déjà disponibles.
Je ne pense pas qu'il y ait de causes psychologiques au développement de cette pratique culturale. Il fallait faire la preuve que cette pratique est économiquement viable dans le système français, avec des variétés françaises et un système de culture adapté, et la preuve vient d'être apportée.
Question d'André Gallais
Vous avez montré que le mélange de variétés différentes est efficace pour limiter le développement épidémique. Je suppose que l'on ne peut pas mélanger n'importe quoi, sinon cela risquerait d'être aux dépens de la production et de sa qualité. Comment raisonner le choix des variétés à associer ?
Réponse de Claude de Vallavieille-Pope
Les variétés à associer doivent être compatibles d'un point de vue technologique et agronomique. Les critères retenus avec J.-M. Meynard, agronome à Grignon, sont une précocité à la montaison identique entre les variétés de blé, pour que le mélange reste équilibré, et une même hauteur des plantes, pour éviter l'ombrage de certaines variétés par rapport aux autres. Les tests systématiques pour l'aptitude spécifique et générale au mélange impliquent un travail laborieux, comparant toutes les variétés pures et les mélanges binaires. Nous proposons de tester certains mélanges binaires pour éliminer les couples incompatibles, et ensuite de composer des mélanges plus complexes à partir des binaires sélectionnés. Quatre à cinq composants constituent un bon compromis entre efficacité du mélange et contraintes pratiques. Des recherches devront être poursuivies sur ce sujet, pour à la fois introduire une diversité pour les gènes de résistance et trouver des critères d'assemblage.