

1 Introduction
Plants are subjected to several environmental stresses that adversely affect growth, metabolism and yield. Water is one of the most important environmental factors that regulate plant growth and development. Drought is the major limiting factor in many parts of the world, which seriously affects plant growth and yield [1]. The lack of adequate moisture leading to water stress is common occurrence in rain-fed areas, brought about by infrequent rains and poor irrigation [2]. Water deficit (commonly known as drought) can be defined as the absence of adequate moisture necessary for a plant to grow normally and complete its life cycle [3]. Plants experience drought stress either when the water supply to roots becomes difficult or when the transpiration rate becomes very high [4].
Among the diverse consequences of the drought effect on plant development, restricted nutrient and water acquisition are commonly recognized [5]. Increasing evidences suggest that drought induces oxidative stress in various plants, in which reactive oxygen species (ROS), such as superoxide radical (O⋅−2), hydroxy radical (⋅OH), hydrogen peroxide (H2O2) and alkoxy radical (RO⋅) are produced [6]. The toxic superoxide radical has a half-life of less than 1 s and is usually rapidly dismutated by superoxide dismutase (SOD) to H2O2, a product that is relatively stable and can be detoxified by catalase and peroxidases. These metalloenzymes constitute an important primary defence of cells against superoxide free radicals generated under stress conditions; thereby, increased SOD activity is known to confer oxidative stress tolerance [7].
Triazole compounds such as TDM, hexaconazole, uniconazole and paclobutrazol, etc., are widely used as fungicides and they also possess varying degrees of plant-growth regulating properties, mediated by their interference with the isoprenoid pathway and subsequent shift in the balance of important plant hormones, including GA, ABA and cytokinins [8]. They inhibit gibberellin and ergosterol biosynthesis in plants and fungi, respectively [9]. Triazoles affect the isoprenoid pathway and alter the levels of certain plant hormones by inhibiting gibberellin synthesis, reducing ethylene evolution and increasing cytokinin levels [10]. Triazole-treated plants have a more efficient free-radical scavenging system, which enables them to detoxify active oxygen [11]. Some of the previous works carried out in our lab revealed the morphological and physiological changes associated with triazole treatment in various plants, including inhibition of plant growth, decreased internodal elongation, increased chlorophyll levels, enlarged chloroplasts, thicker leaf tissue, increased root-to-shoot ratio, increased antioxidant potentials, and enhancement of alkaloid production [12–16].
Comparatively, little work has been devoted to water stress problems and methods to overcome drought stress injuries in this plant. The drought stress amelioration by triazole compounds is of major research interest, because these compounds have innate potentiality for increasing antioxidant enzymes and molecules in oxidative-stressed plants [1,11]. So, in the present investigation, an attempt has been made to evaluate the drought stress ameliorating ability of TDM [1-(4-chlorophenoxy)-3,3-dimethyl-1-(1H-1,2,4-triazol-1-yl)-2-butanone] [C14H16ClN3O2], , a triazole fungicide, with special emphasis to osmoregulators, like AA, PRO and GB, and to antioxidant enzymes, like APX, PPO, CAT and SOD, in drought-stressed Helianthus annuus L. plants in pot culture.
2 Materials and methods
2.1 Plant material and drought-stress induction
The triazole compound named triadimefon (TDM) was obtained as 25% WP Bayleton-registered trademark, from Bayer, India Ltd., Mumbai. Helianthus annuus L. seeds are surface sterilized with a 0.2% HgCl2 solution for 5 min, with frequent shaking, and thoroughly washed many times with deionized water to remove HgCl2. Plastic pots of 40-cm diameter and 45-cm height size were used for the study. The pots were filled with 10 kg of soil mixture containing red soil; sand and farm yard manure at 1:1:1 ratio and 240 pots were arranged in completely randomized block design (CRBD). One set of pots was kept as control and three other sets of pots were used for drought-stress and TDM treatments. Seeds were sown and the seedlings were thinned to 1 per pot on the 10th day after sowing (DAS). The plants were allowed to grow up to 30 DAS. From the 30th to the 50th day, all the potted plants were grown under a polyhouse. Control irrigation was made on alternative days, mild-stress irrigation once in three day from the 30th to the 50th day, medium-stress irrigation once in six days from the 30th to the 50th day, severe-stress irrigation once in nine days from the 30th to the 50th day, with 3-, 6-, and 9-day-interval drought (DID) with 15 mg l−1 TDM treatment and 15 mg l−1 TDM alone on the 30th DAS. After the drought period, all the pots were irrigated at 1-day intervals until harvest. Plants were uprooted randomly on the 51st DAS, washed carefully and separated into root, stem and leaves for estimating antioxidant contents and antioxidant enzyme activities.
2.2 Osmolyte concentration
2.2.1 Amino acid (AA) content
Extraction and estimation of the AA content was followed by the method of Moore and Stein [17]. 0.5 g of plant material was taken in a pestle and mortar and homogenized with 10 ml of 80% boiling ethanol. The extract was centrifuged at 800 g for 15 min and the supernatant was made up to 10 ml with 80% ethanol and used for the estimation of free AAs. 1 ml of ethanol extract was taken in a 25-ml test tube and neutralized with 0.1 N sodium hydroxide using methyl red indicator, to which 1 ml ninhydrin reagent was added. The contents were boiled in a boiling water bath for 20 min, then 5 ml of diluted reagent was added, cooled and diluted to 25 ml with distilled water. The absorbance was read at 570 nm in a spectrophotometer. The standard graph was prepared by using glycine. The AA content was calculated using the standard graph. The results were expressed in milligrams per gram of dry weight.
2.2.2 Proline (PRO) concentration
The PRO content was estimated by the method of Bates et al. [18]. The plant material was homogenized in 3% aqueous sulfosalicylic acid and the homogenate was centrifuged at 10,000 rpm. Supernatant was used for the estimation of the PRO content. The reaction mixture consisted of 2 ml of acid ninhydrin and 2 ml of glacial acetic acid, which was boiled at 100 °C for 1 h. After termination of the reaction in an ice bath, the reaction mixture was extracted with 4 ml of toluene; absorbance was read at 520 nm.
2.2.3 Glycine betaine (GB) concentration
The amount of GB was estimated according to the method of Grieve and Grattan [19]. The plant tissue was finely ground, mechanically shaken with 20 ml of deionised water for 24 h at 25 °C. The samples were then filtered and filtrates were diluted to 1:1 with 2 N H2SO4. Aliquots were kept in centrifuge tubes and cooled in ice water for 1 h. Cold KI–I2 reagent was added, and the reactants were gently stirred with a vortex mixture. The tubes were stored at 4 °C for 16 h and then centrifuged at 10,000 rpm for 15 min at 0 °C. The supernatant was carefully aspirated with a fine glass tube. The periodide crystals were dissolved in 9 ml of 1,2-dichloroethane. After 2 h, the absorbance was measured at 365 nm using GB as a standard and expressed in mg g−1 DW.
2.3 Enzyme extractions and assays
2.3.1 Ascorbate peroxidase (APX) (EC 1.11.1.1)
Ascorbate peroxidase (APX) (EC 1.11.1.1) activity was determined according to Asada and Takahashi [20]. The reaction mixture (1 ml) contained 50 mM of potassium phosphate buffer (pH 7.0), 0.5 mM of ascorbic acid, 0.1 mM of H2O2, and 200 μl of enzyme extract. The absorbance was read as the decrease at 290 nm against the blank, correction was done for the low, non-enzymatic oxidation of ascorbic acid by H2O2 (extinction coefficient: 2.9 mM−1 cm−1). The enzyme activity was expressed in U mg−1 protein (U = change in 0.1 absorbance min−1 mg−1 of protein).
2.3.2 Polyphenol oxidase (PPO) (EC 1.10.3.1)
Polyphenol oxidase (PPO) activity was assayed by the method of Kumar and Khan [21]. The assay mixture for PPO contained 2 ml of 0.1 M phosphate buffer (pH 6.0), 1 ml of 0.1 M catechol and 0.5 ml of enzyme extract. This was incubated for 5 min at 25 °C, after which the reaction was stopped by adding 1 ml of 2.5 N H2SO4. The absorbance of the purpurogallin formed was read at 495 nm. 2.5 N H2SO4 was added to the blank at the zero time of the same assay mixture. PPO activity is expressed in U mg−1 protein (U = change in 0.1 absorbance min−1 mg−1 of protein).
2.3.3 Catalase (CAT) (EC 1.11.1.6)
Catalase (CAT) was measured according to Chandlee and Scandalios [22], with modification. The assay mixture contained 2.6 ml of 50 mM potassium phosphate buffer (pH 7.0), 0.4 ml of 15 mM H2O2 and 0.04 ml of enzyme extract. The decomposition of H2O2 was followed by the decline in absorbance at 240 nm. The enzyme activity was expressed in U mg−1 protein (U = 1 mM of H2O2 reduction min−1 mg−1 of protein).
2.3.4 Superoxide dismutase (SOD) assay
The crude enzyme extract was prepared for assay of SOD by the method of Hwang et al. [23]. The enzyme protein was determined according to Bradford [24] for all the three enzymes for expressing the specific activity of enzymes. SOD (EC 1.15.1.1) activity was assayed according to Beauchamp and Fridovich [25]. The reaction mixture contained of riboflavin, 0.1 M of methionine, of potassium cyanide (KCN) and of nitroblue tetrazolium salt (NBT) dissolved in 3 ml of 0.05 M sodium phosphate buffer (pH 7.8). Three millilitres of the reaction medium were added to 1 ml of 5-enzyme extract. The mixtures were illuminated in glass test tubes by two sets of Phillips 40-W fluorescent tubes in a single row. Illumination was started to initiate the reaction at 30 °C for 1 h. Identical solutions that were kept under dark served as blanks. The absorbance was read at 560 nm in the spectrophotometer against the blank. SOD activity was expressed in units (U mg−1 protein). One U is defined as the amount of change in the absorbance per 0.1 h−1 mg−1 of protein.
2.4 Statistical analysis
The pot culture was carried out in completely randomized design (CRD). The data are expressed as mean ± SD for seven samples in each group.
3 Results and discussion
3.1 Effect of drought, TDM and their combination on osmolyte concentration
Drought stress increased the AA (Fig. 1), PRO (Fig. 2) and GB (Fig. 3) contents in all the parts of the sunflower when compared to control. Drought stress with TDM treatment leads to an enhancement in biochemical contents when compared to drought-stressed and control plants. AA content has been shown to increase under drought condition in Vigna [1]. Similar results were obtained in Abelmoschus [26] and Vigna unguiculata [3]. The accumulation of AA may be due to protein hydrolysis and also may occur in response to the change in the osmotic adjustment of their cellular contents [26]. Free AA accumulation is a more important account for most of the changes in osmotic potential. The accumulation of AA under stress at all the growth stages indicates the possibility of their involvement in osmotic adjustment [27]. Osmotic adjustment is one of the important mechanisms that alleviate some of the detrimental effects of water stress [26]. The extent of increase was higher in leaf, followed by root and stem. The GB content increased under salt stress in Catharanthus [28]. Aliphatic quaternary ammonium compounds (QAC) such as GB, stachydrine, homostachydrine, trigonelline have been found to accumulate in a large number of plants exposed to salt and water stress. Increased proline accumulation was reported in Catharanthus under different abiotic stress situations [29–31]. Increased proline in stressed plants may be an adaptation to overcome the stress conditions. Proline accumulated under stressed conditions supplies energy for growth and survival and thereby helps the plant to tolerate the stress. Proline accumulation in plants might be a scavenger and acting as an osmolyte. The reduced proline oxidase may be the reason for increasing proline accumulation [32–34].
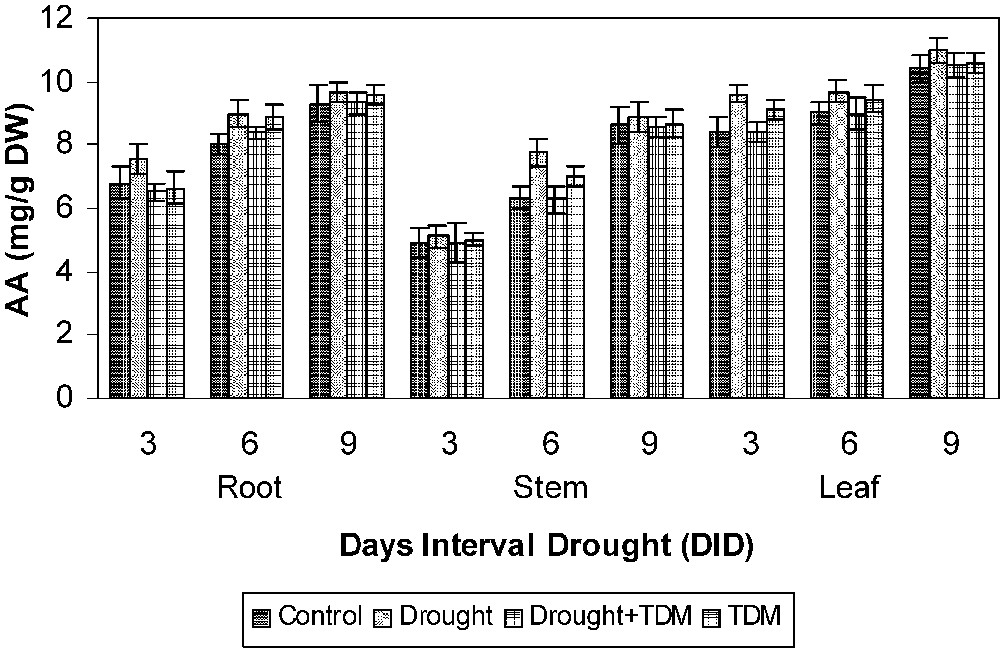
Effects of drought, TDM and their combination on the amino acid (AA) content of H. annuus plants. Values are mean ± SE of seven replicates.
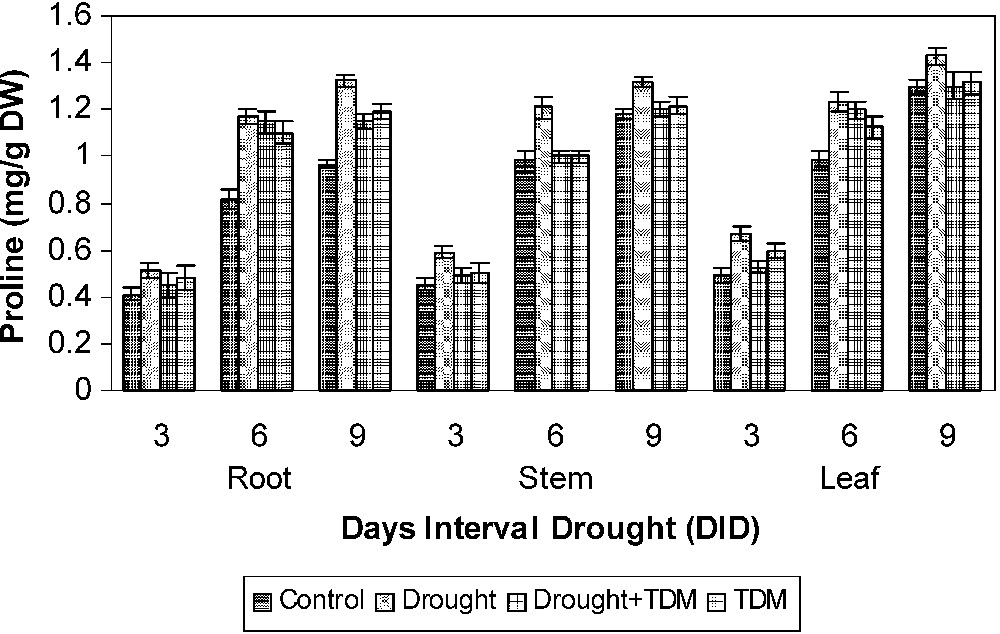
Effects of drought, TDM and their combination on the proline content of H. annuus plants. Values are mean ± SE of seven replicates.
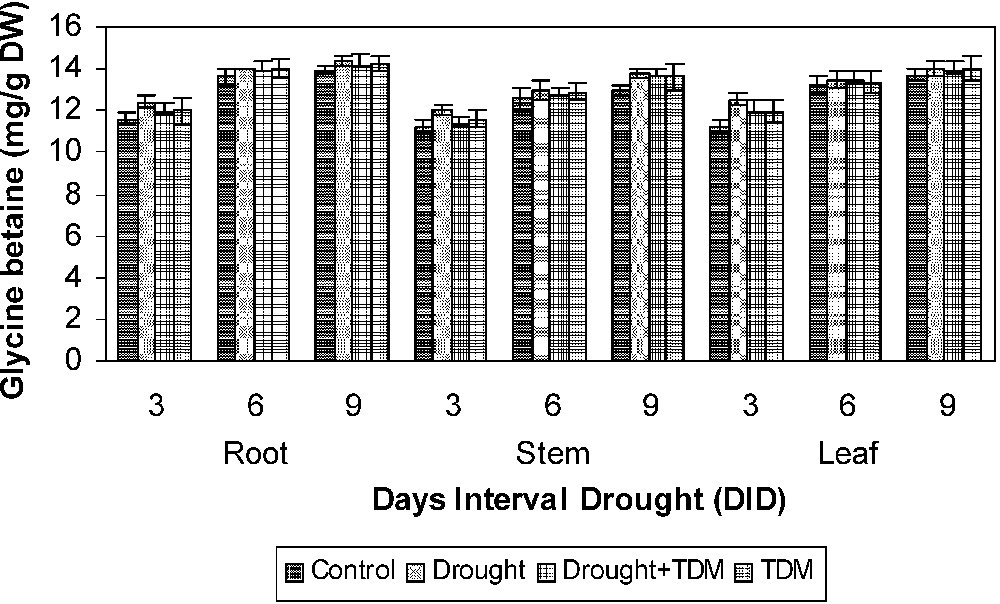
Effects of drought, TDM and their combination on glycine betaine content of H. annuus plants. Values are mean ± SE of seven replicates.
TDM increased the AA content in unstressed plants. Similar results were observed in Catharanthus [14,30,35]. Propiconazole and paclobutrazol induced a moderate increase in AA in higher plants [36–38]. Treatment with TDM significantly increased the free PRO content in Catharanthus roseus [11]. Triazole induced a transient raise in ABA content [8] and this raise in ABA induced by TDM might have caused the increase in AA and proline content in sunflower.
3.2 Effect of drought, TDM and their combination on APX activity
APX activity (Fig. 4) increases in all the drought treatments when compared to control. Drought stress with TDM treatment decreased APX activity in drought-stressed plants, and increased it in control plants. TDM treatment increased APX activity when compared to the case of control and drought-stressed plants. Similar results were obtained by many workers in many higher plants under drought stress [1,2,6,11,39–41]. APX found in organelles is believed to scavenge H2O2 produced from the organelles, whereas the function of cytosolic APX is probably to eliminate H2O2 that is produced in the cytosol or apoplast and that has diffused from organelles. In the chloroplast, H2O2 can be detoxified by the ASA–GSH–NAPDH system, which has been catalyzed by APX [10,12–14].
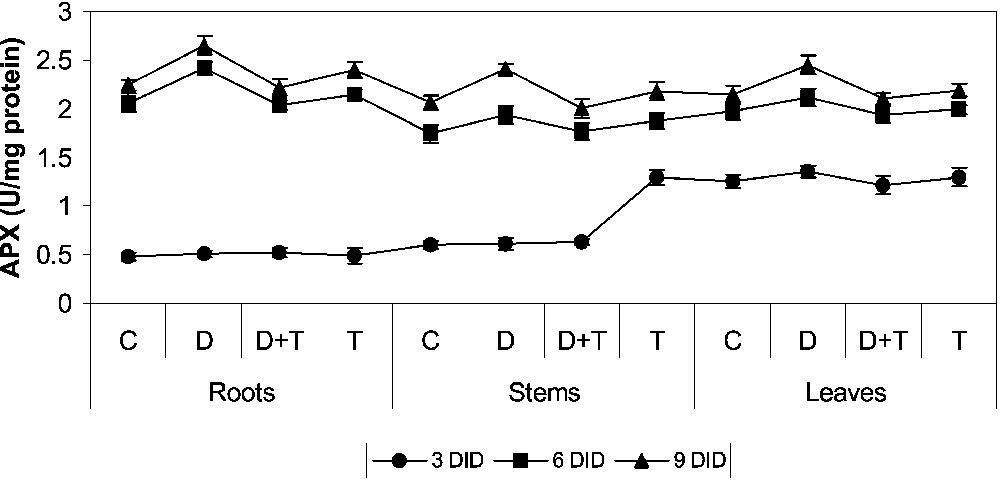
Effects of drought, TDM and their combination on ascorbate peroxidase (APX) activity of H. annuus plants. Values are mean ± SE of seven replicates.
3.3 Effect of drought, TDM and their combination on PPO activity
PPO activity increased in drought treatment when compared to control (Fig. 5). TDM treatment resulted in an enhancement of the PPO activity when compared to drought-stressed and control plants. TDM treatment in combination with drought increased PPO activity when compared to control plants. ROS scavenging is important in imparting tolerance against oxidative stress. It may be presumed that enhancement of the antioxidative system favours water-stress resistance [42,43].
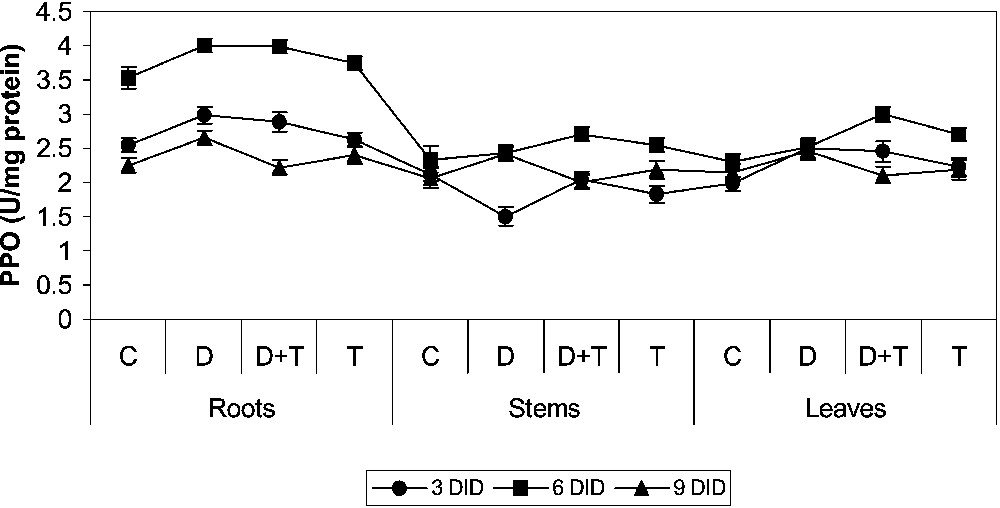
Effects of drought, TDM and their combination on polyphenol oxidase (PPO) activity of H. annuus plants. Values are mean ± SE of seven replicates.
3.4 Effect of drought, TDM and their combination on CAT activity
CAT activity (Fig. 6) was increased in all parts of drought-stressed cowpea plants and of those undergoing TDM treatment when compared to control. TDM treatment decreased catalase activity when compared to drought stress and TDM treatment, and increased it in controls. This result is in accordance with the findings in Catharanthus roseus [10,43]. The combined action of CAT and SOD converts the toxic O⋅−2, H2O2 into water and molecular oxygen, averting the cellular damage under unfavourable conditions like water stress [1].
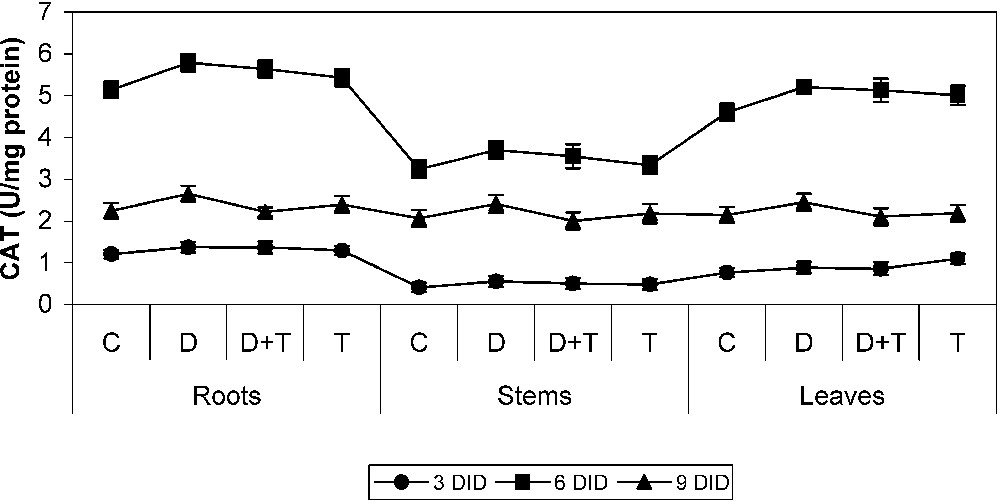
Effects of drought, TDM and their combination on catalase (CAT) activity of H. annuus plants. Values are mean ± SE of seven replicates.
3.5 Effect of drought, TDM and their combination on SOD activity
SOD activity increased in all the DID stress and with TDM treatments when compared to control (Fig. 7). TDM treatment decreased SOD activity when compared to drought-stress and TDM treatment, and increased it in the control. Triazoles increased the antioxidant potential in oxidative stressed plants under treatment when compared to control [12–14,44,45]. It was reported that SOD enhances water-stress tolerance in plants. The cytosolic Cu/Zn–SOD was induced strongly by stress, while Cu/Zn–SOD remained largely unaffected [46,47].
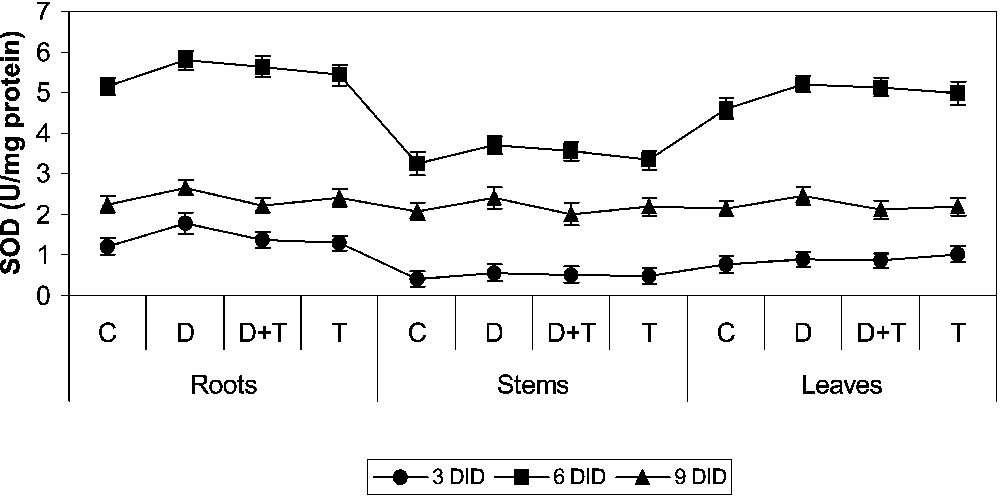
Effects of drought, TDM and their combination on superoxide dismutase (SOD) activity of H. annuus plants. Values are mean ± SE of seven replicates.
4 Conclusion
Thus, from these results, it is clear that plants are highly regulated by TDM, in terms of enhanced components of osmoprotectants under drought stress. Drought-stressed plants under TDM treatment maintain a balance between formation and detoxification of activated oxygen species, leading to partial improvement of their response to drought-induced oxidative stress.