

1 Introduction
The role of the landscape in the distribution of the animal populations has been relatively little studied yet; it largely depends on the biology of the species [1], particularly in relation to its diet (specialist or opportunist feeders) and capacity to move as well as to the landscape variables taken into account [2]. In addition, the dynamics of a population plays a potentially important role because the natural resources, and consequently the distribution and the structure of the habitats, intervene in the speed of population limitation and thus in the capacity to produce emigrants allowing geographical expansion [3–6]. However, these elements are seldom taken into account in the literature, because it is generally considered that a species responds unequivocally to the same landscape characteristics.
The Grey Heron (Ardea cinerea) was probably present throughout France at the beginning of the 19th century, but became almost extinct at the beginning of the following century due to the persecution by fish-farmers and anglers [7,8]. Only two significant breeding colonies remained in eastern and western France. From these two refuges, the species showed a very strong expansion, both numerically and geographically, involving a progressive recovering of its initial distribution. However, this recovery varied depending on the areas, due to both the direct action of man and the structure of the hydrographical network. The specific structures of the landscape in each area and the regional history of the heron populations could influence the strategies of breeding colonies' distribution. In western France, the ‘mother’ colony of Grand-Lieu led to the installation of ‘daughter’ colonies in willow marshes for a long time, before the new colony of Guérande in the Loire-Atlantique ‘département’ (1966) generated a secondary lineage of coastal colonies settling in coniferous trees near the coast as far north as Finistère [9]. At this regional scale, the strategy observed in eastern France appeared to be homogeneous relative to the elements of the hydrographical network that influenced the distribution of the colonies in comparison to the western situation [9]. However, the relationship of a species with its landscape may differ according to the scale of analysis [2]. The aim of this study was to analyse more precisely the role played by the hydrographical network structure in eastern France alone, at a lower scale of analysis and without the influence of the western population data, and within the conceptual framework of landscape ecology, to determine whether there were differences in local strategies of spatial use. This paper complements the previous one [9] comparing the eastern and western French refuge areas that were taken to be representative of the French situation. These two old populations are still most important in this country [4], and constitute the largest European population of this species.
2 Materials and methods
2.1 Species data set
We used 69 colonies in the eastern part of France (Fig. 1), counted during the National Heronries Census of 1994 [4]. The method used for this census was similar to that for the preceding national censuses of 1974, 1981, 1985, and 1989 [4]. In theory, all the nests occupied during the reproduction of 1994 were listed between May and July. We can consider that this census was quasi-exhaustive in the areas retained for this work, and that the distribution of the colonies accurately reproduces the reality of the occupation by birds.
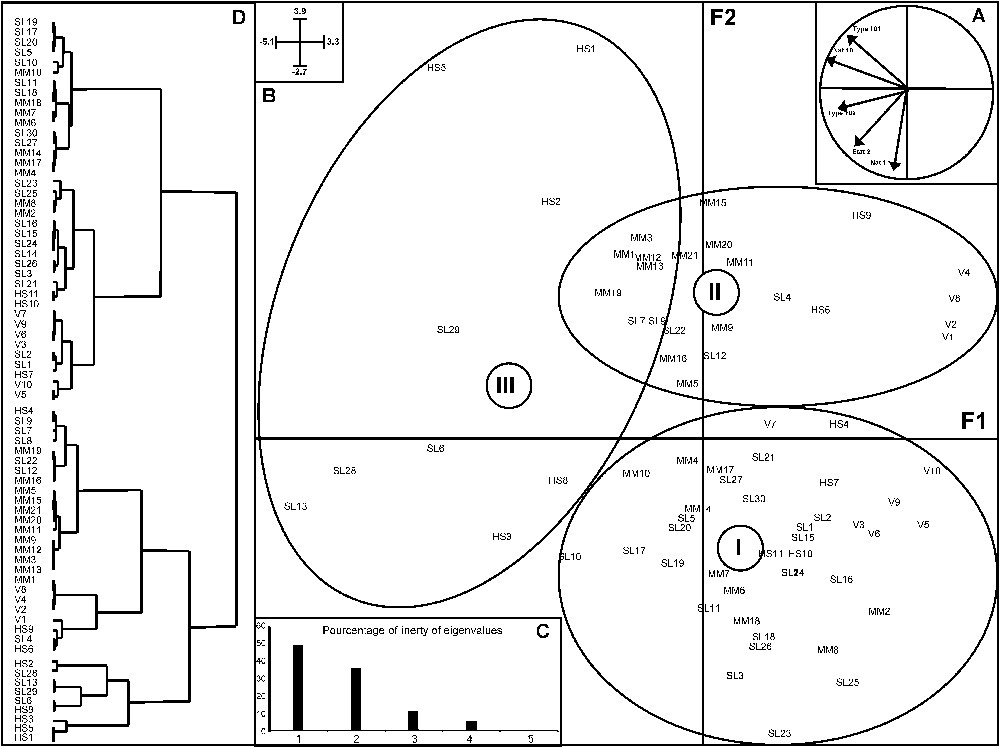
Scatter diagram of PCA axes 1 and 2 of the hydrographical variables for the colony sites on the 25-km scale. A: Correlation circle with variables. B: Histogram of eigenvalues. C: Scatter plot of the sites. D: Dendrogram of the site ellipses (I, II, III), distinguishing the colony sites according to the hierarchical ascendant classification (Ward). The number indicated on the ellipse represents its centre of gravity. Coordinates of the axes are on the top-left corner of the figure.
2.2 Environmental data set
We took into account various descriptors of the hydrographical network, separating in particular the liner network and the surface one. The linear network corresponded to all the flows of permanent and temporary water, whereas the surface one corresponds to the ponds and marshes.
These data were extracted from BD-Carthage version 1.1, which is a numerical file designed jointly by the Department of the Environment and the National Geographical Institute (IGN) and represents the entire hydrographical network, both linear and surface. It describes the nature (natural, channel, ditch, estuary), the state (permanent or temporary) and the width of the rivers. It also describes the ponds according to their nature (permanent or temporary freshwater or seawater) and their type (marsh, coastal, etc.). From these variables, we largely retained a set of 12, which were the most important in this part of France according to a previous study [9]. However, according to a preliminary analysis here, we retained the five that were the most important and were not correlated. Thus, we only took into account the variables describing the natural hydrographical network (Nat1), the permanent areas of freshwater (Nat10), the ponds (Typ102), the biggest rivers (Typ101), and the temporary rivers (Etat2).
2.3 Statistical analysis
Firstly, we measured the lengths and areas of the various types defined above, in a radius of 25 km around the colonies. This radius is an accurate representation of the potential feeding area of this species [7–9].
Secondly, a Principal-Component Analysis (PCA) was carried out on these variables, followed by a hierarchical ascendant classification using the Ward method, which maximizes the dissimilarity between the colonies, in order to classify different groups of colonies according to the hydrographical structure of their foraging area. Analyses and graphics were performed using ADE-4 Software [10]. Next, a one-way ANOVA [11] was carried out to compare the size of the colonies in each group.
3 Results
The distribution of colonies appeared to be strongly dependent on the hydrological network. The PCA (Fig. 1) distinguished three groups of colonies. The F1 axis translated a gradient of pond areas, while the F2 axis opposed the very large rivers to the small rivers. The group III was distinguished from the others according to axis 1, whereas the two others were only divided according to axis 2. The geographical position of colonies did not really determine the groups to which they belong (Fig. 2). This location is explained better by the structure of the hydrological network. However, the one-way ANOVA did not show any significant difference in the size of colonies between groups (Fig. 3).
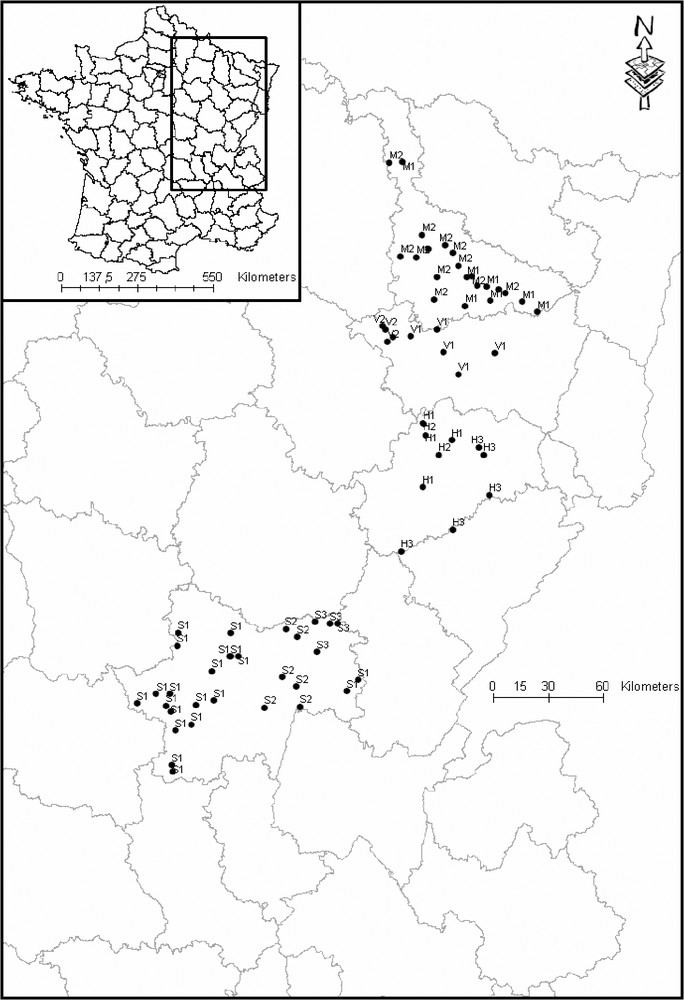
Study area and colonies location. Label indicates the number of the group to which each colony belongs.
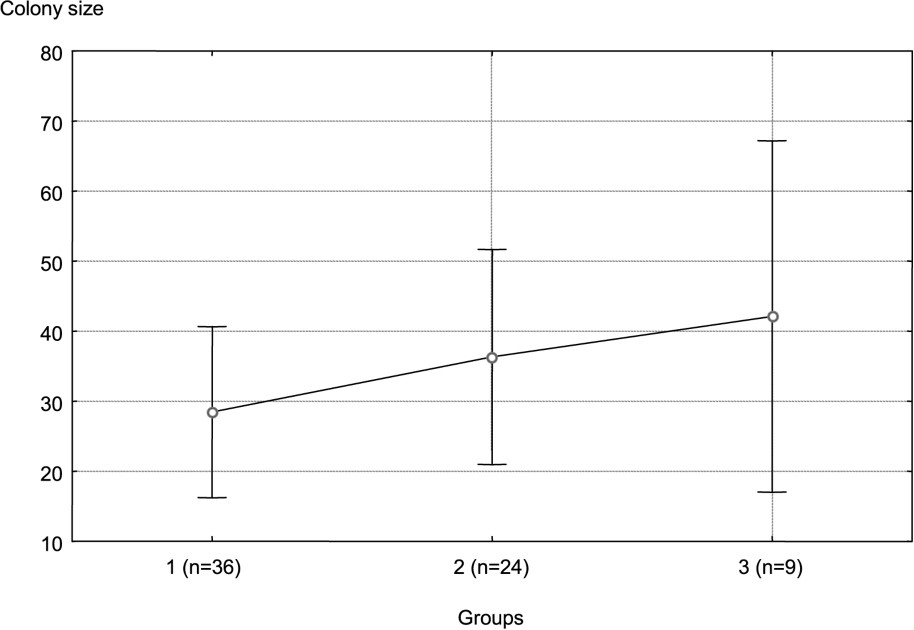
Mean colony size of each group. Error bars represent ± one standard deviation from the mean.
3.1 Group I
The 36 colonies of group I were distributed over the whole study area. However, their location was influenced by the same type of hydrographical structure: a dense network of small (Nat1) and frequently intermittent rivers (Etat2).
3.2 Group II
The 24 colonies of group II were also distributed throughout the whole study area, even though most of them were located in the northern part. The colonies of this group were opposed to those of the second group according to axis 2. Their location are influenced, not by the small rivers but, on the contrary, by the large ones (Nat101), with a low density.
3.3 Group III
The last group included nine colonies, which were located along the same river. Their location was influenced by the presence of ponds (Type 102), which were more (Nat10) or less (Etat2) permanent.
4 Discussion
While a previous study [9] conducted at a national scale did not exhibit different patterns of the use of landscape characteristics in eastern France, as opposed to western France, where geographical and historical lineages were linked to the main landscape characteristics (littoral, estuarine, inland waters), the present study highlights less evident relationships with hydrological characteristics, confirming the role of the scale of analysis, well known in landscape ecology. As in the previous study [9], this study was carried out only at breeding colony locations and not at the feeding sites of herons, which forage primarily on thousands of individual food territories [3,12,13]. During the breeding period, individuals mainly eat aquatic prey [7,14–16], which are not really represented by the landscape of the colony itself, but colonies are located according to their feeding sites as well as the security conditions present at the site [7,13]. Thus, the site of the colony is not randomly selected. Ardeids prefer locations where there are large areas of wetlands or long lengths of banks [7,17,18]. The specific structures of landscape of each area could influence the strategies of breeding colony distribution. In Brittany, colonies of grey herons showed three strategies of habitat use: estuary marshes, littoral (shore) and continental waters, each one being characterized by a different hydrological landscape. Regarding their location, the colonies located in the continental area of Brittany were influenced by the same overall ecogeographical variables as those influencing the presence of colonies in the east of France. In the latter, no heterogeneity of response was found in the global analysis [9], in spite of the extent of the zone studied. However, the present study, made at the scale of eastern France alone, allowed statistically significant differences of landscape to be highlighted, even though all colonies used the same kind of habitat for establishing their nest (mainly broad-leaved trees), in contrast to western France, where the three groups of colonies used different kinds of trees (willows in marshes for the estuary group, conifers for the littoral group, broad-leaved trees for the continental group), forming three ecological lineages of colonies, probably depending on the habitat where the birds were born. In eastern France, we distinguished three types of exploited areas. Some colonies settled in part of the study area with a landscape characterized by a network of small intermittent rivers. These elements of hydrographical landscape provide the birds with shallow water, in which they can easily forage. Colonies of this type represented about half of the colonies studied. The second type of colonies was located near large rivers, in landscape where the density of river was low. In this environment, grey herons foraged along the bank of the river, where the water depth enabled them to forage. This type represents, as the first, nearly half of the studied colonies. The last group was more restricted, and included only nine colonies, which were located along the same river. The location of these colonies was influenced by the number of ponds and their area. For this group, we distinguished a sub-group of colonies that were more influenced by non-permanent ponds, and a second sub-group, for which permanent ponds mostly contributed to the location of colonies.
These different types of habitat used by grey herons have no significant effect on the size of the colonies: they seem to be all of equal environmental quality. However, colony size tends to increase from the first to the third group. We can explain that by the density of rivers. Effectively, there is a gradient of increasing river density from group 1 to group 3. Thence, when the density of small rivers is high, feeding territories are uniformly distributed, and the colonies can be located at the closest to these habitats, a situation that favours the multiplication of small colonies. On the contrary, the less important is river density: the most-feeding territories will be distant from each other.
The relation between the size of the colonies and the structure of the landscape needs more investigations, particularly by taking into account the fitting of the various elements of the hydrographical network within the landscape, and by discriminating the water depth, this species exploiting only shallow water.
Acknowledgements
This study was supported by grants from the French Ministry of the Environment and SESLG. We thank AOMSL, GNFC, and LPO of Lorraine who participated in heronries census. We also thank anonymous referees for helpful comments on a previous version of the manuscript.