

1 Introduction
Many male birds sing in choruses that peak at dawn and at dusk during the reproduction season. Functions associated with bird song are various: attracting and stimulating females, repelling rivals, conveying information about the quality of the singer. Reports generally concern songbirds or oscines [1]. The singing activity of gallinaceous males such as quail occurs in traditional reproduction areas or ‘mating centres’ where several males gather (generally less than ten European quail in a reproduction area). Pairs are formed at these mating centres and, once mated, males stay with their females and no longer participate in the chorus. Males then sing only very occasionally, for example when their females are out of sight [2,3]. In the wild, we know more about European quail than about Japanese quail. As Japanese quail are domesticated, they are easier to study in the laboratory than in the wild. As the behavioural repertoires of wild European quail and domestic Japanese quail are very similar, we investigated whether Japanese quail performed choruses similar to those of European quail observed in the wild.
Each male quail has an individual inborn song theme. Documented observations on the reproductive behaviour of Japanese quail in the wild are few. However, the structure of European quail male choruses is well known [4]. Males at a mating centre can share precise temporal characteristics of songs, such as intervals between syllables, and so, could be kin-related [5]. The fact that males in a chorus are kin-related could be the result of affinities between kin or that migration brought them all back to the same area. Songs could be a means for inter-individual recognition as male quail in the wild do not generally see one another. On these sites, males could compete for females or co-operate to attract more females during vocal emission. Competition could be expressed by strong synchronization among males and the quantity of song emitted could increase with the number of males in the group.
The aim of this study was, first, to investigate the fine structure of choruses in the Japanese quail. In particular, we raised the following questions: Do all males invest similarly in a chorus? Are some males more active than others? In the laboratory, we monitored the parts played by individual Japanese male quail in the general vocal activity of groups during reproduction choruses, and we estimated the stability of inter-individual differences. Constancy of this vocal structure was then tested experimentally. In a preliminary study, we analysed the consequences of removing one male on the singing activities of the others. In the field, males leave the chorus group one after the other to pair with females. They then stop singing. The question was what impact this leaving could have on the rest of the group. As field data [4] suggest that singing groups are formed of kin-related birds, we compared a group of brothers to a group of non-kin.
2 Materials and methods
2.1 Animals
The males tested were two years old, and therefore in their second reproduction season. All were fully sexually developed, according to several indices, including development of their cloacal glands [6] and singing activity. Five groups were observed: four groups of kin-related males: K1, K2, K3, K4 (respectively four, five, five, and seven brothers) and one group of six non-related males: NK. Quail families were produced in our laboratory from breeders from an industrial husbandry. The non-related males came from the same industrial husbandry. The six males in this NK group were taken randomly from a stock of several thousand birds and so they were very likely unrelated. Groups were placed in batteries in different sound-attenuated chambers and birds in a group had been together for more than six months. Birds were placed in individual cages with a feeder and a drinker under a 14:10-h light:dark cycle, without twilight, and a constant temperature (). Observations were made during the dark period with a dim green light of less than .
2.2 Measures and experimental protocol
The general singing activity of entire groups was recorded with a computer (Fig. 1). Daily organization was similar to the specific rhythm of singing activity previously observed for the Japanese quail in the laboratory and for the European quail in the field during the breeding season [4,7,8]. The most intense activity was recorded during the two hours preceding lights-on and could be compared to the dawn peak of choruses in the field. Singing activity of each male was recorded during these two hours preceding lights-on.
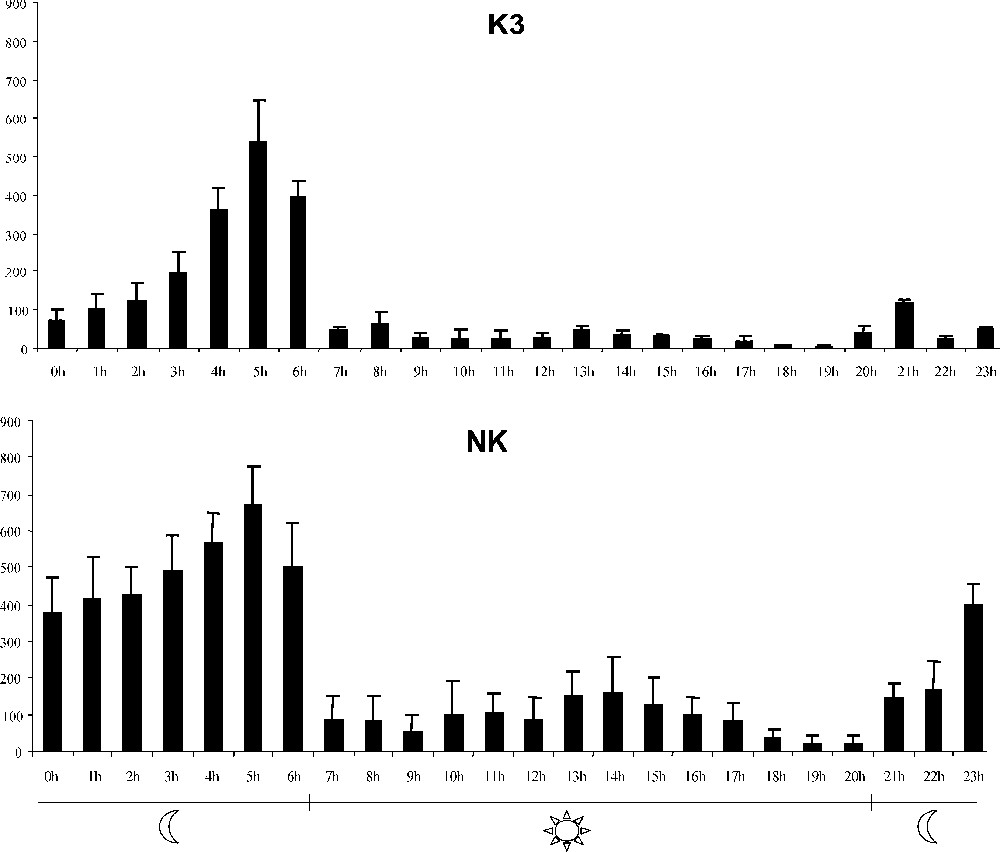
Daily organization of singing activity in a group of mature Japanese quail brothers (K3; N=5) and in a group of non-related males (NK; N=6). Data for all the singing males were pooled. Hourly mean activity was calculated over consecutive days.
First, during these two hours, the respective proportions of song activity of the different males in the chorus were noted for five days (days 1, 2, 3, 4 and 7), for the five groups. Second, the most actively singing male was removed on day 8 from two groups with similar numbers of males (K3: and NK: ), and similar general singing activities (the quantities of song of the main singers were approximately the same and the song profiles of the other males were similar in these two groups), and the individual singing activities of the remaining birds were recorded for four days (days 9, 10, 12 and 14). Non-parametrical statistical tests estimated significance of data (Statview SAS, 92-98). Spearman tests calculated synchronization correlations between two males [9]. The Spearman null hypothesis was that two variables (two males) were independent and the alternative hypothesis was that the rank values of two variables were correlated (two males were synchronized). The rho values vary from −1 (high rank of one variable correlated with low rank of the other) to +1 (high rank of one variable correlated with high rank of the other), 0 indicates the absence of correlation between variables.
3 Results
3.1 First phase
During the two hours preceding lights-on, and for the five days of recording, total singing activities for the males of the five groups were very intense, as for example more than 1000 songs were recorded in groups K3 ( songs; males), NK (; ) and K4 (; ). Mean activity of the other two groups approximated only 500 songs (K1: ; ) (K2: ; ). So, on the whole, this suggests that numbers of singers and numbers of songs are correlated. However, the two groups with five brothers, K2 and K3, differed significantly, group K3 males sang more (Mann–Whitney U test, ).
Inter-individual differences were observed in all groups (Fig. 2; Table 1). One male sang significantly more than all the other males in his group, in four of the five groups (Mann–Whitney U test, K1 ; K3 ; K4 ; NK ). In the last group, K2, two males sang significantly less than the three others (Table 1). Males in a group could be ranked in a regular hierarchical order (Fig. 2), but only some differences between two successive males were significant (Table 1). These differences were stable over the five records, from day 1 to day 7 (ANOVA for all the groups). The proportion of song activity for the most active male in each group decreased with increasing numbers of singers: from 44% of the total singing in the group of four singers (K1), to 34% and 35% in the two groups of five singers (K2 and K3), 34% in the group of six singers (NK) and finally to 21% in the group of seven singers (K4). In the four groups with one significantly more active male (K1, K3, NK and K4), the respective proportions of songs emitted by this male in the choruses were inversely correlated with the numbers of singers (Fig. 3). Thus, during the two-hour recording sessions, the most active male in the group of four (K1) emitted 44% of approximately 500 songs, i.e. approximately 220, and that the most active male in the group of seven (K4) emitted 21% of approximately 1700 songs, i.e. approximately 350.
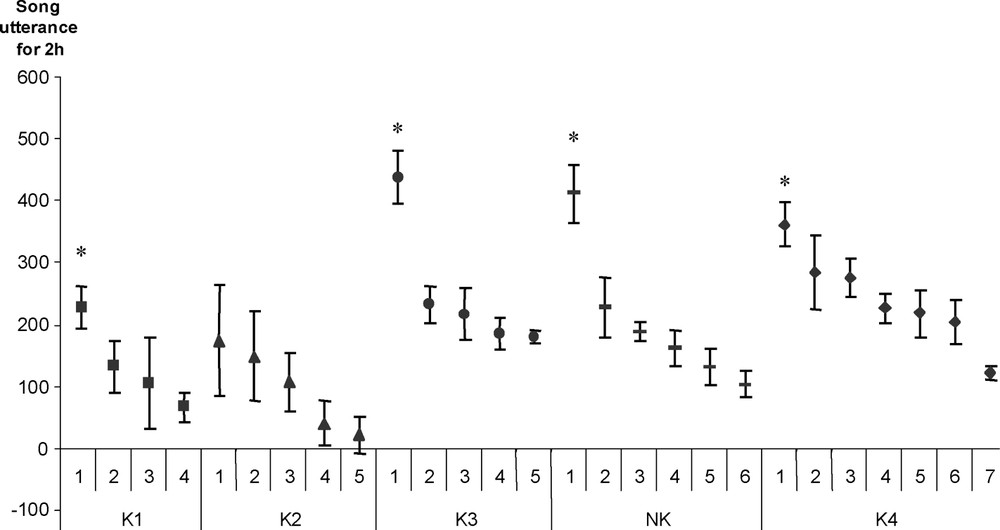
Individual singing data. During the two hours preceding lights-on and for the five groups, number of songs emitted by each male, in each group. *: the activity of this male was significantly higher than that of all the other males (Mann–Whitney, p<0.02).
Inter-individual differences of singing activities for the five groups tested. K1, K3, NK and K4: kin-related males, NK: unrelated males. Bold type: significant values at p<0.05
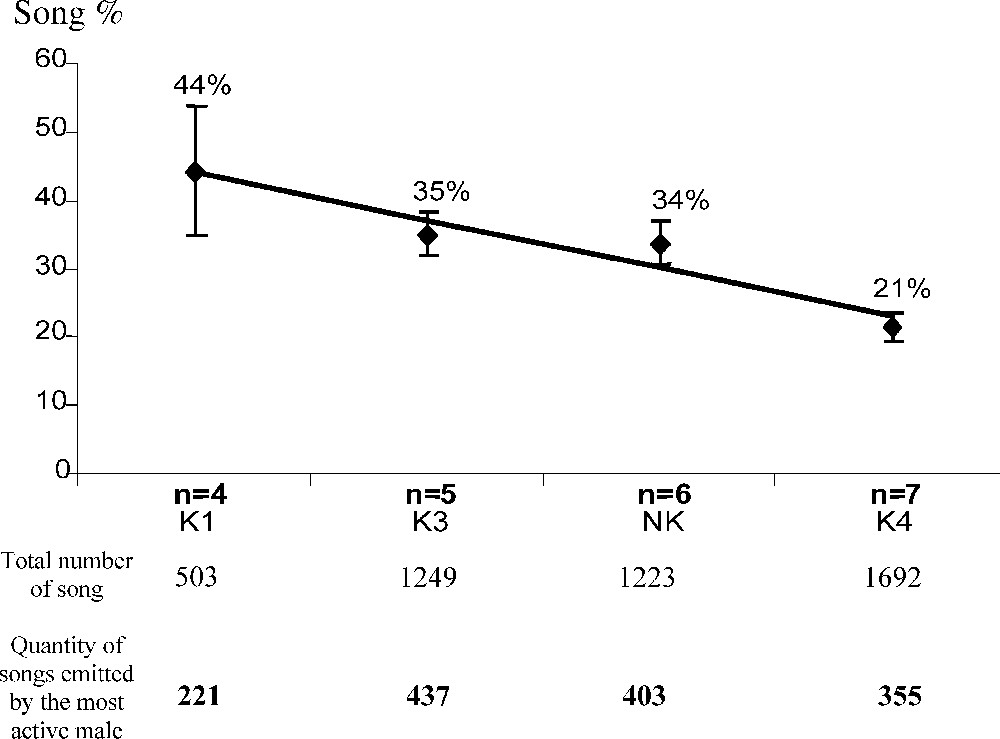
Singing activity (in % of the chorus) by the most active male in relation to the number of singers in the four groups K1, K3, NK, and K4, where one male sang more intensively (y=−6.9593x+50.973).
3.2 Second phase
This part included a preliminary study of the impact of the removal of a male, to compare with the departure of a male from a group in the wild. To test the importance of affiliation on the stability of a group we compared song emissions between a group of affiliated (K3) and a group of non-affiliated (NK) males. The groups of kin and non-kin birds reacted differently following the removal of the most active males.
In the group of kin-related males (K3), the total singing activity during the two hours preceding lights-on, initially songs for five brothers, dropped to for the four remaining brothers. This 31% decrease corresponds to the individual proportion of song of the most active brother (35%, see above) that was removed. Thus, removal did not modify significantly individual singing activities of the four remaining males (Fig. 4).
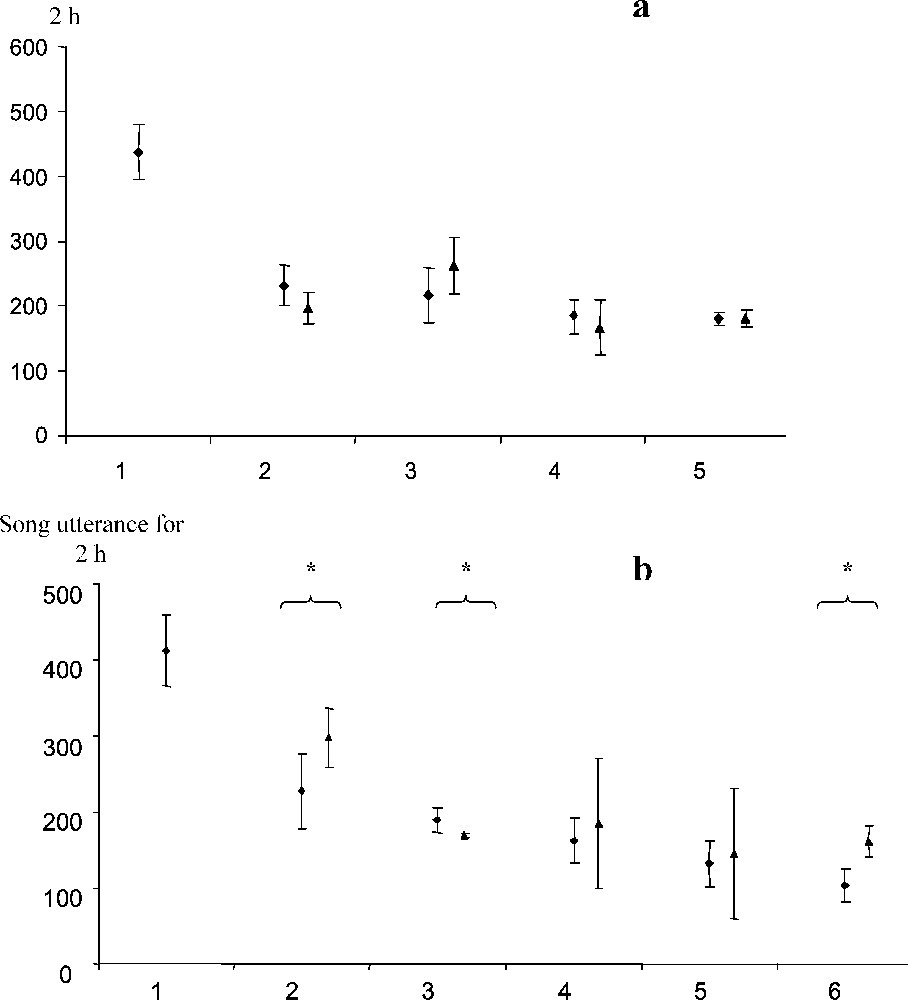
Effect of removal of the most active singing male on singing by the other males in the group, for two groups of males. (a) Group of kin-related quail, K3, (b) group of non-related quail, NK. ♦ Before removal of the most active male. ▴ After removal of the most active male. * Significant difference between before and after removal, Mann–Whitney U test (p<0.05).
In the group of six unrelated males (NK), the total singing activity was initially songs during the two hours preceding lights-on. The total activity of the five remaining males decreased to . This 19% decrease is less than the relative part (33%) of the most active male when he was present in the chorus. Standard deviation for the four records after the removal of the most active male (days 9, 10, 12 and 14) was nearly double that for the five previous records (days 1, 2, 3, 4 and 7), when all the males were present. Moreover, the singing activity of individuals varied differently (Fig. 4). Two of the males sang significantly more than during the previous period, one male sang less (Mann–Whitney U test ) and the two other males increased their activities, but not regularly over the four days of recording.
The levels of inter-individual synchronization of singing activity for 5-minute periods (yielding observations for two hours) were calculated before and after removal of the most active male for the two groups. Inter-individual synchronizations were calculated using a Spearman test of correlation.
In the group of five brothers (K3), only the two less active birds were significantly correlated before (, ) and after (, ) removal. In the group of the six unrelated males (NK), the singing activity of the less active male was correlated before removal with that of three others (in the three cases, ; ), but not with that of the most active male. Furthermore, we found significant inter-individual correlations in three other comparisons between the other males (with intermediate emission levels). After removal of the most active male only two of the six pairs remained synchronized. Coefficients of inter-individual synchronization in the kin group did not differ significantly before and after removal of the most active male (Wilcoxon test). However, inter-individual synchronization coefficients decreased significantly between day 7 and day 14 (Wilcoxon test, ) and between day 9 and day 14 () in the group of unrelated birds.
4 Conclusion and discussion
This study of quail males in reproduction groups revealed inter-individual differences implying organization of the singing activities in a group. The daily organization of general singing activity presents a marked peak at dawn. Recordings of the activities of males for two hours during this peak revealed that, in four out of the five experimental groups, one male sang significantly more than all the other males in his group and the quantities of song emitted differed among males in a group, varying gradually from the most frequent singer to individuals that sing much less. This was true whatever the number of males in the chorus and relationships (brothers or not) within the group. During the two hours preceding lights-on, the numbers of songs increased, depending on the numbers of males present, from around 500 songs in a group of four males, to approximately 1700 songs in a group of seven males. The respective proportions of songs emitted by the most active male decreased when the number of males in the group increased. These differences remained stable over the seven observation days. Thus, the organization of a quail chorus group is characterized by the presence of a song ‘master’.
One functional value associated with quail choruses is the stimulation of sexually developing females. These females present privileged phases of sensitivity, in particular between 7 and 11 h after dusk, a period that corresponds to the dawn peak of singing activity under a spring photoperiod [10]. Our results show that the larger the group of males, the stronger their vocal stimulation for females. Previous results stressed the importance of the quantity (frequency) as well the quality (different voices) of vocal stimulation [11]. A male would benefit from being in a chorus group with several males because the access to sexually mature females would be enhanced. The question remains as to why there is a song ‘master’ in a chorus and what is the functional value of this organization? Singing frequency is an individual characteristic and one male with a high-singing-frequency phenotype is found in most groups This structure appears to be inherent to Japanese quail choruses. Detailed monitoring of chorus ontogeny is necessary to understand the establishment of this structure.
Comparisons of inter-individual synchronization levels of singing activities suggested that the ‘master’ singer did not synchronize, over 5-min periods, the activities of the others, especially in the group of five brothers. The different males can sing all the time during the 2-hour observations. Thus, stimulation of females is intense and continuous during the two hours preceding lights-on and we suppose that in this case males cooperate by alternating song production. However, at the entire day scale, vocal activities of all the males were well synchronized.
Nevertheless, the intense activity of one male always stands out in the general activity of a chorus. This male can thus draw the attention of females to himself and therefore favour his mating. In many birds and specially those with male choruses and/or leks, female mate choice is now well known [12] and intense vocal activity could be an honest indication of male quality for females [13,14].
As shown in this preliminary study, the removal of the most vocally active male seemed to have more important consequences in the group of unrelated males. In this group, several males modified their singing frequencies significantly from day to day and previous inter-individual synchronizations were lost. On the contrary, the group of the remaining four brothers seemed to retain its initial characteristics. After the removal, interactions among the remaining brothers seemed unchanged, whereas, the vocal hierarchy appeared challenged in the group of unrelated males. However, this study included only two groups and detailed replicates are required to confirm this conclusion.
Quail choruses in the field probably include kin-related males, as their vocalization themes are similar [4]. This assumption is supported by genetic analyses [15]. Owing to the relatively short survival rates of quail in the wild, the possibility that sons encounter their fathers is low. That is why it is very likely that birds in choruses in the wild are brothers. Kin-related males of several lekking bird species aggregate, and the established males can gain direct and indirect benefits [16].
Our preliminary study stressed the great stability of the chorus group of brothers. Avian kin recognition has been extensively studied during the past two decades [17], but most experiments deal with parent-offspring recognition, a first step to preferential allocation of help to nearest kin, as in cooperative breeding birds. Bateson [18] showed that Japanese quail preferred to mate with first cousins. Therefore, according to our findings, the chorus structure is likely to support a kind of cooperation between chorusing male quails. Our preliminary result with non-kin birds indicates that this might be especially true when males are kin-related. However, further investigations are clearly needed to confirm this fact.
Acknowledgments
We are grateful to Dr Ann Cloarec for the corrections of the English version of the manuscript.