

1 Introduction
The term stress is most often used subjectively and with various meanings. The physiological definition and appropriate term for stress are referenced as responses to different situations. The flexibility of normal metabolism allows the development of responses to environmental changes, which fluctuate regularly and predictably over daily and seasonal cycles. Thus, every deviation of a factor form and its optimum does not necessarily result in stress. Stress with a constraint or with highly unpredictable fluctuations imposed on regular metabolic patterns causes injury, disease, or aberrant physiology. Stress is the altered physiological condition caused by factors that tend to alter equilibrium. Strain is any physical and chemical change produced by a stress [1–4].
Environmental stresses trigger a wide variety of plant responses, ranging from altered gene expression and cellular metabolism to changes in growth rate and plant productivity. Plant reactions exist to circumvent the potentially harmful effects caused by a wide range of both abiotic and biotic stresses, including light, drought, salinity, and high temperatures. Among the environmental stresses, drought stress is one of the most adverse factors to plant growth and productivity. Knowledge of the biochemical and molecular responses to drought is essential for a holistic perception of plant-resistance mechanisms to water-limited conditions in higher plants [5–11].
Water stress is the major problem in agriculture and the ability to withstand such stress is of immense economic importance. Water-stress tolerance involves subtle changes in cellular biochemistry. It appears to be the result of the accumulation of compatible solutes and of specific proteins that can be rapidly induced by osmotic stress [12,13]. The numerous physiological responses of plants to water deficits generally vary with the severity as well as with the duration of water stress [14–23]. Related responses induced by water deficits were summarized in Fig. 1.
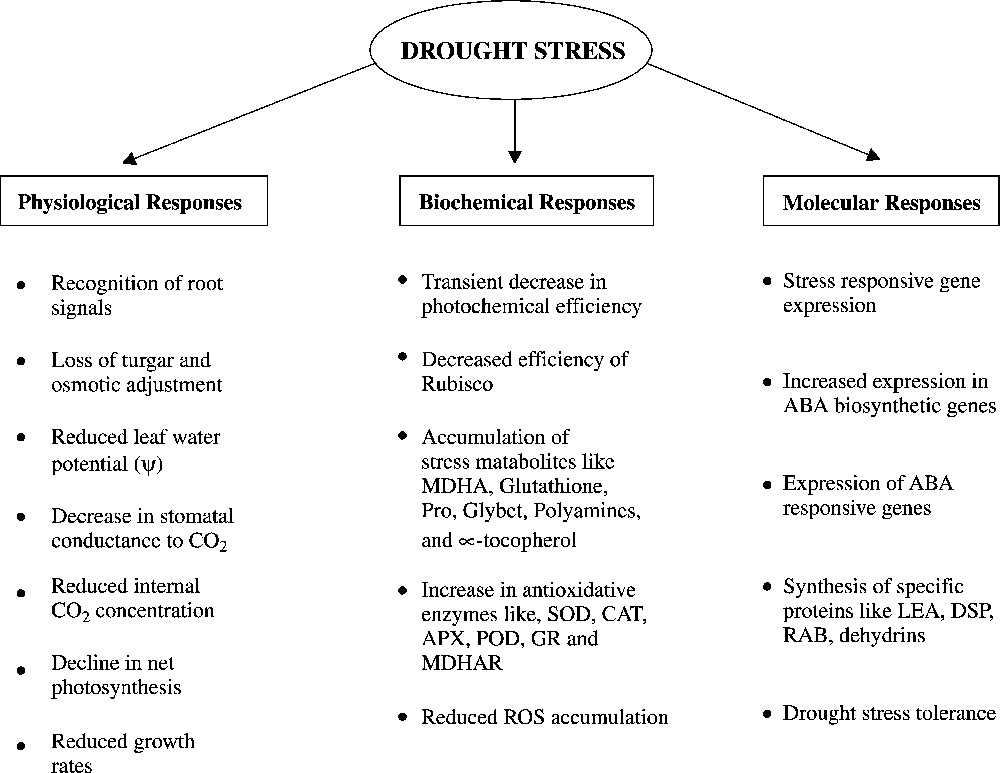
Physiological and molecular basis of drought stress tolerance (see [7,18]).
Water-deficit stress can be defined as a situation in which plant water potential and turgor are reduced enough to interface with normal functions [2–6,24–27]. Water stress is considered to be a moderate loss of water, which leads to stomatal closure and limitation of gas exchange. Desiccation is a much more extensive loss of water that can potentially lead to gross disruption of metabolism and cell structure and eventually to the cessation of enzyme catalyzing reactions (Smirnoff, 1993). Water stress is characterized by reduction of water content, turgor, total water potential, wilting, closure of stomata, and decrease in cell enlargement and growth. Severe water stress may result in arrest of photosynthesis, disturbance of metabolism, and finally death [28–32].
Water stress influences plant growth at various levels, from cell to community [33,34]. The quantity and quality of plant growth depend on cell-division enlargement, and differentiation, and all of these events are affected by water stress [35,36]. Hsiao (1973) concluded that water stress inhibits cell enlargement more than cell division. It reduces plant-growth inhibition of various physiological and biochemical processes, such as photosynthesis, respiration translocation, ion uptake, carbohydrates, nutrient metabolism, and hormones [37–39].
Drought stress has adverse influence on water relations in Arachis hypogeae [40], photosynthesis in peanuts [41] and mineral nutrition, metabolism, growth and yield of groundnut [42,43]. In addition, drought conditions influence the growth of weeds, agronomic management, and nature and intensity of insects, pests, and diseases in Arachis hypogeae [44–47].
WUE (water-use efficiency) is traditionally defined as the ratio of dry-matter accumulation to water consumption over a season. Increasing WUE could theoretically affect plant growth. When water is limited, the productivity of plants that use a finite water supply more efficiently would be positively affected, as in peanut [48]. WUE measurements may be made at three levels: (i) in single leaf using gas-exchange techniques, (ii) in whole plants grown in containers, and (iii) at the canopy level based on evapotranspiration in the field [49,50].
Variation in WUE within species can be assessed gravimetrically. However, reliable estimates of WUE under field conditions may be difficult, owing to the lack of technologies to assess the belowground biomass. However, gravimetric technique can be adequately adopted to estimate genotypic variation in pot-culture experiments. Recent studies have shown that carbon isotope discrimination occurring during carbon assimilation by leaves is closely related to WUE in various crops [52–59], suggesting that carbon isotope discrimination technology can be used to screen genotypes for WUE.
Plant growth and productivity is adversely affected by nature wrath in the form of various abiotic and biotic stress factors. Plants are frequently exposed to many stress conditions, such as low temperature, salt, drought, flooding, heat, oxidative stress, and heavy-metal toxicity. Various anthropogenic activities have accentuated the existing stress factor [60,61]. Water stress may arise from two conditions, either due to excess of water or water deficit. The more common water stress encountered is water-deficit stress, known as drought stress. Water deficit stress has a profound impact on ecological and agricultural systems [62]. The reactions of the plant to water stress differ significantly at various organizational levels, depending upon the intensity and the duration of stress as well as on the plant's species and its stage of development [61–63]. Understanding the plants' responses to drought is of great importance and is also a fundamental part for making the crops stress tolerant [7,8,63].
In sunflower, a number of responses to water deficits have been identified, which were associated with better performances under drought conditions [64]. One trait, which is potentially very important because of the interactions between temperature and water supply on plant growth, is that of enhanced early vigour [65,66]. Sunflowers' basal temperatures are higher than those of winter cereals [67]. There are reports on osmotic adjustment under drought stress in sunflower. Osmotic adjustment can contribute to yield maintenance under pre-anthesis drought conditions in the sunflower [68–70].
2 Effect of water deficit on morphological parameters
Growth is one of the most drought-sensitive physiological processes due to the reduction of turgor pressure. Cell expansion can only occur when turgor pressure is greater than the cell wall yield threshold. Water stress greatly suppresses cell expansion and cell growth due to the low turgor pressure [71,72].
Mostly plants acclimated to drought by osmotic adjustment. The loss of metabolic activity occurred only at severe stress conditions. The increased stomatal resistance under stress levels indicates the efficiency of the species to conserve water in Albizzia [73,74]. Water deficit reduced the plant growth in pearl millet under drought stress by Kusaka et al. (2005) and in okra [75].
Water is being used economically for growth processes. WUE maintains the better metabolic status of the plant, thus better growth potential in multipurpose agroforestry tree species is achieved [76,77]. Various internal and external factors influence growth besides its genetical make-up. Growth is an important tool for assessing crop productivity in various crops [78,79]. Drought at any phenophase can affect almost every aspect of growth of the above- and below-ground parts in multipurpose agroforestry tree species [77,78]. Osmotic regulation can enable the maintenance of cell turgor for survival or assist plant growth under severe drought conditions in pearl millet [80].
Crops are exposed to a variety of environmental stresses, viz., drought, salinity or low temperature constitute some of the most serious limitations to crop growth in Helianthus annuus [81,82]. The reduction in plant height is associated with the decline in the cell enlargement and more leaf senescence in the plant Abelmoschus esculentus under water stress [83].
2.1 Root length
Root characteristics, especially root length, root length density, and the number of thick roots, are important for a plant to have comparatively well-established aboveground parts by exploiting the available water, as in rice [84]. Drought avoidance due to a profound root system that enhances the ability of a plant to capture water is a fundamental adaptation mechanism to drought [85–91].
A prolific root system can confer the advantage to support accelerated plant growth during the early crop growth stage and extract water from shallow soil layers that is otherwise easily lost by evaporation [92]. More severe drought stress suggests that the dynamics of root growth under drought conditions might be a key factor to the understanding of the contribution of roots to drought avoidance [93].
Drought stress decreased the root length in Albizzia seedlings [94]. Similar results were observed in Erythrina seedlings [95], Eucalyptus microtheca seedlings [96], and Populus species [97]. Water stress reduces the biomass of fibrous roots in Avocado cultivars [98] and in pearl millet [84,86].
The root-to-shoot ratio increases under water-stress conditions to facilitate water absorption [98]. The growth rate of wheat and maize roots was found decreasing under moderate and high water-deficit stress [99]. However, the development of the root system increases water uptake and maintains the right osmotic pressure through higher proline levels [100–104]. An increased growth was reported by [72] in mango under water stress. The root dry weight decreased under mild and severe water stress in sugar beet [105]. A significant decrease in root length was reported in water-stressed Populus species by [106]. The importance of root systems in acquiring water has long been recognized. A prolific root system can confer the advantage to support accelerated plant growth during the early crop-growth stage and extract water from shallow soil layers that is otherwise easily lost by evaporation [107]. Past studies report that the root-to-shoot ratio increases under water-stress conditions to facilitate water absorption [108] and that it is related to the ABA content of roots and shoots [109]. The root growth was not significantly reduced under water deficits in maize and wheat [110].
2.2 Stem length
Stem length was decreased in Albizzia seedlings under drought stress [111]. Similar results were observed in Erythrina [112], Eucalyptus microtheca seedlings [96], and Populus species [98]. Continuous water deficit results in fewer and smaller leaves, which have smaller and more compact cells and greater specific leaf weight in peanut [113].
Water stress was a very important limiting factor at the initial phase of plant growth and establishment. There was a significant reduction in shoot height in Populus cathayana under deficit stress [97]. In soybean, the stem length decreased under water-deficit stress, but this decrease was not significant when compared to well-watered control plants. The plant height reduced up to 25% in water-stressed Citrus seedlings [114]. Stem length was significantly affected under water stress in potato [115].
2.3 Leaf area
During water stress, the total leaf area per plant decreased significantly in Eragrostis curvula, Oryza sativa, Abelmoschus esculentum, and in Asteriscus maritimus [116–120]. Leaf-area plasticity is important to maintain the control of water use in crops. In Sorghum, leaf area reduced significantly under water stress. This reduction occurred before stomatal conductance decreased in the remaining viable leaf area [26,35,89].
Leaf area was affected adversely in both main shoot and tillers of all the varieties. Reduction in leaf area by water stress is an important cause of reduced crop yield through reduction in photosynthesis [116]. The reduction in plant height and leaf area under water stress may be associated with the decline in the cell enlargement and more leaf senescence in Abelmoschus esculentum.
Leaf water potential, osmotic potential, and relative water content decreased in stressed plants at all the growth stages in sorghum. The decrease in osmotic potential in response to water deficit was more compared to the leaf water potential at all the growth stages, indicating the ability of the leaves to maintain turgor through osmotic adjustment in sorghum [117–129].
Water deficits reduce the number of leaves per plant and individual leaf size, leaf longevity and leaf reduced by decreasing the soil's water potential. Leaf area expansion depends on leaf turgor, temperature, and assimilating supply for growth, which are all affected by drought in Arachis hypogeae [7–11].
Water-deficit stress mostly reduced leaf growth and in turn the leaf area in many species of plants, like Populus, Ziziphus, etc. [128–131]. Significant interspecific differences between two sympatric Populus species were found in the total number of leaves, total leaf area, and total leaf biomass under drought stress. The leaf growth was more sensitive to water stress in wheat, but it was not so in the case of maize [132].
2.4 Fresh and dry weight
Prolonged water stress reduced the biomass of fibrous roots in Avocado cultivars [133]. The performance and biomass production potential of trees depend on the maintenance of a higher physiological status and economical utilization of resources in agroforestry tree species. A decrease in total dry matter may be due to the considerable decrease in plant growth, photosynthesis and canopy structure, as indicated by leaf senescence during water stress in Abelmoschus esculentum. Changing resource pools (e.g., water or nutrient availability) may also affect the distribution of biomass [134–137]. Drought stress decreases mean plant biomass, whereas it increases both the relative variation in plant biomass and the concentration of mass within a small fraction of the population. This is supported by earlier studies [138,139] conducted at single field sites or in pots.
Drought stress decreased the plant biomass in Cyamopsis tetragonoloba [140] and spring wheat. Similar results were observed in earlier studies in wheat, Asteriscus maritimus, and Albizzia seedling [140–142]. Leaf biomass was less in water-stressed plants compared to unstressed plants in the case of tree species. The reduction in total biomass was reported in groundnut cultivars under water stress due to the reduction in the pod mass rather than in the vegetative mass [143]. Morphological parameters like fresh and dry weights have a profound effect in water-limited conditions. There was a one-third reduction in fresh and dry weights of the Ziziphus rotundifolia plant under drought conditions [144]. Progressive drought resulted in a significant reduction in early allocation of dry matter and decreased fresh and dry weight in all plant parts in Populus davidiana. Under water-deficit stress, the biomass production was decreased in Populus cathayana and drought severely affected all growth parameters. Plant productivity under drought stress is strongly related to the processes of dry matter partitioning and temporal biomass distribution [145]. A moderate stress tolerance, as shown by dry weight production in transgenic plants, was noticed based on relative shoot growth studies under stress conditions, like drought [146]. Defoliation originating from water stress in maize plants resulted in reduced biomass and thin biomass [147]. Regulated deficit irrigation and partial root drying caused a significant reduction in shoot biomass when compared to control in common bean plants [148]. There was a significant reduction in root dry weight on induction of drought stress in cotton by PEG [149]; there was a significant reduction in shoot dry weight due to water-stress treatments in sugar beet genotypes [150], and mild stress affected the dry weights of shoots, while shoot dry-weight loss was greater than root dry-weight loss under severe stress. Reduced biomass was met with water-stressed soybean plants. The dry weight of Poncirus trifoliate seedlings decreased to a considerable extent under water stress.
A common adverse effect of water stress on crop plants is the reduction in fresh and dry biomass production [151]. Reduced biomass production due to water stress has been observed in almost all genotypes of sunflower [72,87,92]. However, some genotypes showed better stress tolerance than others did. Tahir et al. (2002) evaluated 25 inbred lines of sunflower for drought tolerance. They reported a decrease in plant height, leaf area, head diameter, 100-achene weight, yield per plant, and plant biomass due to water stress. They further suggested that these traits could be used as a selection criterion for higher yield per plant under water deficit. In a field experiment, Prabhudeva et al. (1998) subjected sunflower genotypes to water stress at bud initiation and/or seed filling stage. They observed that seed and biological yield were reduced most by the imposition of water stress at the bud-initiation and seed-filling stages, than at the seed-filling stage only.
2.5 Yield parameters
Sunflower yields were higher for winter season than for spring plantings [151,152]. In early plantings of sunflower, the yield increase was associated with both an increase in grain number and in individual grain weight. There have been reports on increased yield under water-limited conditions in various plants [56,62,79,83]. Sinclair and Muchow (2001) analyzed many physiological and morphological traits that could contribute to an increase in grain yield in drought situations. The portioning of dry matter to the head is critical in the process of yield determination in water-stressed sunflower [149]. The effect of water deficits on the harvest index of sunflower is complex due to the interactions between the timing and intensity of the stress relative to the developmental processes that determine the components of yield.
Experiments with sunflower where water deficits were imposed at different growth stages generated a two-fold difference in the harvest index [124,130], challenging the view that harvest index may be considered constant over a range of water deficits. Drought during the reproductive stage reduces the harvest index.
The yield components, like grain yield, grain number, grain size, and floret number, were found decreased under pre-anthesis drought stress treatment in sunflower. Water stress greatly reduced the grain yield of maize plants, and this reduction in grain yield was dependent upon the level of defoliation water stress during early reproductive growth reduces yield in soybean, usually as a result of fewer pods and seeds per unit area [150,151]. Under non-compact soil conditions, salinity, water logging, and saline-watered logged treatments significantly reduced grain yield in wheat genotypes. In water-stressed soybean, the seed yield was far below when compared to well-watered control plants [152–154].
Seed yield and yield components are severely affected by water deficit. Water stress reduced the head diameter, 100-achene weight and yield per plant in sunflower [18,26,33,41]. These scientists also observed significant but negative correlation of head diameter with fresh root and shoot weight under water stress. A positive and significant relation was recorded between dry shoot weight and achene yield per plant. Reddy et al. (1998) supported the view that water stress for more than 12 days at the grain-filling and flowering stage of sunflower (grown in sandy loam soil) was most damaging and reduced the achene yield. Mozaffari and Zeinali (1997) suggested that higher stalk diameter, greater plant height, and seed yield are suitable as selection criteria for drought-tolerant cultivars. Nandhagobal et al. (1996) subjected sunflower to water stress by skipping irrigation at germination, vegetative stage, button initiation, flowering, and at seed-filling stages. Skipping irrigation at flowering stage caused much reduction in yield during the summer season. Water stress at flowering also gave the lowest seed oil content.
3 Conclusions and perspectives
Drought is a worldwide problem, constraining global crop production and quality seriously, and recent global climate change has made this situation more serious. Drought is also a complex physical-chemical process, in which many biological macromolecules and small molecules are involved, such as nucleic acids, proteins, carbohydrates, lipids, hormones, ions, free radicals, mineral elements. In addition, drought is related to salt stress, cold stress, high-temperature stress, acid stress, alkaline stress, pathological reactions, senescence, growth, development, cell circle, UV-B damage, wounding, embryogenesis, flowering, signal transduction, and so on. Therefore, drought is connected with almost all aspects of biology [8–12,16–23,39,50,51]. Currently, drought study has been one of the main directions in global plant biology and biological breeding. Anatomical changes induced by water deficits in higher plants were better-observed indicators, which can be directly applied to agriculture and handled. To aim at exploring efficient anatomical indices, much information has been documented, but more attention should be paid to link them with physiological and molecular one.
In a word, the study of the physiological mechanisms of wheat anti-drought has much work to do. Molecular biology aspects of wheat cannot substitute for this important part, but strengthen the research and provide a broad future and platform. It is easy to see that one cell or molecule cannot be alive in natural fields and not provide any economic effect for human beings. The combination of molecular biology, plant physiology, and of other related disciplines is the key. Many achievements in biotechnological and traditional breeding of wheat are good examples. Although some progresses in terms of exploration of the molecular nature of wheat anti-drought have also taken place, many problems exist. Currently, from the view of the globe, sustainable development is the key point. The necessary way to solve the issue of sustainable development is by biological measures, in which plants will play greater roles and crops will play the greatest functions with no doubt. To aim at taking advantage of the full use of crop physiological potential for high production and safe food with better quality, the following problems remain to be solved. What is the relationship between mineral elements (in particular, K+ and Na+ in soils) with root signal transduction (pathways)? Many former studies showed that K+ was little connected with anti-drought behaviour, but recent research and our results displayed that it was linked with wheat's drought-resistance. What is the exact soil water-stress threshold of the individual wheat genotype? This is of much importance to drought-resistant plant breeding, to saving-water agriculture, and to precise agriculture under global climate change. What are the details that constitute the network of the regulatory system of drought, cold, UV-B, freezing, acidity, salinity, wounding, pathogenicity, senescence, cell death? How is each one linked with other parts? What is the (transient) connection among different physiological adaptive regulatory pathways at different levels? What role do endogenous hormones play in this course? What is the crosstalk among them when abiotic or/and biotic stress happen(s)? The redox state in plants is important, and how is it regulated by drought signal? What is the best allocation of different crops and grass-shrub-forest in worsening arid and semi-arid areas for obtaining economic and ecological effects? A widespread use of data resources for fine gene functions and structure of different plants (species) being from model plants, Arabidopsis thaliana and rice, how large is their reliability? No doubt, expanded detecting of the plants' range is more urgent. Therefore, we think that there is much work to do in physiological studies at different scales with the increasing atmospheric change.
Acknowledgements
Research in Professor Shao HB's laboratory is jointly supported by the 973 Project of China (2007CB106803), the Doctoral Foundation of QUST (0022221), the Shao Ming-An's Innovation Team Projects of Education Pinistry of China, and the Specialized Initiation Foundation of Excellent Ph.D. Dissertation of the Chinese Academy of Sciences.