

Abridged English version
Usually, one talks of heterostyly when the individuals of the same species are distinguishable by the relative length of the styles of their hermaphrodite flowers (long and short in distyly; long, medium and short in tristyly). In all cases, each individual bears only one type of flower. Heterostyly is generally accompanied by a polymorphism of pollen and of the papillary cells of stigmas.
Observations of Atriplex halimus wild populations from different Mediterranean basin regions have shown style length homogeneity, as well at the individual level as between individuals. Only two Tunisian populations (Kairouan and Tadjerouine) have shown heterogeneity of style length, and a bracteole trimorphism of female flowers.
From the observation of intra-individual variability of style length in Atriplex halimus, we have been led to undertake a statistical study of this material to determine if it consisted in a continuous variability or a discontinuous one, consisting in an intra-individual tristyly.
The plants studied belong to two clones representing the two populations of Tunisia (Tadjerouine, T1 and Kairouan, T2). Three morphotypes (a, b, c) were recognized, without any confusion, depending on the form and the degree of the bracteole union (characters that are used in key determinations for Amaranthaceae) and on style length.
Style lengths were measured on:
- – three samples of flowers, for each morphotype (a, b, c) of the two clones;
- – three samples of each morphotype (a, b, c), on Tadjerouine clone;
- – randomly, on one sample (Tadjerouine).
In order to characterize the papillary cells of the long and short styles, only the longest styles of (a) and the shortest of (c) morphotypes were considered.
The pollen size was measured on the two clones.
All the results reflect a non-normality of the different distributions. The results were the following:
- – for each morphotype (a, b, c), the distribution of style length presents three major peaks (long, medium, short), each morphotype characterized by its own mean and dispersion. This distribution shows that tristyly is expressed in each bracteolate flower morphotype;
- – random sampling, independently of morphotypes, shows a style length distribution in the Tadjerouine clone, with nine very distinct peaks, which could be interpreted as a combination of tristyly with the three morphotypes (a, b, c);
- – comparing bracteoles of the morphotype (a) with those of Atriplex nummularia, collected in the same region, one finds a very remarkable resemblance, not only in the form and degree of union of bracteoles, but also in the style length. A. nummularia is known as being long-styled, as observed on local samples. Similarly, comparing the bracteoles of the morphotype (c) with those classically described for A. halimus, one finds again a resemblance in form and degree of union, but also in the shortest styles. Moreover, A. nummularia pollen corresponds to the large pollen of our results, as to its size and density of apertures. A. halimus pollen, as verified on a morphotype (c) population, collected in Tunisia, is similar to the small pollen described in the results.
Though unusual, the intra-individual heterostyly presents other characters related to the phenomenon, i.e. a polymorphism of the pollen and of the stigma papillary cells. Pollen lodges, in T1 clone male flowers, always contain a mixture of three types of pollen, characterized by size, colour and sculpture.
Concerning the manifestation of this heterostyly, one can mention:
- – that so far, it is the only case of heterostyly observed in the species, genus, and more generally in the Amaranthaceae family, to which the Atriplex genus belongs;
- – that styles of different sizes are found in female flowers, even though, in some instances, this heterostyly can affect hermaphrodite flowers;
- – that, whereas characters of floral sterile pieces are invariable no matter the style size in classical heterostyly, we can distinguish here three floral morphotypes of heterostyly, depending on the form, size and degree of bracteole union, each one displaying a tristyly;
- – that this heterostyly is intra-individual, with nine floral morphotypes per individual, because of the combination of three style lengths and three bracteole morphotypes.
Is there any advantage for the species?
In the case of this material, the classical notion of auto-incompatibility cannot be applied stricto sensu; in the same individual, small pollen and large pollen grains are produced, therefore, this is a situation of autocompatibility stricto sensu. Due to this autocompatibility, individuals give rise to an abundant progeny in a given area. This device is of a great importance, since the individual behaves as a population by itself.
This intra-individual heterostyly might be in relation with a hybridisation between two tetraploid species: A. halimus, a monoecious, short-styled plant, and A. nummularia, a dioecious long-styled plant species, for which heterostyly has never been reported up to now.
The hybridisation hypothesis seems to be confirmed by artificial cross breeding between A. halimus and A. nummularia. Out of 100 akenes from these crossings, 12 contained well-formed seeds, which germinated. Considering that, in the areas studied, these two species have partially overlapping flowering periods, such a proportion is quite significant. In traditional hybridisation, the two species are represented by two individuals of different genotypes, whereas in our material, one individual has one genotype, but with the phenotypes (a) and (c) of the two parent species. All this occurs as if one of our plants is equivalent to a population of (a) and (c), with intermediary forms (b).
The existence of intermediary forms could suggest an introgression, but, as regards a possible hybridisation, we are led to conclude to an equivalence between a plant and a population.
All that precedes suggests the existence of an intra-individual genetic variation. However, other authors have pointed out that a plant can behave as an assembly of different phenotypes and genotypes of the same species. However, in our material, the plant looks like an assembly of phenotypes and genotypes of two different species, A. halimus, and A. nummularia. Actually, among the nine categories of female flowers observed on the same plant, the phenotypes (a) and (c) correspond respectively to those of Atriplex nummularia and A. halimus, to which seven intermediate phenotypes are added.
In this case, we cannot put forward some somatic mutations in order to explain the expression in A. halimus, of characters (bracteoles, style, pollen) of A. nummularia. It is highly improbable that such an event occurs on a large scale at the plant level.
Therefore, A. halimus is an original species, since it reveals characters, in the reproductive apparatus, which modify the notion of individual, bringing new problems to solve to biologists and geneticists.
1 Introduction
Jusqu'à présent, l'hétérostylie était décrite chez des individus d'une même espèce dont les fleurs hermaphrodites diffèrent par la longueur relative de leurs styles (longs ou courts : distylie ; longs, moyens ou courts : tristylie). Cette hétérostylie est généralement accompagnée d'un polymorphisme des papilles stigmatiques et du pollen. Dans tous les cas, chaque individu ne porte qu'un seul type de fleur. Cette hétérostylie est observée au moins dans 25 familles d'Angiospermes [1].
Les observations de populations sauvages d'Atriplex halimus dans différentes régions du Bassin méditerranéen (Algérie : 2 ; Syrie : 2 ; Espagne : 5 ; France : 5 ; Maroc : 7 ; Tunisie : 12) ont montré une homogénéité de la taille des styles, tant à l'échelle de l'individu qu'entre les individus de chaque population. Seules deux des populations tunisiennes (Kairouan et Tadjerouine) ont montré une hétérogénéité de la taille des styles, mais aussi, contrairement aux autres, un trimorphisme des bractéoles de leurs fleurs femelles.
Les observations de ces deux populations nous ont fait penser que nous étions en présence d'une hétérostylie intra-individuelle, contrairement à l'hétérostylie interindividuelle classique. L'importance théorique d'une hétérostylie intra-individuelle est telle, que nous avons conduit une étude statistique de ce matériel particulier, afin de déterminer si, à l'échelle d'un individu, on est en présence d'une variabilité continue ou discontinue.
2 Matériel et méthodes
2.1 Matériel végétal
Nos observations et mesures sont de deux ordres : taille des styles et des papilles stigmatiques des fleurs femelles, d'une part, taille et caractères du pollen des fleurs mâles, d'autre part.
Des semences sont récoltées en Tunisie sur deux populations d'A. halimus tétraploïdes (
2.2 Trimorphie des fleurs femelles
Trois morphotypes (a, b et c) sont reconnus, d'après la forme et le degré de soudure des bractéoles (soudées dans leur moitié inférieure a et b, ou uniquement à la base c ; Fig. 1). Ces trois morphotypes sont parfaitement distincts, car il n'existe pas de variabilité continue de la forme et du degré de soudure des bractéoles.

Bractéoles des fleurs femelles, pour chacun des morphotypes a, b, c, d'un même individu d'Atriplex halimus. Les zones de soudure sont représentées en noir, les flèches indiquent leurs limites distales (échelle × 8).
2.3 Biométrie des styles
Afin d'étudier l'éventuelle discontinuité de la variation intra-individuelle de la longueur des styles, trois approches sont adoptées. Nos observations se sont limitées, dans la plupart des cas, à un seul individu, et ce pour plusieurs raisons : d'une part, l'extrême difficulté des observations (compacité des glomérules floraux, petitesse des fleurs, abondance des trichomes) ; d'autre part, la nécessité d'avoir des effectifs importants.
- • Pour rechercher une possible corrélation entre les trois morphotypes (a, b et c) et la taille des styles au sein des deux clones (T1 et T2) cultivés à Porquerolles, on constitue trois lots de 50 fleurs (10 fleurs par individu) pour chaque morphotype, soit 300 fleurs en tout.
- • Pour étudier la distribution des longueurs de styles (di- ou tristylie) au sein de chaque morphotype, on constitue, à partir d'un individu du clone T1, trois lots de 100 fleurs de chaque morphotype, soit 300 fleurs en tout.
- • Afin d'étudier précisément cette distribution, indépendamment des morphotypes, et d'en conclure ou non à l'éventuelle discontinuité du caractère étudié, un prélèvement aléatoire de 795 fleurs est fait sur un individu (T1) cultivé en conditions de serre.
2.4 Papilles stigmatiques
Pour caractériser les cellules papillaires, on s'est intéressé aux styles les plus longs des (a) et les plus courts des (c). On effectue, sur chacun des cinq individus des deux clones T1 et T2 de Porquerolles, un tri de cinq fleurs de type (a) à styles longs (entre 500 et 700 μm) et de cinq fleurs de type (c) à styles courts (entre 100 et 300 μm).
2.5 Biométrie du pollen
Le diamètre de 1000 grains est évalué sur 10 fleurs mâles, à raison de 100 grains par fleur, pour l'individu à l'origine du clone T1. Une coloration à l'acétocarmin est effectuée pour contrôler la viabilité des grains.
2.6 Analyse statistique
Elle est faite avec les logiciels SAS et Minitab (version 13 pour Windows 2000, Minitab Inc. USA). Une ANOVA est réalisée pour rechercher d'éventuelles interactions (longueur des styles et des cellules papillaires).
3 Résultats
3.1 Longueur des styles
- •
Les résultats obtenus à la suite d'observations faites sur les deux clones cultivés à Porquerolles, sur la base de trois lots de 50 fleurs pour chaque morphotype (a, b, c), montrent que (Figs. 2 et 3, Tableau 1), pour les deux clones, les fleurs de type (a) ont des styles qui sont, en moyenne, plus longs (
- •
La distribution des longueurs, en unités arbitraires, de styles de trois lots de 100 fleurs de chaque morphotype, pour un individu du clone T1 (Fig. 4), établit que chaque morphotype présente trois pics (styles longs-L, moyens-M, courts-C) se distinguant nettement d'un ensemble de pics mineurs :
- • Le prélèvement aléatoire de 795 fleurs, réalisé sur un individu du clone T1 cultivé en conditions de serre, montre (Fig. 5) que la distribution des longueurs de styles fait apparaître neuf pics, séparés par de très nettes discontinuités. Ces pics sont entourés de pics mineurs. C'est encore le reflet d'une non-normalité de la distribution. Dans ce prélèvement aléatoire, où le biais introduit par le tri (a, b, c) est évité, tout porte à croire que les neuf pics observés sont le résultat, à l'échelle de l'individu, de la tristylie des trois morphotypes.

Tristylie et polymorphisme pollinique au sein d'un même individu d'Atriplex halimus (MEB). De haut en bas, styles, papilles stigmatiques et pollen. Style long (L), style moyen (M), style court (C) et cellules papillaires stigmatiques d'un style long (a) et d'un style court (b). Gros et petit grains de pollen et détail de leur exine (voir également le Tableau 2), (stg : stigmates).

Longueurs en millimètres des styles chez les trois morphotypes de fleurs femelles d'Atriplex halimus. Clone T1 (a1, b1, c1) et clone T2 (a2, b2, c2). Représentation des différents quartiles, des valeurs maximales et minimales (traits verticaux) et des valeurs aberrantes (•).
Moyennes et médianes des longueurs en millimètres des styles chez les différents morphotypes pour les deux clones T1 (a1, b1, c1) et T2 (a2, b2, c2) d'Atriplex halimus
| Effectif | Moyenne | Médiane | ||
| Clones | T1 | 150 | 0,028 | 0,03 |
| T2 | 150 | 0,028 | 0,03 | |
| Fleurs | a1 | 50 | 0,038 | 0,03 |
| a2 | 50 | 0,035 | 0,03 | |
| b1 | 50 | 0,029 | 0,03 | |
| b2 | 50 | 0,031 | 0,03 | |
| c1 | 50 | 0,016 | 0,015 | |
| c2 | 50 | 0,017 | 0,02 |

Longueurs des styles en unités arbitraires (ua) selon les morphotypes floraux (a, b, c). Les trois principaux pics de chaque morphotype : styles longs (La, Lb, Lc), moyens (Ma, Mb, Mc), courts (Ca, Cb, Cc). Les barres d'erreur, au risque 5%, sont portées sur chaque pic.
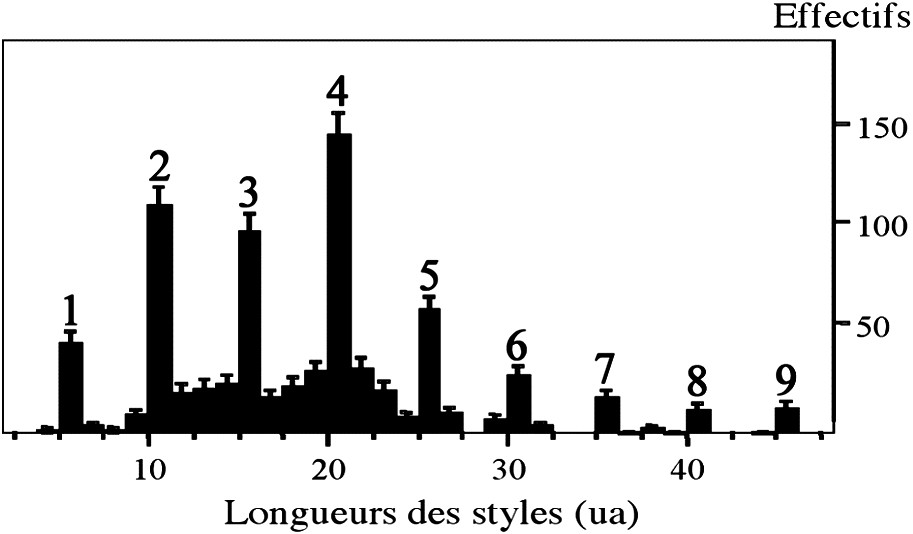
Distribution des longueurs de styles en unités arbitraires (ua) de 795 fleurs femelles à deux bractéoles, prélevées aléatoirement sur un individu du clone T1 cultivé en conditions de serre. Les pics se distinguant du bruit de fond sont numérotés de 1 à 9. Les barres d'erreur, au risque 5%, sont portées sur chaque pic.
3.2 Longueur et largeur des papilles stigmatiques
Les papilles stigmatiques sont unicellulaires. Leur taille moyenne diffère selon la longueur du style (Fig. 2). Pour chaque clone, les cellules papillaires les plus longues, 62,30 μm (T1) et 102,3 μm (T2), sont significativement liées aux styles longs, et les cellules papillaires courtes 37,60 μm (T1) et 71,83 μm (T2) aux styles courts, avec
3.3 Pollen
Les loges polliniques des anthères des fleurs mâles du clone T1 contiennent toujours trois types de pollen en mélange, se distinguant par leur taille, leur couleur, mais aussi par leur sculpture (Tableau 2). En revanche, ils sont tous sphériques. L'analyse de la variance montre une tendance très marquée en faveur d'un trimorphisme pollinique, avec
Caractéristiques de l'exine des trois types de grains de pollen d'Atriplex halimus
| Pollen | I | II | III | IV |
| Petit | 17 | circulaires | sphérique | 4,5 à 8 |
| (0,9 à 1) | ||||
| Intermédiaire | 7 à 8 | circulaires | étoilée | 1,1 |
| (1,2 à 3,5) | ||||
| Gros | >8 | circulaires | étoilée | 2 |
| (1,2 à 1,3) |
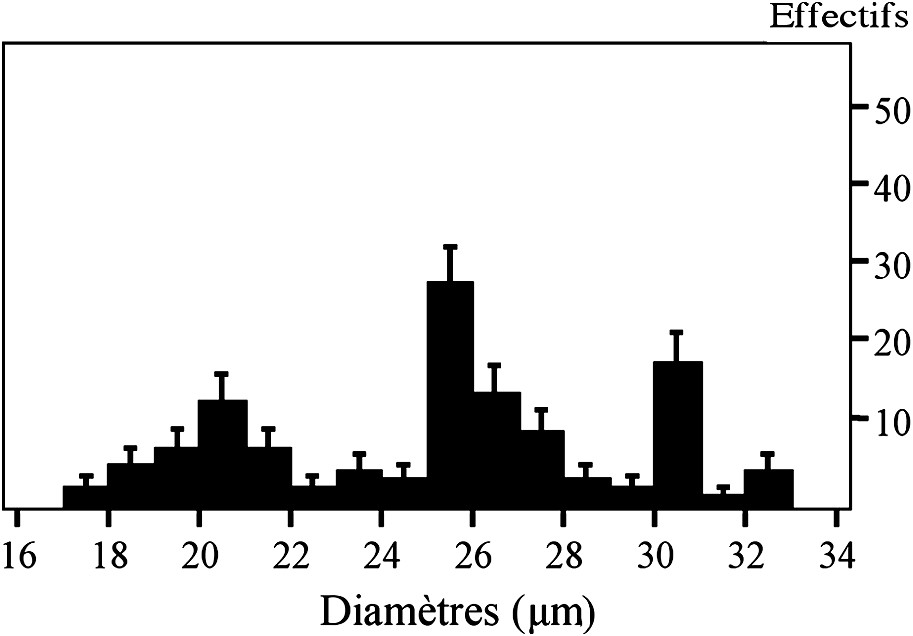
Distribution des diamètres (μm) des grains de pollen pour un individu du clone T1. Les barres d'erreur, au risque 5%, sont portées sur chaque pic.
Nos observations ne permettent pas d'établir de corrélation entre la longueur des étamines, les dimensions et la couleur du pollen, car, dans une même loge pollinique, on trouve toujours trois types de pollen, un petit de couleur orange, des grains moyens et des gros de couleur jaune ; en outre, d'autres paramètres sont impliqués : densité moyenne et taille moyenne des pores, forme des opercules, et densité moyenne des épines (Tableau 2).
3.4 Trimorphisme et hétérostylie
Les trois morphotypes sont bien distincts et portés par le même individu. Ils peuvent être séparés par de nombreux caractères (forme, taille et degrés de soudure des bractéoles), caractères utilisés dans les clés de détermination des Atriplex [4], tout comme la longueur des styles dans leur description. Mais, si l'on compare les bractéoles du morphotype (a) à celles d'A. nummularia (espèce dioïque), récoltées dans la même zone, on constate une ressemblance très remarquable portant, non seulement sur leur forme et leur degré de soudure, mais également sur la taille des styles qui sont longs, A. nummularia étant connu comme longistyle, ce que nous avons vérifié sur les échantillons de la région. De même, si l'on compare les bractéoles du morphotype (c) à celles classiquement décrites pour A. halimus, on constate encore une ressemblance, portant non seulement sur leur forme et leur degré de soudure, mais aussi sur la taille des styles, qui sont courts (Fig. 7). En outre, le pollen d'A. nummularia (

Implication d'Atriplex nummularia dans la variation intra-individuelle des fleurs d'Atriplex halimus. Styles longs (La, Lb, Lc), moyens (Ma, Mb, Mc), courts (Ca, Cb, Cc). Cette généralisation illustre les résultats présentés sur la Fig. 4.
Parmi les populations tétraploïdes étudiées en Afrique du Nord [3], la trimorphie des bractéoles et la tristylie ne se manifestent que chez deux d'entre elles, Tadjerouine et Kairouan, situées à proximité de populations d'A. nummularia. En revanche, au sein de ces dernières, on n'observe pas de trimorphisme des bractéoles, ni d'hétérostylie.
4 Conclusion
D'une manière générale, quand une espèce se révèle particulièrement polymorphe au sein d'une station ou entre stations de son aire, il convient, avant d'envisager les différentes hypothèses qui pourraient expliquer ce phénomène, de rechercher dans quelle mesure le polymorphisme interindividuel n'est pas lié à un polymorphisme intra-individuel.
4.1 Originalité de l'hétérostylie observée
L'hétérostylie mise en évidence dans les populations tunisiennes étudiées d'A. halimus est unique :
- • c'est le seul cas observé jusqu'à maintenant dans l'espèce et le genre, et plus généralement dans la famille des Amaranthacées, à laquelle appartiennent les Atriplex ;
- • cette hétérostylie concerne normalement les fleurs femelles, alors que, jusqu'à présent, elle n'était signalée que chez des fleurs hermaphrodites ;
- • si, dans les cas d'hétérostylie classique, les caractères des pièces florales stériles sont invariables, A. halimus révèle ici trois morphotypes floraux, d'après la taille, la forme et le degré de soudure des bractéoles ;
- • enfin et surtout, la tristylie des populations tunisiennes est observée au niveau intra-individuel et non interindividuel, comme d'habitude ; chaque pied porte neuf types floraux résultant de la combinaison des trois types de styles et de bractéoles (styles longs La, Lb, Lc ; moyens Ma, Mb, Mc ; courts Ca, Cb, Cc).
Il n'en reste pas moins que les différentes manifestations morphologiques de l'hétérostylie, qu'elles soient inter- ou intra-individuelles, sont les mêmes. C'est ainsi que, dans le cas de la tristylie classique, Darwin [5] signale, chez Oxalis valdiviana et Oxalis regnelli, la présence, comme chez les deux populations d'A. halimus étudiées, de grains de pollen de trois tailles différentes et de deux couleurs.
4.2 Intérêt pour l'espèce
Quel type de pollen assure la fécondation chez chacun des morphotypes ?
Généralement, chez les espèces présentant de l'hétérostylie au niveau de fleurs hermaphrodites, le gros pollen des fleurs à style court pollinise les fleurs à style long, et réciproquement pour le petit pollen. Ce mécanisme, en rapport avec l'auto-incompatibilité, n'est jamais total comme Darwin [5] avait pris soin de le signaler, en particulier chez les Primevères.
On peut remarquer que la notion d'auto-incompatibilité classique ne s'appliquerait pas sensu stricto dans le cas de notre matériel, puisque petits et gros grains de pollen proviennent justement du même individu et qu'il y a obligatoirement autocompatibilité, la compatibilité étant une notion se jugeant à l'échelle de l'individu et non de la fleur elle-même.
En d'autres termes, dans le cas d'une hétérostylie classique (interindividuelle), distylie ou tristylie, les plantes sont désavantagées si elles sont isolées, puisque la probabilité est alors faible qu'une plante longistylée soit pollinisée par une plante brachystylée, alors que, pour un individu isolé des provenances étudiées, compte tenu de son autocompatibilité globale, une descendance abondante est assurée, comme nous avons pu le vérifier. Ce dispositif est d'une importance majeure, l'individu se comportant alors comme une population à lui seul, la variation entre populations ou entre individus d'une même population se retrouvant, ici, au niveau d'une seule plante.
4.3 Hypothèses sur les causes de la variation intra-individuelle
Il est à remarquer que les variations de la longueur du style sont maintenues quelles que soient les conditions de l'environnement (serre et île de Porquerolles), ce qui nous conduit à rechercher des causes qui ne soient pas d'ordre physiologique.
Nous avons vu l'étroite corrélation qui existe entre le morphotype (a) et A. nummularia d'une part, le morphotype (c) et A. halimus d'autre part. En outre, il faut signaler l'existence de sept morphotypes intermédiaires résultant de la combinaison entre la forme des bractéoles et la longueur du style. On peut alors se demander si l'hétérostylie intra-individuelle ne serait pas liée à l'hybridation de ces deux espèces tétraploïdes : A. halimus, monoïque, à style court, et A. nummularia, dioïque, à style long, espèces chez lesquelles il n'a jamais été signalé, jusqu'à présent, de cas d'hétérostylie.
4.3.1 Hybridation et introgression
Des résultats préliminaires ont montré que, pour les deux clones, le poids des semences et des graines est le plus élevé pour le morphotype (b), quelle que soit la longueur du style, celles-ci germant aussi plus rapidement. Ce caractère évoque la vigueur hybride. Cette différence de vitesse de germination peut être intéressante en milieu aride.
L'hypothèse d'une hybridation semble confirmée par des croisements artificiels que nous avons effectués entre A. halimus et A. nummularia. Sur 100 akènes issus de ces croisements, 12 contenaient des graines bien formées qui ont germé [3]. On peut estimer cette proportion d'autant moins négligeable que, dans la nature, ces deux espèces ont, dans les régions étudiées, des périodes de floraison se recouvrant partiellement. Il n'en reste pas moins, que dans le cas d'une hybridation classique, les deux individus croisés sont de génotypes différents, alors que, dans le cas de notre matériel, un seul individu, représentant théoriquement un seul génotype, comporte des caractères (a) et (c) des deux espèces « parentales », A. halimus et A. nummularia. Tout se passe comme si un pied de nos cultures était l'équivalent d'une population de morphotypes (a), (b) et (c), chacun manifestant de la tristylie, ce qui peut faire penser à une introgression, mais, comme pour l'hybridation éventuelle, on est conduit à conclure à l'équivalence entre une plante et une population (Fig. 7).
4.3.2 Une plante, assemblage de génotypes et de phénotypes différents
Tout ce qui précède suggère l'existence d'une variation génétique intra-individuelle. Il a déjà été montré qu'une plante peut se comporter comme un assemblage de phénotypes et de génotypes différents d'une même espèce. C'est ainsi que Thomson et al. [6], en testant la variabilité de 18 systèmes enzymatiques sur diverses espèces de figuier étrangleur, ont mis en évidence des différences alléliques entre les branches d'un même arbre : au total 45 différences alléliques pour 13 arbres, représentant six espèces. Murawski [7] obtient des résultats comparables en travaillant sur la couronne d'arbres appartenant à deux espèces de Légumineuses guyanaises. Cet auteur et Hallé [8] pensent que la croissance indéfinie d'un végétal favoriserait l'accumulation de mutations somatiques à l'origine de la variabilité intra-individuelle. Mais, dans le cas de notre matériel, la plante se comporte comme un assemblage de phénotypes et de génotypes de deux espèces différentes, A. halimus et A. nummularia. En effet, parmi les neuf catégories de fleurs femelles observées sur une même plante (Fig. 7), A. halimus et A. nummularia correspondent respectivement aux phénotypes (Cc) et (La).
Dans ce cas, on ne peut invoquer des mutations somatiques pour tenter d'expliquer, sur des bases génétiques classiques, une telle diversité au sein d'un même individu. En effet, il est impossible qu'un ensemble de mutations fasse apparaître une série de caractères (bractéoles, style, pollen) propres à l'A. nummularia, à tous niveaux de la plante, et en grand nombre, chez un seul individu d'A. halimus.
A. halimus est une espèce originale, car elle révèle un mélange de particularités de l'appareil reproducteur à l'échelle de la plante [9–12], dont l'importance relativise la notion d'individu. L'existence de ce nouveau modèle expérimental qu'offrent les individus de ces populations tunisiennes hétérostyles permet aux biologistes et aux généticiens de poser des problèmes, comme ceux de la variation génétique intra-individuelle et de l'hybridation interspécifique, accessibles par les méthodes de la biologie moderne.
Remerciements
Les auteurs remercient le Pr. A. Nougarède pour l'intérêt qu'elle a porté à ce travail, le Conservatoire botanique national méditerranéen (France) pour l'entretien de la collection située à Porquerolles. Ces recherches ont reçu l'appui de l'Union européenne (contrat ERB IC 18-CT98-0390).